
Role of Chitin and Chitosan in Ruminant Diets and Their Impact on Digestibility, Microbiota and Performance of Ruminants



Abstract
1. Introduction
2. Chemical Structures of Chitin and Chitosan
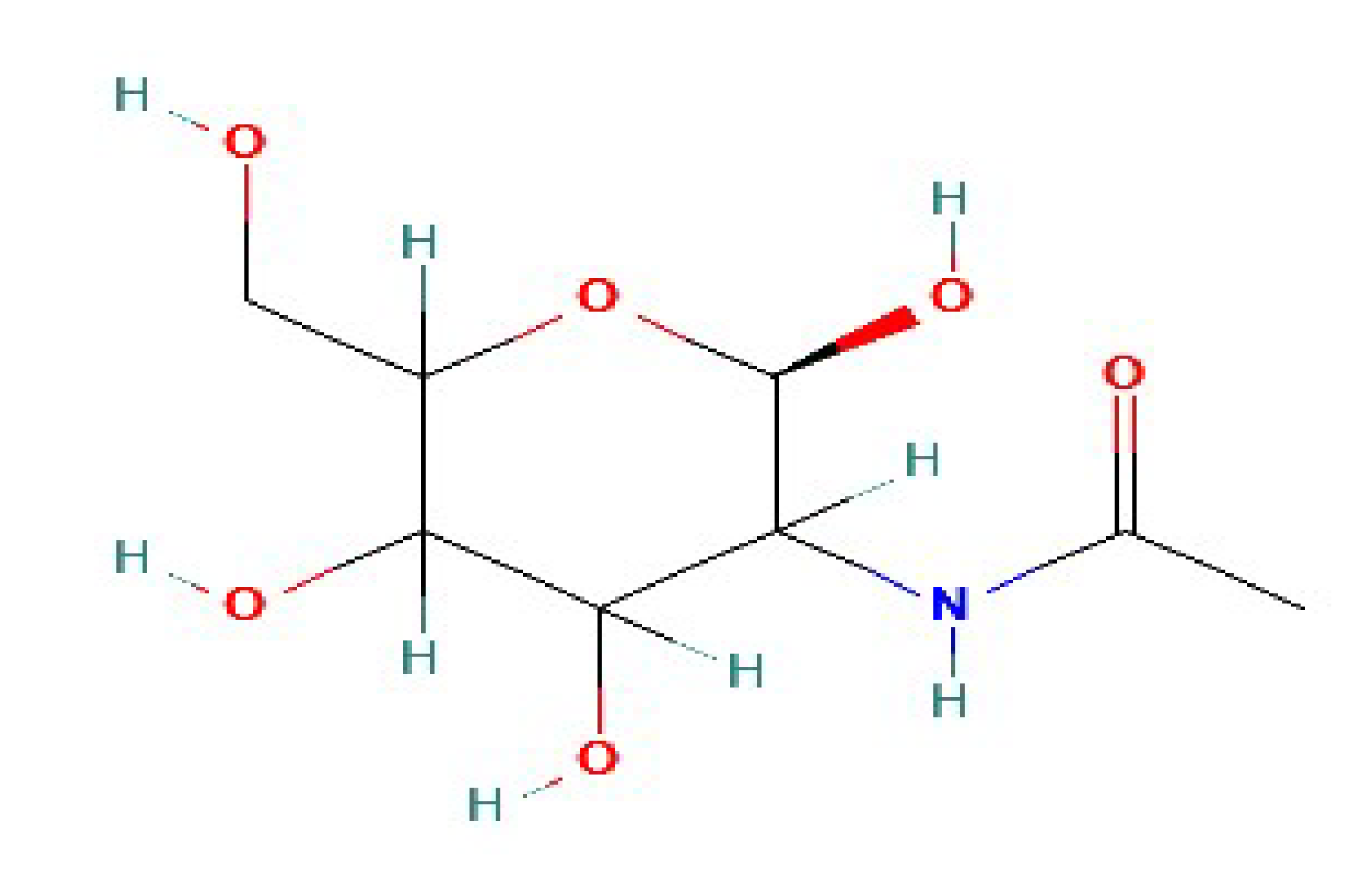
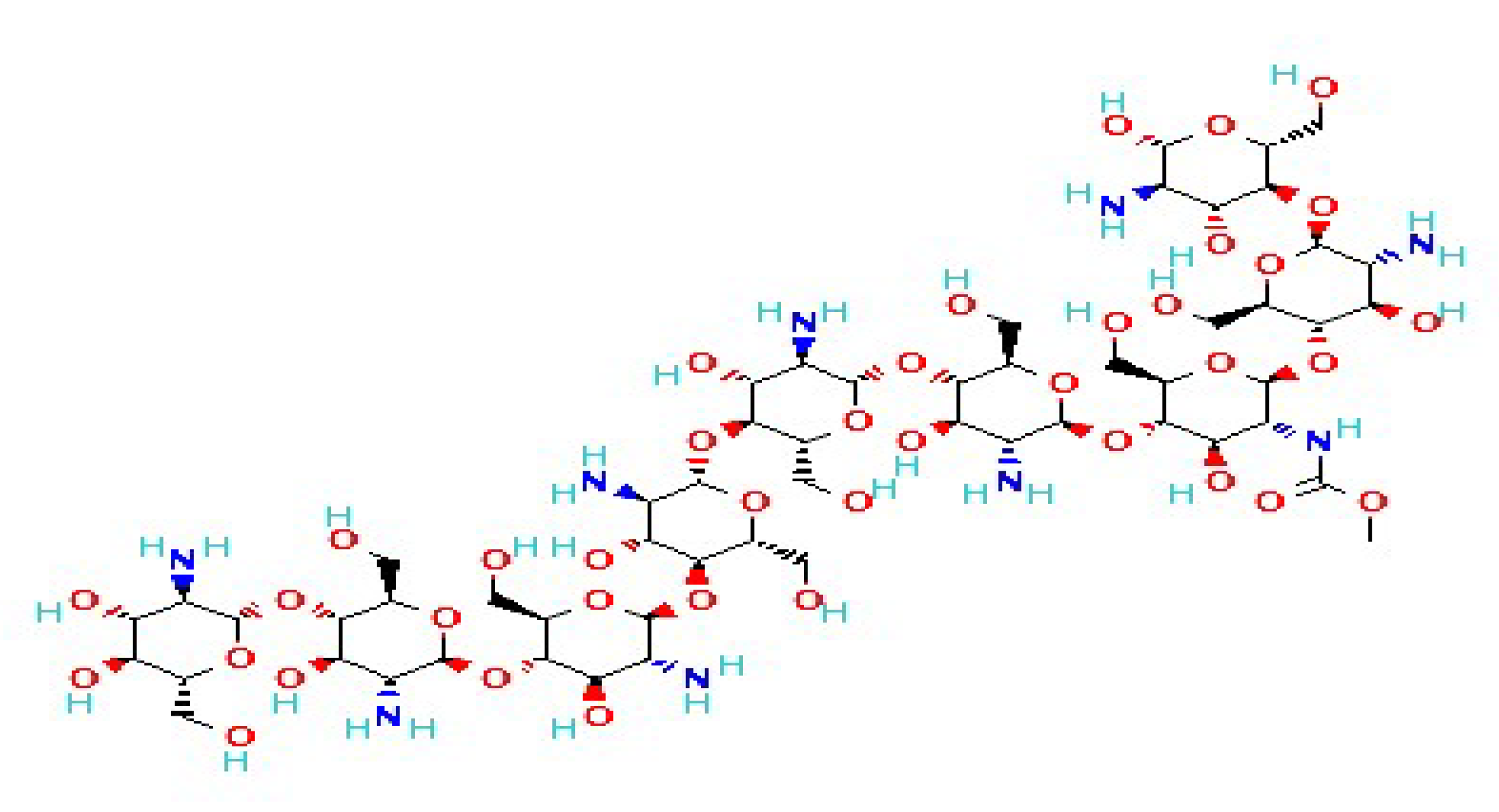
2.1. Chitin
2.2. Chitosan
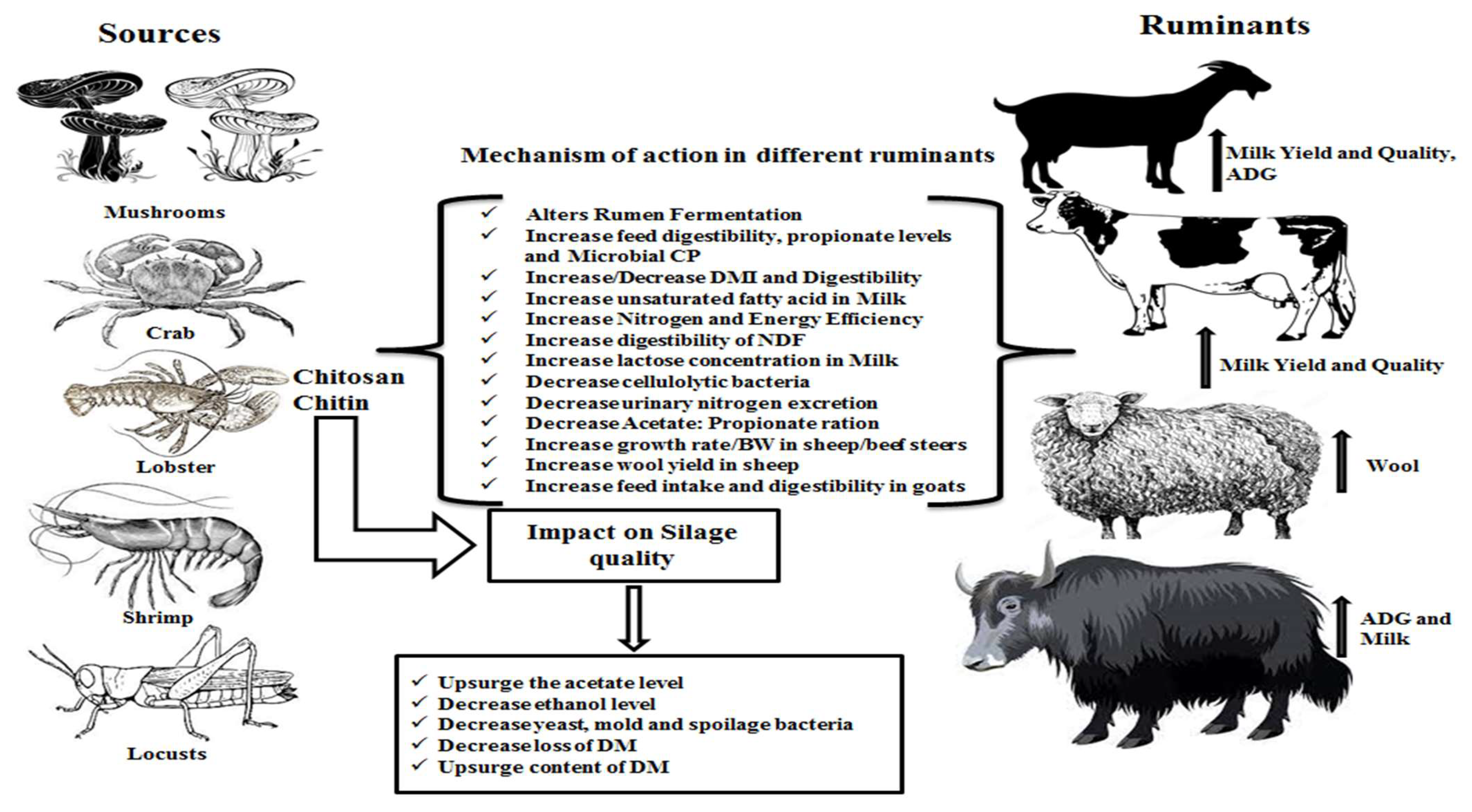
3. Chitin and Chitosan Sources
3.1. Chitin
3.2. Chitosan
4. Modes of Actions in the Rumen on Substrates, Microbiomes, Fermentation, Volatile Fatty Acids, pH, Microbial Protein Synthesis, and Methane Mitigation
4.1. Feed Efficiency, Rumen Fermentation, Volatile Fatty Acid, and Milk Composition and Production
4.2. Bacterial Population and Methane Mitigation
5. Supplementation of Chitin and Chitosan and Its Impact on Performance of Ruminants
6. Advantages of supplementing chitin and chitosan in ruminant diets
7. Future applications of chitin and chitosan in ruminant feeding
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.; Lei, X.; Chen, X.; Yin, Q.; Shen, J.; Yao, J. Long-term and combined effects of N-[2-(nitrooxy) ethyl]-3-pyridinecarboxamide and fumaric acid on methane production, rumen fermentation, and lactation performance in dairy goats. J. Anim. Sci. Biotechnol. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ren, H.; Liu, S.; Cai, C.; Han, J.; Li, F.; Yao, J. Dynamics of methanogenesis, ruminal fermentation, and alfalfa degradation during adaptation to monensin supplementation in goats. J. Dairy Sci. 2018, 101, 1048–1059. [Google Scholar] [CrossRef] [PubMed]
- Koide, S. Chitin-chitosan: Properties, benefits and risks. Nutr. Res. 1998, 18, 1091–1101. [Google Scholar] [CrossRef]
- Singla, A.; Chawla, M. Chitosan: Some pharmaceutical and biological aspects-an update. J. Pharm. Pharmacol. 2001, 53, 1047–1067. [Google Scholar] [CrossRef]
- Swiatkiewicz, S.; Swiatkiewicz, M.; Arczewska-Wlosek, A.; Jozefiak, D. Chitosan and its oligosaccharide derivatives (chito-oligosaccharides) as feed supplements in poultry and swine nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Arachchi, J.K.V.; Jeon, Y.-J. Food applications of chitin and chitosans. Trends Food Sci. Technol. 1999, 10, 37–51. [Google Scholar] [CrossRef]
- Hirano, S.; Itakura, C.; Seino, H.; Akiyama, Y.; Nonaka, I.; Kanbara, N.; Kawakami, T. Chitosan as an ingredient for domestic animal feeds. J. Agric. Food Chem. 1990, 38, 1214–1217. [Google Scholar] [CrossRef]
- Okamoto, Y.; Nose, M.; Miyatake, K.; Sekine, J.; Oura, R.; Shigemasa, Y.; Minami, S. Physical changes of chitin and chitosan in canine gastrointestinal tract. Carbohydr. Polym. 2001, 44, 211–215. [Google Scholar] [CrossRef]
- Lodhi, G.; Kim, Y.-S.; Hwang, J.-W.; Kim, S.-K.; Jeon, Y.-J.; Je, J.-Y.; Ahn, C.-B.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Chitooligosaccharide and its derivatives: Preparation and biological applications. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Zheng, L.-Y.; Zhu, J.-F. Study on antimicrobial activity of chitosan with different molecular weights. Carbohydr. Polym. 2003, 54, 527–530. [Google Scholar] [CrossRef]
- Holappa, J.; Hjálmarsdóttir, M.; Másson, M.; Rúnarsson, Ö.; Asplund, T.; Soininen, P.; Nevalainen, T.; Järvinen, T. Antimicrobial activity of chitosan N-betainates. Carbohydr. Polym. 2006, 65, 114–118. [Google Scholar] [CrossRef]
- Yoon, H.J.; Moon, M.E.; Park, H.S.; Im, S.Y.; Kim, Y.H. Chitosan oligosaccharide (COS) inhibits LPS-induced inflammatory effects in RAW 264.7 macrophage cells. Biochem. Biophys. Res. Commun. 2007, 358, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Liu, H.-T.; Wei, P.; Xu, Q.-S.; Bai, X.-F.; Du, Y.-G.; Yu, C. Chitosan oligosaccharides inhibit LPS-induced over-expression of IL-6 and TNF-α in RAW264. 7 macrophage cells through blockade of mitogen-activated protein kinase (MAPK) and PI3K/Akt signaling pathways. Carbohydr. Polym. 2011, 84, 1391–1398. [Google Scholar] [CrossRef]
