
Screening of Poly-Glutamic Acid (PGA)-Producing Bacillus Species from Indian Fermented Soybean Foods and Characterization of PGA

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
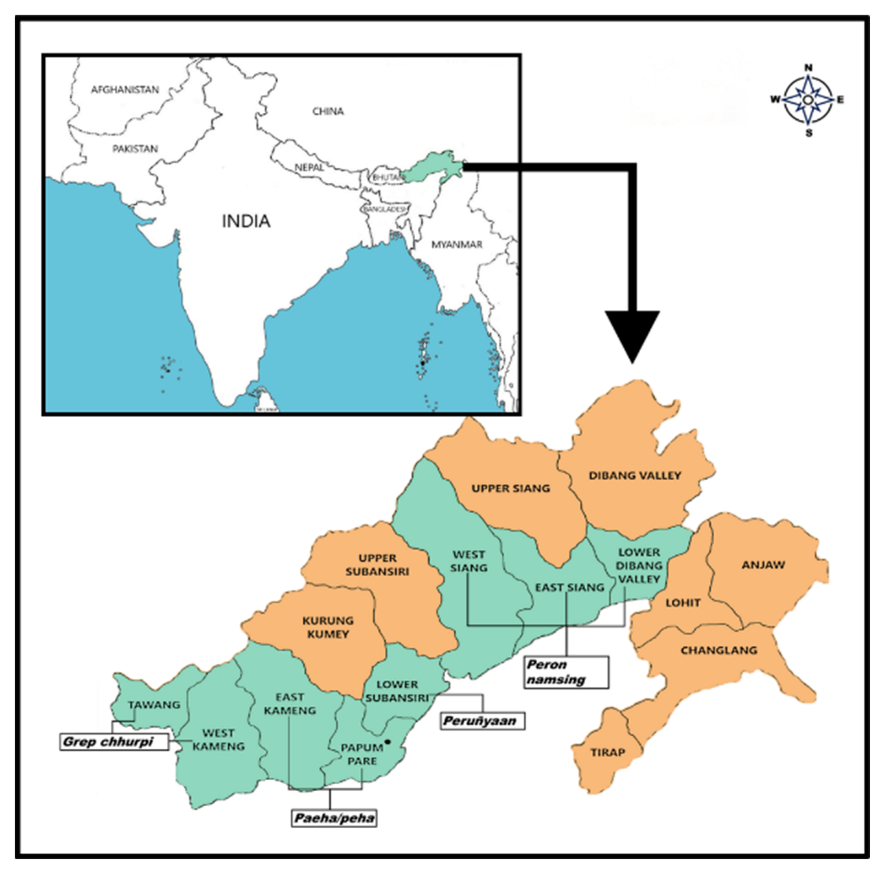
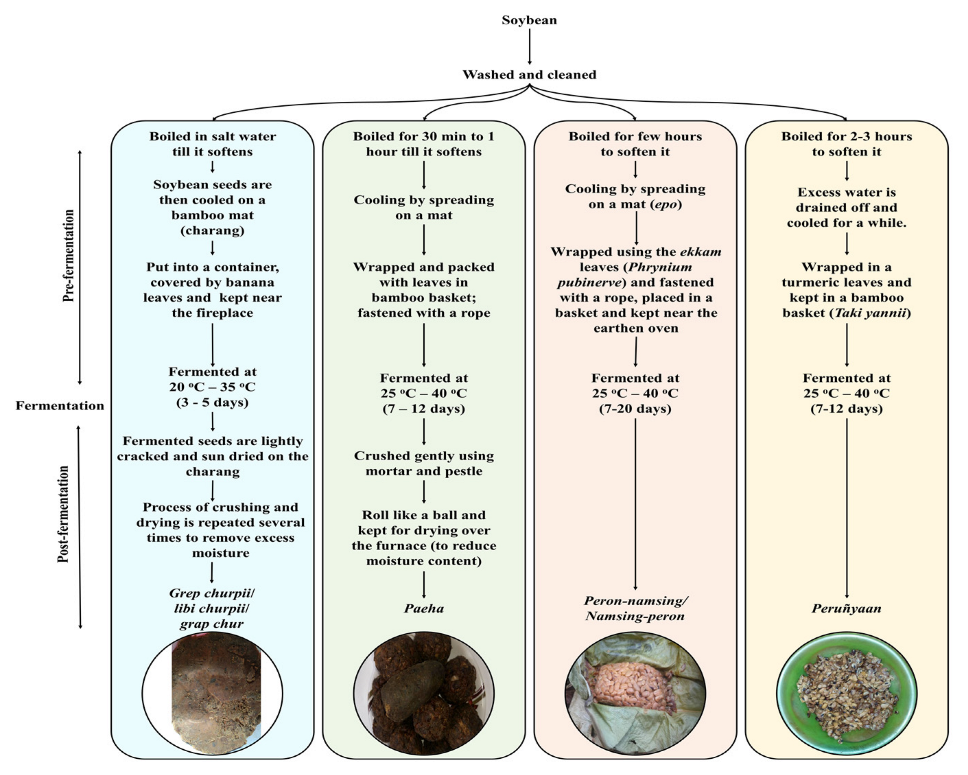
2.1. Collection of Samples
2.2. Analysis of pH
2.3. Isolation of Bacteria
2.4. Measurement of Stickiness
2.5. Measurement of Viscosity
2.6. Phenotypic Characteristics
2.7. Genotypic Identification
2.7.1. Genomic DNA Isolation
2.7.2. PCR Amplification
2.7.3. Purification of PCR Amplicons
2.7.4. 16S rRNA Gene Sequencing
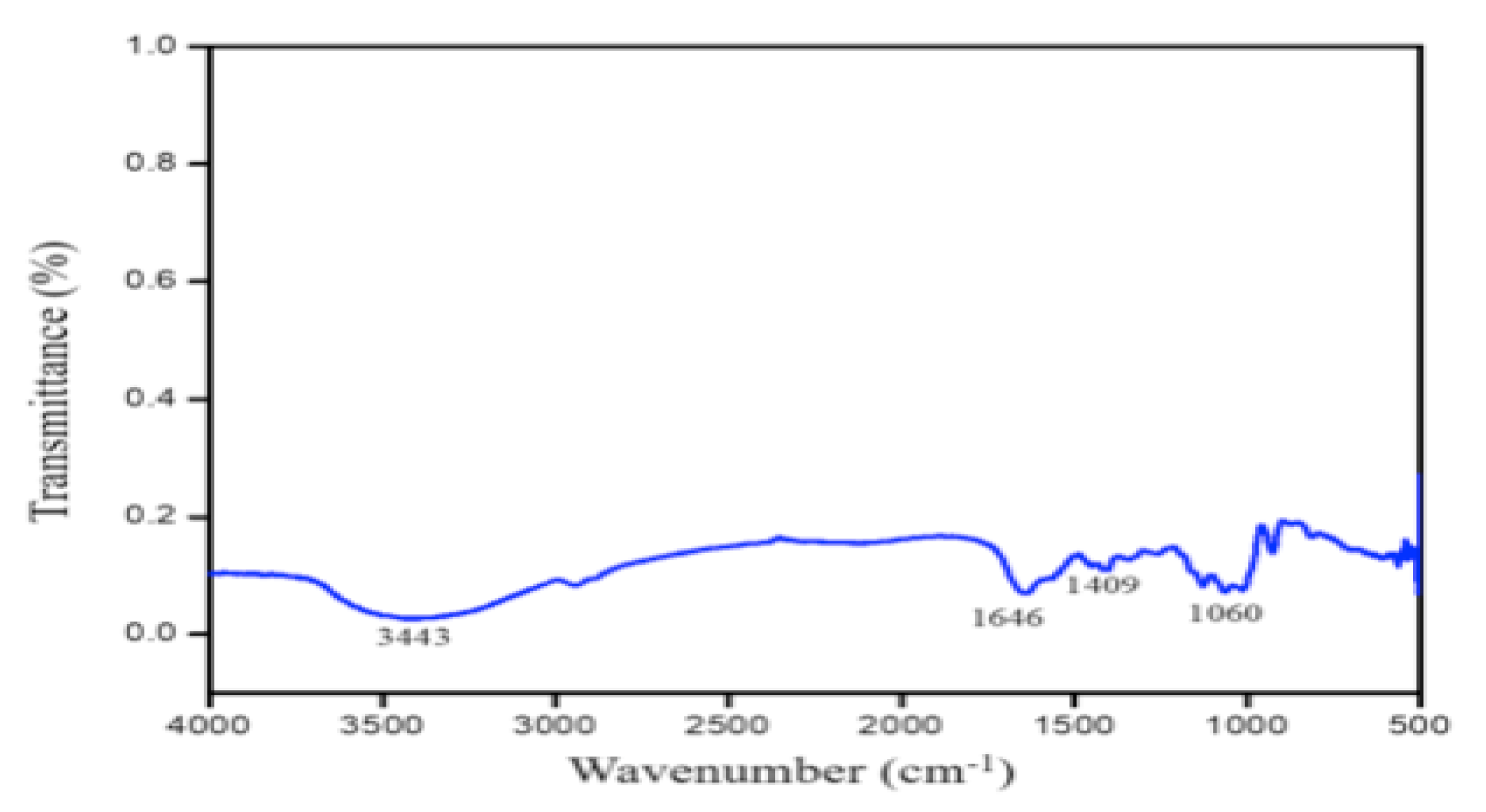
2.8. Characterization of PGA
2.8.1. Extraction of PGA from Bacterial Isolates
2.8.2. Amino Acid Analysis
2.8.3. Gel Filtration Analysis
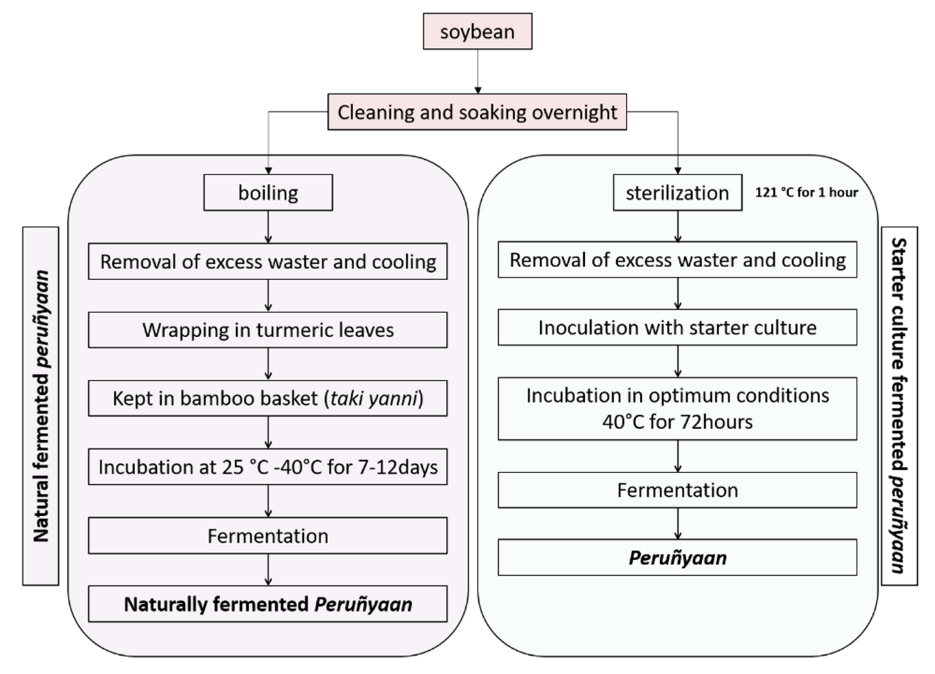
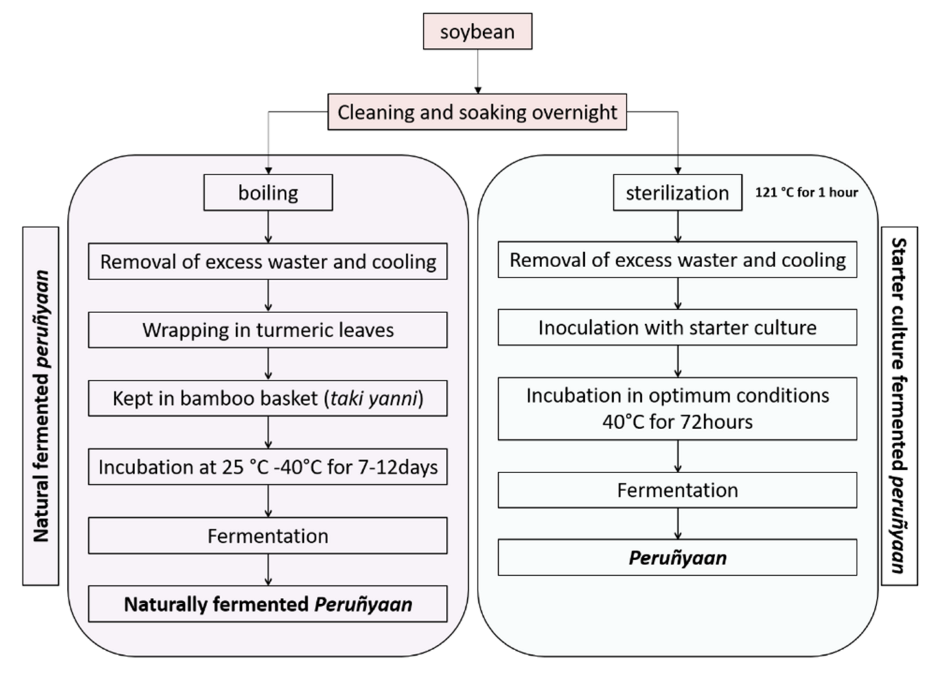
2.9. Preparation of Peruñyaan Using Bacillus safensis Subsp. safensis PY1-19
PGA Production
2.10. Characterization of B. safensis Subsp. safensis PY1-19
2.10.1. Hemolysis
2.10.2. Phytase Activity
2.10.3. Antagonistic Properties against Pathogenic Bacteria
2.11. Bioinformatics Analysis
2.12. Statistics and Visualization
3. Results
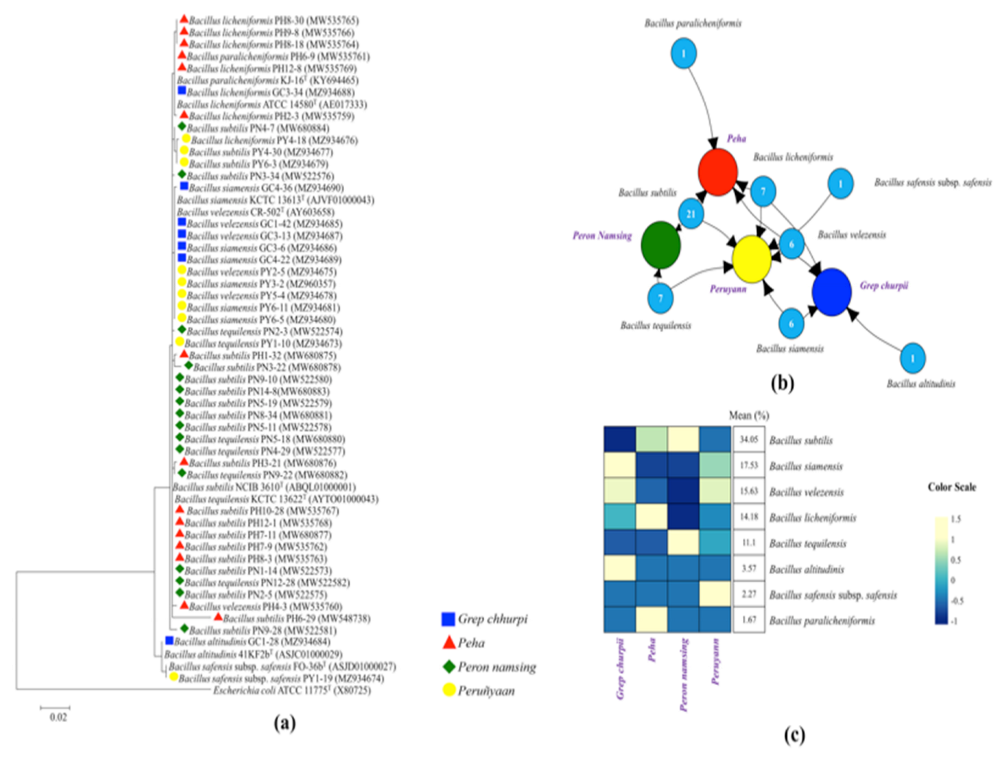
3.1. Bacillus Species Diversity
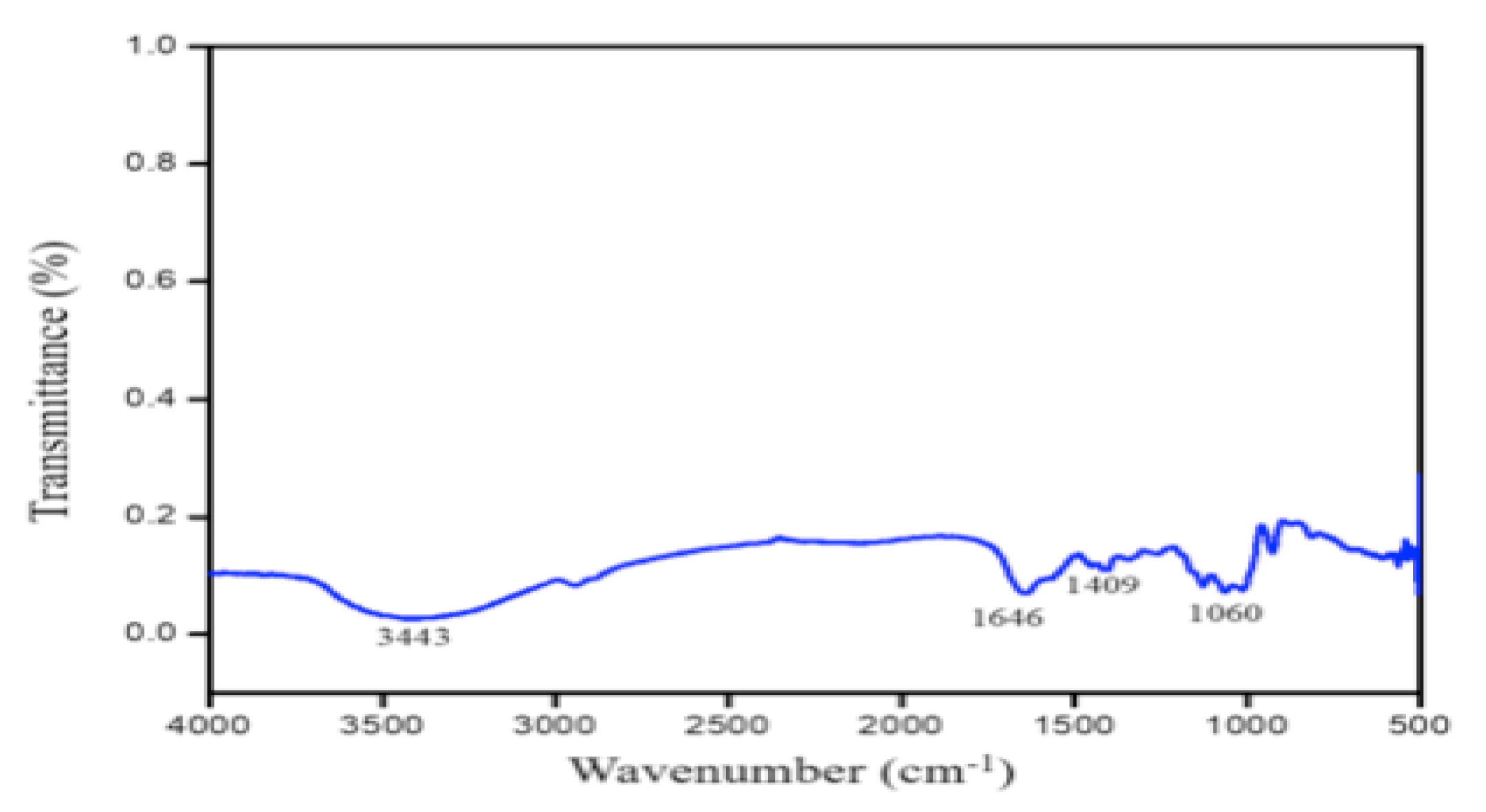
3.2. Characterization of PGA
3.3. Selection of High-PGA-Producing B. safensis subsp. safensis PY1-19
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tamang, J.P.; Cotter, P.D.; Endo, A.; Han, N.S.; Kort, R.; Liu, S.Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented foods in a global age: East meets West. Compr. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Gallone, B.; Voordeckers, K.; Verstrepen, K.J. Domestication of industrial microbes. Curr. Biol. 2019, 29, R381–R393. [Google Scholar] [CrossRef]
- Tamang, J.P.; Holzapfel, W.H.; Watanabe, K. Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P. Naturally fermented ethnic soybean foods of India. J. Ethn. Foods 2015, 2, 8–17. [Google Scholar] [CrossRef]
- Hartley, I.E.; Liem, D.G.; Keast, R. Umami as an ‘alimentary’ taste. a new perspective on taste classification. Nutrients 2019, 11, 182. [Google Scholar] [CrossRef]