- Kim, K.W.; Thomas, R. Antioxidative activity of chitosans with varying molecular weights. Food Chem. 2007, 101, 308–313. [Google Scholar] [CrossRef]
- Shen, K.-T.; Chen, M.-H.; Chan, H.-Y.; Jeng, J.-H.; Wang, Y.-J. Inhibitory effects of chitooligosaccharides on tumor growth and metastasis. Food Chem. Toxicol. 2009, 47, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Nishimura, S.-i.; Nishi, N.; Numata, F.; Tone, Y.; Tokura, S.; Azuma, I. Adjuvant activity of chitin derivatives in mice and guinea-pigs. Vaccine 1985, 3, 379–384. [Google Scholar] [CrossRef]
- Zaharoff, D.A.; Rogers, C.J.; Hance, K.W.; Schlom, J.; Greiner, J.W. Chitosan solution enhances both humoral and cell-mediated immune responses to subcutaneous vaccination. Vaccine 2007, 25, 2085–2094. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, J.; Xia, W. Hypocholesterolaemic effects of different chitosan samples in vitro and in vivo. Food Chem. 2008, 107, 419–425. [Google Scholar] [CrossRef]
- Halmemies-Beauchet-Filleau, A.; Rinne, M.; Lamminen, M.; Mapato, C.; Ampapon, T.; Wanapat, M.; Vanhatalo, A. Alternative and novel feeds for ruminants: Nutritive value, product quality and environmental aspects. Anim. 2018, 12, s295–s309. [Google Scholar] [CrossRef]
- Herrero, M.; Henderson, B.; Havlík, P.; Thornton, P.K.; Conant, R.T.; Smith, P.; Wirsenius, S.; Hristov, A.N.; Gerber, P.; Gill, M. Greenhouse gas mitigation potentials in the livestock sector. Nat. Clim. Change 2016, 6, 452–461. [Google Scholar] [CrossRef]
- Kumirska, J.; Czerwicka, M.; Kaczyński, Z.; Bychowska, A.; Brzozowski, K.; Thöming, J.; Stepnowski, P. Application of spectroscopic methods for structural analysis of chitin and chitosan. Mar. Drugs 2010, 8, 1567–1636. [Google Scholar] [CrossRef] [PubMed]
- Muzzarelli, R.A.; Jeuniaux, C.; Gooday, G.W. Chitin in Nature and Technology; Springer: New York, NY, USA, 1986; Volume 385. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- El Knidri, H.; Belaabed, R.; Addaou, A.; Laajeb, A.; Lahsini, A. Extraction, chemical modification and characterization of chitin and chitosan. Int. J. Biol. Macromol. 2018, 120, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Kurita, K. Controlled functionalization of the polysaccharide chitin. Prog. Polym. Sci. 2001, 26, 1921–1971. [Google Scholar] [CrossRef]
- Pillai, C.K.; Paul, W.; Sharma, C.P. Chitin and chitosan polymers: Chemistry, solubility and fiber formation. Prog. Polym. Sci. 2009, 34, 641–678. [Google Scholar] [CrossRef]
- Kumar, M.N.R. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Petri, D.F.; Donegá, J.; Benassi, A.M.; Bocangel, J.A. Preliminary study on chitosan modified glass ionomer restoratives. Dent. Mater. 2007, 23, 1004–1010. [Google Scholar] [CrossRef]
- Tajik, H.; Moradi, M.; Razavi Rohani, S.M.; Erfani, A.M.; Jalali, F.S.S. Preparation of chitosan from brine shrimp (Artemia urmiana) cyst shells and effects of different chemical processing sequences on the physicochemical and functional properties of the product. Mol. 2008, 13, 1263–1274. [Google Scholar] [CrossRef]
- Muzzarelli, R.; Muzzarelli, C. Chitosan chemistry: Relevance to the biomedical sciences. Polysacch. I 2005, 186, 151–209. [Google Scholar]
- Arai, K. Toxicity of chitosan. Bull. Tokai Reg Fish Res. Lab. 1968, 56, 86–94. [Google Scholar]
- Anggraeni, A.S.; Jayanegara, A.; Laconi, E.B.; Kumalasari, N.R.; Sofyan, A. Marine by-products and insects as a potential chitosan source for ruminant feed additives. Czech. J. Anim. Sci. 2022, 67, 295–317. [Google Scholar] [CrossRef]
- Jiménez-Ocampo, R.; Valencia-Salazar, S.; Pinzón-Díaz, C.E.; Herrera-Torres, E.; Aguilar-Pérez, C.F.; Arango, J.; Ku-Vera, J.C. The role of chitosan as a possible agent for enteric methane mitigation in ruminants. Anim. 2019, 9, 942. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cai, C.; Li, J.; Li, J.; Li, J.; Sun, T.; Wang, L.; Wu, H.; Yu, G. Chitosan-based nanomaterials for drug delivery. Mol. 2018, 23, 2661. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.L.; Khor, E.; Tan, T.K.; Lim, L.Y.; Tan, S.C. Concurrent production of chitin from shrimp shells and fungi. Carbohydr. Res. 2001, 332, 305–316. [Google Scholar] [CrossRef]
- Duffy, C.; O’Riordan, D.; O’Sullivan, M.; Jacquier, J.C. In vitro evaluation of chitosan copper chelate gels as a multimicronutrient feed additive for cattle. J. Sci. Food Agric. 2018, 98, 4177–4183. [Google Scholar] [CrossRef]
- Gandra, J.R.; Takiya, C.S.; Oliveira, E.R.d.; Paiva, P.G.d.; Goes, R.H.d.T.; Gandra, É.R.d.S.; Araki, H.M.C. Nutrient digestion, microbial protein synthesis, and blood metabolites of Jersey heifers fed chitosan and whole raw soybeans. Rev. Bras. De Zootec. 2016, 45, 130–137. [Google Scholar] [CrossRef]
- Naveed, M.; Phil, L.; Sohail, M.; Hasnat, M.; Baig, M.M.F.A.; Ihsan, A.U.; Shumzaid, M.; Kakar, M.U.; Khan, T.M.; Akabar, M. Chitosan oligosaccharide (COS): An overview. Int. J. Biol. Macromol. 2019, 129, 827–843. [Google Scholar] [CrossRef]
- Philibert, T.; Lee, B.H.; Fabien, N. Current status and new perspectives on chitin and chitosan as functional biopolymers. Appl. Biochem. Biotechnol. 2017, 181, 1314–1337. [Google Scholar] [CrossRef]
- Puvvada, Y.S.; Vankayalapati, S.; Sukhavasi, S. Extraction of chitin from chitosan from exoskeleton of shrimp for application in the pharmaceutical industry. Int. Curr. Pharm. J. 2012, 1, 258–263. [Google Scholar] [CrossRef]
- Al Sagheer, F.; Al-Sughayer, M.; Muslim, S.; Elsabee, M. Extraction and characterization of chitin and chitosan from marine sources in Arabian Gulf. Carbohydr. Polym. 2009, 77, 410–419. [Google Scholar] [CrossRef]
- Campana-Filho, S.P.; Britto, D.d.; Curti, E.; Abreu, F.R.; Cardoso, M.B.; Battisti, M.V.; Sim, P.C.; Goy, R.C.; Signini, R.; Lavall, R.L. Extraction, structures and properties of alpha-and beta-chitin. Química Nova 2007, 30, 644–650. [Google Scholar] [CrossRef]
- Noishiki, Y.; Takami, H.; Nishiyama, Y.; Wada, M.; Okada, S.; Kuga, S. Alkali-induced conversion of β-chitin to α-chitin. Biomacromolecules 2003, 4, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Synowiecki, J.; Al-Khateeb, N.A. Production, properties, and some new applications of chitin and its derivatives. Crit Rev Food Sci Nutr. 2003, 43, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, I.; Mengíbar, M.; Harris, R.; Paños, I.; Miralles, B.; Acosta, N.; Galed, G.; Heras, Á. Functional characterization of chitin and chitosan. Curr. Chem. Biol. 2009, 3, 203–230. [Google Scholar]
- Kurita, K.; Tomita, K.; Ishii, S.; Nishimura, S.I.; Shimoda, K. β-chitin as a convenient starting material for acetolysis for efficient preparation of N-acetylchitooligosaccharides. J. Polym. Sci. Part A: Polym. Chem. 1993, 31, 2393–2395. [Google Scholar] [CrossRef]