- Tamang, J.P. Dietary culture and antiquity of the Himalayan fermented foods and alcoholic fermented beverages. J. Ethn. Foods 2022, 9, 30. [Google Scholar] [CrossRef]
- Tamang, J.P.; Nikkuni, S. Selection of starter culture for production of kinema, fermented soybean food of the Himalaya. World J. Microbiol. Biotechnol. 1996, 12, 629–635. [Google Scholar] [CrossRef]
- Chettri, R.; Bhutia, M.O.; Tamang, J.P. Poly-γ-glutamic acid (PGA)-producing Bacillus species isolated from Kinema, Indian fermented soybean food. Front. Microbiol. 2016, 7, 971. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Z.; Li, S.; Tian, Z.; Ma, X. Study on the mechanism of production of γ-PGA and nattokinase in Bacillus subtilis natto based on RNA-seq analysis. Microb. Cell Fact. 2021, 20, 83. [Google Scholar] [CrossRef]
- Kharnaior, P.; Tamang, J.P. Metagenomic–metabolomic mining of kinema, a naturally fermented soybean food of the Eastern Himalayas. Front. Microbiol. 2022, 13, 868383. [Google Scholar] [CrossRef]
- Tamang, J.P.; Das, D.; Kharnaior, P.; Pariyar, P.; Thapa, N.; Jo, S.W.; Yim, E.J.; Shin, D.H. Shotgun metagenomics of cheonggukjang, a fermented soybean food of Korea, community structure, predictive functionalities and amino acids profile. Food Res. Int. 2022, 151, 110904. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.; Navale, G.R.; Dharne, M.S. Poly-gamma-glutamic acid biopolymer: A sleeping giant with diverse applications and unique opportunities for commercialization. Biomass Convers. Biorefinery 2021, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Kim, J.H.; Kim, K.W.; Lee, B.J.; Kim, D.G.; Kim, Y.O.; Lee, J.H.; Kong, I.S. Physicochemical properties, production, and biological functionality of poly-γ-d-glutamic acid with constant molecular weight from halotolerant Bacillus sp. SJ-10. Int. J. Biol. Macromol. 2018, 108, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Ajayeoba, T.A.; Dula, S.; Ijabadeniyi, O.A. Properties of poly-γ-glutamic acid producing-Bacillus species isolated from Ogi liquor and lemon-Ogi liquor. Front. Microbiol. 