- Weinhold, M.X.; Sauvageau, J.C.; Keddig, N.; Matzke, M.; Tartsch, B.; Grunwald, I.; Kübel, C.; Jastorff, B.; Thöming, J. Strategy to improve the characterization of chitosan for sustainable biomedical applications: SAR guided multi-dimensional analysis. Green Chem. 2009, 11, 498–509. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Duran, A.; Kahve, H.I. The effect of chitosan coating and vacuum packaging on the microbiological and chemical properties of beef. Meat Sci. 2020, 162, 107961. [Google Scholar] [CrossRef]
- Kirwan, S.F.; Pierce, K.M.; Serra, E.; McDonald, M.; Rajauria, G.; Boland, T.M. Effect of chitosan inclusion and dietary crude protein level on nutrient intake and digestibility, ruminal fermentation, and N excretion in beef heifers offered a grass silage based diet. Anim. 2021, 11, 771. [Google Scholar] [CrossRef]
- Zanferari, F.; Vendramini, T.H.A.; Rentas, M.F.; Gardinal, R.; Calomeni, G.D.; Mesquita, L.G.; Takiya, C.S.; Rennó, F.P. Effects of chitosan and whole raw soybeans on ruminal fermentation and bacterial populations, and milk fatty acid profile in dairy cows. J. Dairy Sci. 2018, 101, 10939–10952. [Google Scholar] [CrossRef]
- Del Valle, T.A.; de Paiva, P.G.; de Jesus, E.F.; de Almeida, G.F.; Zanferari, F.; Costa, A.G.; Bueno, I.C.; Rennó, F.P. Dietary chitosan improves nitrogen use and feed conversion in diets for mid-lactation dairy cows. Livest. Sci. 2017, 201, 22–29. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.; Garcia-Rodriguez, A. Use of chitosans to modulate ruminal fermentation of a 50: 50 forage-to-concentrate diet in sheep. J. Anim. Sci. 2010, 88, 749–755. [Google Scholar] [CrossRef]
- Araújo, A.P.C.d.; Venturelli, B.C.; Santos, M.C.B.; Gardinal, R.; Cônsolo, N.R.B.; Calomeni, G.D.; Freitas, J.; Barletta, R.V.; Gandra, J.R.; Paiva, P. Chitosan affects total nutrient digestion and ruminal fermentation in Nellore steers. Anim. Feed. Sci. Technol. 2015, 206, 114–118. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.; Garcia-Rodriguez, A. Dose–response effects of chitosans on in vitro rumen digestion and fermentation of mixtures differing in forage-to-concentrate ratios. Anim. Feed. Sci. Technol. 2009, 151, 215–227. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and mode of action of chitosan and ivy fruit saponins on the microbiome, fermentation and methanogenesis in the rumen simulation technique. FEMS Microbiol. Ecol. 2016, 92, fiv160. [Google Scholar] [CrossRef]
- Dias, A.; Goes, R.; Gandra, J.; Takiya, C.; Branco, A.; Jacaúna, A.; Oliveira, R.; Souza, C.; Vaz, M. Increasing doses of chitosan to grazing beef steers: Nutrient intake and digestibility, ruminal fermentation, and nitrogen utilization. Anim. Feed. Sci. Technol. 2017, 225, 73–80. [Google Scholar] [CrossRef]
- de Paiva, P.G.; de Jesus, E.F.; Del Valle, T.A.; de Almeida, G.F.; Costa, A.G.B.V.B.; Consentini, C.E.C.; Zanferari, F.; Takiya, C.S.; da Silva Bueno, I.C.; Rennó, F.P. Effects of chitosan on ruminal fermentation, nutrient digestibility, and milk yield and composition of dairy cows. Anim. Prod. Sci. 2016, 57, 301–307. [Google Scholar] [CrossRef]
- Mingoti, R.D.; Freitas Jr, J.; Gandra, J.R.; Gardinal, R.; Calomeni, G.D.; Barletta, R.V.; Vendramini, T.H.A.; Paiva, P.; Rennó, F.P. Dose response of chitosan on nutrient digestibility, blood metabolites and lactation performance in holstein dairy cows. Livest. Sci. 2016, 187, 35–39. [Google Scholar] [CrossRef]
- Vendramini, T.; Takiya, C.; Silva, T.; Zanferari, F.; Rentas, M.; Bertoni, J.; Consentini, C.; Gardinal, R.; Acedo, T.; Rennó, F. Effects of a blend of essential oils, chitosan or monensin on nutrient intake and digestibility of lactating dairy cows. Anim. Feed. Sci. Technol. 2016, 214, 12–21. [Google Scholar] [CrossRef]
- Jiménez-Ocampo, R.; Montoya-Flores, M.D.; Herrera-Torres, E.; Pámanes-Carrasco, G.; Arceo-Castillo, J.I.; Valencia-Salazar, S.S.; Arango, J.; Aguilar-Pérez, C.F.; Ramírez-Avilés, L.; Solorio-Sánchez, F.J. Effect of chitosan and naringin on enteric methane emissions in crossbred heifers fed tropical grass. Anim. 2021, 11, 1599. [Google Scholar] [CrossRef]
- Wencelova, M.; Varadyova, Z.; Mihalikova, K.; Kisidayova, S.; Jalc, D. Evaluating the effects of chitosan, plant oils, and different diets on rumen metabolism and protozoan population in sheep. Turk. J. Vet. Anim. Sci. 2014, 38, 26–33. [Google Scholar] [CrossRef]
- Bach, A.; Calsamiglia, S.; Stern, M. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef]
- Ivan, M. Comparison of duodenal flow and digestibility in fauna-free sheep inoculated with Holotrich protozoa, Entodinium monofauna or total mixed protozoa population. Br. J. Nutr. 2008, 101, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Mould, F.; Ørskov, E.; Gauld, S.A. Associative effects of mixed feeds. II. The effect of dietary addition of bicarbonate salts on the voluntary intake and digestibility of diets containing various proportions of hay and barley. Anim. Feed. Sci. Technol. 1983, 10, 31–47. [Google Scholar] [CrossRef]
- Dijkstra, J.; Ellis, J.; Kebreab, E.; Strathe, A.; López, S.; France, J.; Bannink, A. Ruminal pH regulation and nutritional consequences of low pH. Anim. Feed. Sci. Technol. 2012, 172, 22–33. [Google Scholar] [CrossRef]
- Colmenero, J.O.; Broderick, G.A. Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci. 2006, 89, 1704–1712. [Google Scholar] [CrossRef]
- Sannes, R.; Messman, M.; Vagnoni, D. Form of rumen-degradable carbohydrate and nitrogen on microbial protein synthesis and protein efficiency of dairy cows. J. Dairy Sci. 2002, 85, 900–908. [Google Scholar] [CrossRef]
- Kang-Meznarich, J.; Broderick, G. Effects of incremental urea supplementation on ruminal ammonia concentration and bacterial protein formation. J. Anim. Sci. 1980, 51, 422–431. [Google Scholar] [CrossRef]
- Chen, H.-C.; Chang, C.-C.; Mau, W.-J.; Yen, L.-S. Evaluation of N-acetylchitooligosaccharides as the main carbon sources for the growth of intestinal bacteria. FEMS Microbiol. Lett. 2002, 209, 53–56. [Google Scholar] [CrossRef][Green Version]
- Haraki, H.; Gandra, J.; Oliveira, E.; Takiya, C.; Goes, R.; Gabriel, A.; Rodrigues, G.; Gandra, E.; Pereira, T.; Damiani, J. Effects of chitosan and whole raw soybeans on feeding behavior and heat losses of Jersey heifers. Iran. J. Appl. Anim. Sci. 2018, 8, 397–405. [Google Scholar]
- Baldwin, R.t.; McLeod, K.; McNamara, J.; Elsasser, T.; Baumann, R. Influence of abomasal carbohydrates on subcutaneous, omental, and mesenteric adipose lipogenic and lipolytic rates in growing beef steers. J. Anim. Sci. 2007, 85, 2271–2282. [Google Scholar] [CrossRef] [PubMed]
- Şenel, S.; McClure, S.J. Potential applications of chitosan in veterinary medicine. Adv. Drug Deliv. Rev. 2004, 56, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.J.; Oh, M.; Yeo, W.-S.; Galvao, K.N.; Jeong, K.C. Underlying mechanism of antimicrobial activity of chitosan microparticles and implications for the treatment of infectious diseases. PLoS ONE 2014, 9, e92723. [Google Scholar] [CrossRef] [PubMed]
- Daetz, R.; Cunha, F.; Bittar, J.; Risco, C.; Magalhaes, F.; Maeda, Y.; Santos, J.; Jeong, K.; Cooke, R.; Galvão, K. Clinical response after chitosan microparticle administration and preliminary assessment of efficacy in preventing metritis in lactating dairy cows. J. Dairy Sci. 2016, 99, 8946–8955. [Google Scholar] [CrossRef]