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Ratha, P.; Deok-Young, J. Factors increasing poly-γ-glutamic acid content of cheonggukjang fermented by Bacillus subtilis 168. Food Sci. Biotechnol. 2019, 28, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Araki, R.; Fujie, K.; Yuine, N.; Watabe, Y.; Maruo, K.; Suzuki, H.; Hashimoto, K. The possibility of suppression of increased postprandial blood glucose levels by gamma-polyglutamic acid-rich natto in the early phase after eating: A randomized crossover pilot study. Nutrients 2020, 12, 915. [Google Scholar] [CrossRef]
- Lee, N.R.; Go, T.H.; Lee, S.M.; Jeong, S.Y.; Park, G.T.; Hong, C.O.; Son, H.J. In vitro evaluation of new functional properties of poly-γ-glutamic acid produced by Bacillus subtilis D7. Saudi J. Biol. Sci. 2014, 21, 153–158. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Wong, S.K.; Kezuka, M.; Oshiro, N.; Chan, H.T. Natto and miso: An overview on their preparation, bioactive components and health-promoting effects. Food Res. 2021, 5, 446–452. [Google Scholar] [CrossRef]
- Tamang, J.P.; Kharnaior, P.; Pariyar, P.; Thapa, N.; Lar, N.; Win, K.S.; Mar, A.; Nyo, N. Shotgun sequence- based metataxonomic and predictive functional profiles of Pe poke, a naturally fermented soybean food of Myanmar. PLoS ONE 2021, 16, e0260777. [Google Scholar] [CrossRef]
- Tork, S.E.; Aly, M.M.; Alakilli, S.Y.; Al-Seeni, M.N. Purification and characterization of gamma poly glutamic acid from newly Bacillus licheniformis NRC20. Int. J. Biol. Macromol. 2015, 74, 382–391. [Google Scholar] [CrossRef]
- Shangpliang, H.N.J.; Tamang, J.P. Phenotypic and genotypic characterisation of lactic acid bacteria isolated from exotic naturally fermented milk (cow and yak) products of Arunachal Pradesh, India. Int. Dairy J. 2021, 118, 105038. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Anju, A.J.; Binod, P.; Pandey, A. Production and characterization of microbial poly-γ-glutamic acid from renewable resources. Indian J. Experimen. Biol. 2017, 55, 405–410. [Google Scholar]
- Longvah, T.; Ananthan, R.; Bhaskarachary, K.; Venkaiah, K. Indian Food Composition Tables; National Institute of Nutrition, Hyderabad, Indian Council of Medical Research; Department of Health Research, Ministry of Health and Family Welfare, Government of India: Hyderabad, India, 2017. [Google Scholar]