- Lanctôt, S.; Fustier, P.; Taherian, A.; Bisakowski, B.; Zhao, X.; Lacasse, P. Effect of intramammary infusion of chitosan hydrogels at drying-off on bovine mammary gland involution. J. Dairy Sci. 2017, 100, 2269–2281. [Google Scholar] [CrossRef]
- Ipharraguerre, I.R.; Clark, J.H. Usefulness of ionophores for lactating dairy cows: A review. Anim. Feed. Sci. Technol. 2003, 106, 39–57. [Google Scholar] [CrossRef]
- Lourenço, M.; Ramos-Morales, E.; Wallace, R. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Anim. 2010, 4, 1008–1023. [Google Scholar] [CrossRef]
- Relling, A.; Reynolds, C. Feeding rumen-inert fats differing in their degree of saturation decreases intake and increases plasma concentrations of gut peptides in lactating dairy cows. J. Dairy Sci. 2007, 90, 1506–1515. [Google Scholar] [CrossRef]
- Barletta, R.V.; Gandra, J.R.; Bettero, V.P.; Araújo, C.E.; Del Valle, T.A.; de Almeida, G.F.; de Jesus, E.F.; Mingoti, R.D.; Benevento, B.C.; de Freitas Júnior, J.E. Ruminal biohydrogenation and abomasal flow of fatty acids in lactating cows: Oilseed provides ruminal protection for fatty acids. Anim. Feed. Sci. Technol. 2016, 219, 111–121. [Google Scholar] [CrossRef]
- NRC, I. Nutrient requirements of dairy cattle. Natl. Res. Counc. 2001, 519. [Google Scholar]
- Yang, S.; Bu, D.; Wang, J.; Hu, Z.; Li, D.; Wei, H.; Zhou, L.; Loor, J. Soybean oil and linseed oil supplementation affect profiles of ruminal microorganisms in dairy cows. Anim. 2009, 3, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, J.; Li, L.; Xia, W. Dietary chitosan improves hypercholesterolemia in rats fed high-fat diets. Nutr. Res. 2008, 28, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.I.; No, H.K.; Meyers, S.P. Physicochemical characteristics and functional properties of various commercial chitin and chitosan products. J. Agric. Food Chem. 1998, 46, 3839–3843. [Google Scholar] [CrossRef]
- Razdan, A.; Pettersson, D. Effect of chitin and chitosan on nutrient digestibility and plasma lipid concentrations in broiler chickens. Br. J. Nutr. 1994, 72, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.; Cai, Y.; Corner, R.; Morvan, B.; Joblin, K. Identification and enumeration of oleic acid and linoleic acid hydrating bacteria in the rumen of sheep and cows. J. Appl. Microbiol. 2000, 88, 286–292. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Sakurada, M.; Tomita, Y.; Onodera, R. Presence in rumen bacterial and protozoal populations of enzymes capable of degrading fungal cell walls. Microbiol. 1994, 140, 631–636. [Google Scholar] [CrossRef][Green Version]
- Kopečný, J.; Hodrová, B. Chitinolytic enzymes produced by ovine rumen bacteria. Folia Microbiol. 2000, 45, 465–468. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. De Zootech. INRA/EDP Sci. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Oba, M.; Allen, M.S. Intraruminal infusion of propionate alters feeding behavior and decreases energy intake of lactating dairy cows. J. Nutr. 2003, 133, c–1099. [Google Scholar] [CrossRef]
- Sudarshan, N.; Hoover, D.; Knorr, D. Antibacterial action of chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Wu, S. Preparation of water soluble chitosan by hydrolysis with commercial α-amylase containing chitosanase activity. Food Chem. 2011, 128, 769–772. [Google Scholar] [CrossRef]
- Da Silva, D.; Santos, G.; Branco, A.; Damasceno, J.; Kazama, R.; Matsushita, M.; Horst, J.; Dos St., W.; Petit, H. Production performance and milk composition of dairy cows fed whole or ground flaxseed with or without monensin. J. Dairy Sci. 2007, 90, 2928–2936. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.D. Antibiotic use in animal feed and its impact on human healt. Nutr. Res. Rev. 2000, 13, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Henry, D.; Ruiz-Moreno, M.; Ciriaco, F.; Kohmann, M.; Mercadante, V.; Lamb, G.; DiLorenzo, N. Effects of chitosan on nutrient digestibility, methane emissions, and in vitro fermentation in beef cattle. J. Anim. Sci. 2015, 93, 3539–3550. [Google Scholar] [CrossRef]
- Pereira, F.M.; Magalhães, T.d.S.; de Freitas Júnior, J.E.; Santos, S.A.; Pinto, L.F.; Pina, D.d.S.; Mourão, G.B.; Pires, A.J.; Júnior, F.J.C.; Mesquita, B.M.d.C. Qualitative profile of meat from lambs fed diets with different levels of chitosan. Part II. Livest. Sci. 2022, 104975. [Google Scholar] [CrossRef]
- Seankamsorn, A.; Cherdthong, A.; So, S.; Wanapat, M. Influence of chitosan sources on intake, digestibility, rumen fermentation, and milk production in tropical lactating dairy cows. Trop. Anim. Health Prod. 2021, 53, 1–9. [Google Scholar] [CrossRef]
- AY, A.-E.; Azab, M.; Esmail, R.; Nafeaa, A.; Shousha, S. The Effects of Live Yeast (Saccharomyces cerevisiae) and Chitosan on Rumen Fermentation Function and Rumen Development in Male Balady Goats. Benha Vet. Med. J. 2019, 37, 59–65. [Google Scholar]
- Gorelik, V.; Rebezov, M.; Lopaeva, N.; Smirnova, E.; Sultanova, S. Morphological and biochemical parameters of cow blood when using chitosan preparations. Proc. E3S Web Conf. 2021, 254, 08025. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, B.; Qi, J.; Zhao, Y.; Guo, X.; Shi, B.; Yan, S. Dietary supplementation of chitosan affects milk performance, markers of inflammatory response and antioxidant status in dairy cows. Anim. Feed. Sci. Technol. 2021, 277, 114952. [Google Scholar] [CrossRef]
- Zhang, Q.; Han, Y.; Yang, Y.; Shen, X. Effects of the seleno-chitosan on daily gain, wool yield, and blood parameter in the Chinese merino sheep. Biol. Trace Elem. Res. 2022, 200, 4704–4711. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.; Carvalho, G.; Magalhães, T.; Júnior, J.F.; Pinto, L.; Mourão, G.B.; Pires, A.; Eiras, C.; Novais-Eiras, D.; Azevêdo, J. Effect of chitosan on production performance of feedlot lambs. J. Agric. Sci. 2018, 156, 1138–1144. [Google Scholar] [CrossRef]
- Jayanegara, A.; Novandri, B.; Yantina, N.; Ridla, M. Use of black soldier fly larvae (Hermetia illucens) to substitute soybean meal in ruminant diet: An in vitro rumen fermentation study. Vet. World 2017, 10, 1439–1446. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Chitosan/Chitin | Trial Animal Model and Duration | Dose | Substrate/Feed | Results | Reference |
|---|---|---|---|---|---|
| Chitosan | Sheep lambs Santa | 0, 136, and 272 mg/kg of BW | Diet | No effect on feed intake, BW, and FC. Improved pH and fatty acid composition in meat, improved meat quality | [97] |
| 90% deacetylation | Dairy cows | 0 and 2% | TMR | No effect on nutrient digestibility, affect rumen fermentation pattern, reduced methane production | [98] |
| Chitosan | Balady male goats (120 days) | 0.2% (2 kg/ton concentrate) | Concentrate diet 3% | Chitosan increases ruminal ammonia nitrogen, no effect on VFA levels, no effect on total protozoal count | [99] |
| Chitosan sucinate deac.75% | black and white breed dairy cow | 0% and 2% | Feed | Improved blood physiological condition and metabolism | [100] |
| Chitosan Deac. 85% | Dairy cows | 0.500, 1000, 1500, 2000 mg/kg | TMR | Improved DMI intake, milk production, antioxidant capacity, | [101] |
| Chitosan | Sheep (45 days) | 0 and 136 mg/kg of BW | Alfalfa hay and concentrate at 50:50 | Chitosan reduce NDF apparent digestibility, ruminal NH3-N concentration and modulates ruminal and fecal fermentative activity | [52] |