- Tamang, J.P. Development of pulverised starter for kinema production. J. Food Sci. Technol. 1999, 36, 475–478. [Google Scholar]
- Mohanraj, R.; Gnanamangai, B.M.; Ramesh, K.; Priya, P.; Srisunmathi, R.; Poornima, S.; Ponmurugan, P.; Robinson, J.P. Optimized production of gamma poly glutamic acid (γ-PGA) using sago. Biocatal. Agric. Biotechnol. 2019, 22, 101413. [Google Scholar] [CrossRef]
- Lee, S.G.; Chang, H.C. Assessment of Bacillus subtilis SN7 as a starter culture for Cheonggukjang, a Korean traditional fermented soybean food, and its capability to control Bacillus cereus in Cheonggukjang. Food Contr. 2017, 73, 946–953. [Google Scholar] [CrossRef]
- Verma, S.; Kumar, M.; Kumar, A.; Das, S.; Chakdar, H.; Varma, A.; Saxena, A.K. Diversity of bacterial endophytes of maize (Zea Mays) and their functional potential for micronutrient biofortification. Curr. Microbiol. 2022, 79, 1–14. [Google Scholar] [CrossRef]
- Hallaj-Nezhadi, S.; Hamdipour, R.; Shahrvirani, M.; Tin, R.Z.; Chapeland-Leclerc, F.; Ruprich-Robert, G.; Esnaashari, S.; Elyasi Far, B.; Dilmaghani, A. Antimicrobial activity of Bacillus sp. isolated strains of wild honey. BMC Complement. Med. Ther. 2022, 22, 78. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Willis, A.D. Rarefaction, alpha diversity, and statistics. Front. Microbiol. 2019, 10, 2407. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P. “Ethno-Microbiology” of ethnic Indian fermented foods and alcoholic beverages. J. Appl. Microbiol. 2022, 133, 145–161. [Google Scholar] [CrossRef]
- Chettri, R.; Tamang, J.P. Bacillus species isolated from tungrymbai and bekang, naturally fermented soybean foods of India. Int. J. Food Microbiol. 2015, 197, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Kharnaior, P.; Tamang, J.P. Bacterial and fungal communities and their predictive functional profiles in kinema, a naturally fermented soybean food of India, Nepal and Bhutan. Food Res. Int. 2021, 140, 110055. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, D.; Heo, G.; Kong, H.; Kim, C.; Lee, J. Genetic background behind the amino acid profiles of fermented soybeans produced by four Bacillus spp. J. Microbiol. Biotechnol. 2021, 31, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Sopheap, E.; Thavrak, H.; Borarin, B.; Chim, C.; Inatsu, Y. Application of antimicrobial Bacillus subtilis strain as a starter culture to improve qualities and safety of fermented soybean (sieng) produced in Cambodia. J. Food Eng. 2019, 9, 333–353. [Google Scholar] [CrossRef]
- Xie, Z.; Li, M.; Wang, D.; Wang, F.; Shen, H.; Sun, G.; Feng, C.; Wang, X.; Chen, D.; Sun, X. Biocontrol efficacy of Bacillus siamensis LZ88 against brown spot disease of tobacco caused by Alternaria alternate. Biol. Contr. 2021, 154, 104508. [Google Scholar] [CrossRef]
- Jeong, D.W.; Kim, H.R.; Jung, G.; Han, S.; Kim, C.T.; Lee, J.H. Bacterial community migration in the ripening of Doenjang, a traditional Korean fermented soybean food. J. Microbiol. Biotechnol. 2014, 24, 648–660. [Google Scholar] [CrossRef]
- Liu, Z.; Guan, X.; Zhong, X.; Zhou, X.; Yang, F. Bacillus velezensis DP-2 isolated from Douchi and its application in soybean meal fermentation. J. Sci. Food Agric. 2021, 101, 1861–1868. [Google Scholar] [CrossRef]
- Chaurasia, L.K.; Tamang, B.; Tirwa, R.K.; Lepcha, P.L. Influence of biosurfactant producing Bacillus tequilensis LK5.4 isolate of kinema, a fermented soybean, on seed germination and growth of maize (Zea mays L.). Biotech 2020, 10, 297. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, H.; Gao, J.; Portieles, R.; Du, L.; Gao, X.; Borotto Nordelo, C.; Borrás-Hidalgo, O. Endophytic Bacillus altitudinis strain uses different novelty molecular pathways to enhance plant growth. Front. Microbiol. 2021, 25, 692313. [Google Scholar] [CrossRef]
- Hwang, C.E.; Cho, K.M.; Joo, O.S. Diversity of Bacillus groups isolated from fermented soybean foods (‘Doenjang’and ‘Kanjang’) and their fermentation characteristics of ‘Cheonggukjang’. Korean J. Food Preser. 2020, 27, 946–958. [Google Scholar] [CrossRef]
- Lateef, A.; Adelere, I.A.; Gueguim-Kana, E.B. The biology and potential biotechnological applications of Bacillus safensis. Biologia 2015, 70, 411–419. [Google Scholar] [CrossRef]