| Chitosan | Feedlot lambs Santa Inês crossbred sheep (90 days) | 136 mg and 272 mg chitosan/kg BW | Roughage to concentrate ratio at 50:50 | Chitosan did not affect the DM intake, improved digestibility. No effect was observed on weight gain, carcass weight not influenced | [102] |
| Chitosan | dairy cow (84 days) | 0, 50, 100 and 150 mg/kg BW | Corn Silage-concentrate 60:40 | Chitosan shifted rumen fermentation, improved nutrient digestibility andpropionate concentrations | [53] |
| Chitosan | dairy cow (84 days) | 0 and 4 g/kg ofDM | Corn silage-to concentrateratio50:50 | Improved feed e ciency, increased milk UFAconcentration | [51] |
| Chitosan | Cattle (25 days) | 0, 2.0 g/kg chitosan (CH)of DM. Wholeraw soybean(WRS) 163.0 g/kg DM; andCH + WRS | Corn silage toconcentrate ratio50:50 | Chitosan improved nutrient digestion anddecrease DMI and reduce nitrogen excreted infeces | [26] |
| Chitosan | Cattle (105 days) | 0, 400, 800, 1200 or 1600 mg/kg DM | Grazing Urochloabrizantha andconcentrate at 150 g/100 kg of LW | Chitosan increased DMI and digestibility, propionate concentration and microbial crudeprotein | [56] |
| Chitosan | Dairy cow (84 days) | 50, 100 and 150 mg/kg BW | Corn silage toconcentrate ratio50:50 | Improved nutrient digestibility withoutaltering productive performance of dairy cows | [58] |
| Chitosan | dairy cow (98 days) | 0, 75, 150, 225 mg/kg BW | Corn silage toconcentrate ratio63:37 | In dairy cattle works like a modulator ofrumen fermentation, increasing milk yield, propionate and nitrogen utilization | [51] |
| Chitosan | Cattle (21 days) | 0.0, 0.5, and 1.0% of DM | High-concentrate (85%) Lowconcentrate (36%) | In vivo: No effect on enteric methaneemissions. In vitro: Low concentrate substrateincreased methane production | [95] |
| Chitosan deace. 95% | HF cross 10 days | 10 g kg-1 DM | TMR | No effect on nitrogen excretion, reduced nutrient digestibility | [50] |
| Chitosan deace 92% | Cross breed Heifers | 0, 1.5, 3 g/kg DMI | TMR | No effect on rumen fermentation, methane production | [61] |
| Chitosan | Cattle (84 days) | 150 mg/kg BW | Maize silage:concentrate ratio50:50 | Chitosan increase the digestibility and reduceacetate to propionate relation | [59] |
| Chitosan | Dairy cow (92 days) | 0 or 4 g/kgchitosan (CH)or Whole RawSoybean(WRS) of DM | Corn silage:concentrate ratio50:50 | Improved ruminal fermentation, increased milk content of UFA, decreasesnutrient intake, digestibility, microbial proteinsynthesis, and milk yield. CH in diets with nolipid supplementation improves feedefficiency of lactating cows | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, A.M.; Qazi, I.H.; Matra, M.; Wanapat, M. Role of Chitin and Chitosan in Ruminant Diets and Their Impact on Digestibility, Microbiota and Performance of Ruminants. Fermentation 2022, 8, 549. https://doi.org/10.3390/fermentation8100549
Shah AM, Qazi IH, Matra M, Wanapat M. Role of Chitin and Chitosan in Ruminant Diets and Their Impact on Digestibility, Microbiota and Performance of Ruminants. Fermentation. 2022; 8(10):549. https://doi.org/10.3390/fermentation8100549
Chicago/Turabian StyleShah, Ali Mujtaba, Izhar Hyder Qazi, Maharach Matra, and Metha Wanapat. 2022. "Role of Chitin and Chitosan in Ruminant Diets and Their Impact on Digestibility, Microbiota and Performance of Ruminants" Fermentation 8, no. 10: 549. https://doi.org/10.3390/fermentation8100549
APA StyleShah, A. M., Qazi, I. H., Matra, M., & Wanapat, M. (2022). Role of Chitin and Chitosan in Ruminant Diets and Their Impact on Digestibility, Microbiota and Performance of Ruminants. Fermentation, 8(10), 549. https://doi.org/10.3390/fermentation8100549