- Wang, D.; Wang, H.; Zhan, Y.; Xu, Y.; Deng, J.; Chen, J.; Cai, D.; Wang, Q.; Sheng, F.; Chen, S. Engineering expression cassette of pgdS for efficient production of poly-γ-glutamic acids with specific molecular weights in Bacillus licheniformis. Front. Bioengineer. Biotechnol. 2020, 8, 728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, S.; Jiang, G.; Gan, L.; Xu, Z.; Tian, Y. Optimization of fermentation conditions, purification and rheological properties of poly (γ-glutamic acid) produced by Bacillus subtilis 1006-3. Prep. Biochem. Biotechnol. 2021, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 6. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al.; EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards) Statement on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 15: Suitability of taxonomic units notified to EFSA until September 2021. EFSA J. 2021, 20, 7045. [Google Scholar] [CrossRef]
- Bourdichon, F.; Laulund, S.; Tenning, P. Inventory of microbial species with a rationale: A comparison of the IDF/EFFCA inventory of microbial food cultures with the EFSA Biohazard Panel qualified presumption of safety. FEMS Microbiol. Lett. 2019, 366, 1–6. [Google Scholar] [CrossRef]
- Wu, T.; Xu, J.; Liu, J.; Guo, W.H.; Li, X.B.; Xia, J.B.; Zhang, Y.M.; Wang, R.Q. Characterization and initial application of endophytic Bacillus safensis strain ZY16 for improving phytoremediation of oil-contaminated saline soils. Front. Microbiol. 2019, 10, 991. [Google Scholar] [CrossRef]
- Rong, S.; Xu, H.; Li, L.; Chen, R.; Gao, X.; Xu, Z. Antifungal activity of endophytic Bacillus safensis B21 and its potential application as a biopesticide to control rice blast. Pestic. Biochem. Physiol. 2020, 162, 69–77. [Google Scholar] [CrossRef]
- Wang, D.; Hwang, J.S.; Kim, D.H.; Lee, S.; Kim, D.H.; Joe, M.H. A newly isolated Bacillus siamensis SB1001 for mass production of poly-γ-glutamic acid. Process Biochem. 2020, 92, 164–173. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, M.; Li, H.; Lu, D.; Ouyang, P. Efficient production of poly (γ-glutamic acid) by newly isolated Bacillus subtilis NX-2. Process Biochem. 2005, 40, 519–523. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Strains Number) | Products (%) | ||||
|---|---|---|---|---|---|
| Grep chhurpi | Peha | Peron namsing | Peruñyaan | Mean | |
| Bacillus subtilis (PH1-32, PH10-28, PH12-1, PH3-21, PH6-29, PH7-11, PH7-9, PH8-3, PN1-14, PN14-8, PN2-5, PN3-22, PN3-34, PN4-7, PN5-11, PN5-19, PN8-34, PN9-10, PN9-28, PY4-30 and PY6-3) | 0 | 53.33 | 64.71 | 18.18 | 34.05 |
| Bacillus siamensis (GC3-6, GC4-22, GC4-36, PY3-2, PY6-11 and PY6-5) | 42.86 | 0.00 | 0.00 | 27.27 | 17.53 |
| Bacillus velezensis (GC1-42, GC3-13, PH4 -3, PY1-3, PY2-5 and PY5-4) | 28.57 | 6.67 | 0.00 | 27.27 | 15.63 |
| Bacillus licheniformis (GC3-34, PH12-8, PH2-30, PH8-18, PH8-30, PH9-8 and PY4-18) | 14.29 | 33.33 | 0.00 | 9.09 | 14.18 |
| Bacillus tequilensis (PN12-26, PN2-3, PN4-29, PN4-32, PN5-18, PN9-22 and PY1-10) | 0.00 | 0.00 | 35.29 | 9.09 | 11.1 |
| Bacillus altitudinis (GC1-28) | 14.28 | 0.00 | 0.00 | 0 | 3.57 |
| Bacillus safensis subsp. Safensis (PY1-19) | 0.00 | 0.00 | 0.00 | 9.10 | 2.27 |
| Bacillus paralicheniformis (PH6-9) | 0 | 6.67 | 0.00 | 0 | 1.67 |
| Diversity indices | |||||
| Taxa_S | 4.0 | 4.0 | 2.0 | 6.0 | |
| Individuals | 7.0 | 15.0 | 17.0 | 11.0 | |
| Simpson_1-D | 0.693 | 0.595 | 0.456 | 0.793 | |
| Shannon_H | 1.277 | 1.063 | 0.649 | 1.673 | |
| Samples | Chi-Square, χ² (p-value) | ||||
| Peron namsing vs Peha | 0.009 * | ||||
| Peron namsing vs Grep churpii | 0.0002 * | ||||
| Peron namsing vs Peruyann | 0.003 * | ||||
| Peha vs Grep churpii | 0.01 * | ||||
| Peha vs Peruyann | 0.04 * | ||||
| Grep churpii vs Peruyann | 0.6 NS | ||||
| Amino acid (pmol) | B. velezensis GC1-42 | B. siamensis GC4-36 | B. subtilis PH3-21 | B. subtilis PN1-14 | B. tequilensis PN9-22 | B. safensis subsp. safensis PY1-19 | B. subtilis PH6-29 | Standard PGA ** |
|---|---|---|---|---|---|---|---|---|
| Aspartic acid | 69.1 | 142.3 | 129.3 | 104.4 | 155.8 | 55.5 | 57.7 | ND * |
| Threonine | 80.1 | 49.0 | 52.8 | 41.8 | 49.6 | 18.7 | 13.7 | ND |
| Serine | 147.0 | 94.6 | 118.3 | 96.0 | 110.8 | 37.7 | 37.7 | ND |
| Glutamic acid | 2298.8 | 3941.8 | 4766.5 | 2608.8 | 3338.1 | 1935.8 | 1100.2 | 69.6 |
| Proline | ND | ND | ND | ND | ND | ND | ND | ND |
| Glycine | 151.3 | 364.2 | 285.9 | 212.6 | 250.5 | 89.0 | 52 | ND |
| Alanine | 189.3 | 398.1 | 469.8 | 266.7 | 493.6 | 137.3 | 492 | ND |
| Cysteine | ND | ND | ND | ND | ND | ND | ND | ND |
| Valine | ND | ND | ND | ND | ND | ND | ND | ND |
| Methionine | ND | ND | ND | ND | ND | ND | ND | ND |
| Isoleucine | ND | ND | ND | ND | ND | ND | ND | ND |
| Leucine | ND | ND | ND | ND | ND | ND | ND | ND |
| Tyrosine | 229.0 | 181.0 | 204.4 | 67.9 | 273.8 | 66.8 | 77.8 | ND |
| Phenylalanine | 271.9 | 230.8 | 295.2 | 82.4 | 482.4 | 46.5 | 55.5 | ND |
| Histidine | 85.7 | 55.0 | 50.8 | 33.3 | 67.5 | 18.3 | 67.2 | ND |
| Lysine | 232.2 | 89.5 | 123.6 | 45.7 | 95.8 | 39.0 | 95.2 | ND |
| Arginine | 65.5 | 36.9 | 45.5 | 48.3 | 14.1 | 9.3 | 15.1 | ND |
| Total | 3819.9 | 5583.2 | 6542.1 | 3607.8 | 5332.0 | 2453.8 | 2064 | 69.6 |
| Glutamic acid (%) | 60.2 | 70.6 | 72.9 | 72.3 | 62.6 | 78.9 | 53.3 | 100.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pariyar, P.; Yaduvanshi, P.S.; Raghu, P.; Tamang, J.P. Screening of Poly-Glutamic Acid (PGA)-Producing Bacillus Species from Indian Fermented Soybean Foods and Characterization of PGA. Fermentation 2022, 8, 495. https://doi.org/10.3390/fermentation8100495
Pariyar P, Yaduvanshi PS, Raghu P, Tamang JP. Screening of Poly-Glutamic Acid (PGA)-Producing Bacillus Species from Indian Fermented Soybean Foods and Characterization of PGA. Fermentation. 2022; 8(10):495. https://doi.org/10.3390/fermentation8100495
Chicago/Turabian StylePariyar, Priyambada, Puneeta Singh Yaduvanshi, Pullakhanadam Raghu, and Jyoti Prakash Tamang. 2022. "Screening of Poly-Glutamic Acid (PGA)-Producing Bacillus Species from Indian Fermented Soybean Foods and Characterization of PGA" Fermentation 8, no. 10: 495. https://doi.org/10.3390/fermentation8100495
APA StylePariyar, P., Yaduvanshi, P. S., Raghu, P., & Tamang, J. P. (2022). Screening of Poly-Glutamic Acid (PGA)-Producing Bacillus Species from Indian Fermented Soybean Foods and Characterization of PGA. Fermentation, 8(10), 495. https://doi.org/10.3390/fermentation8100495