Abstract
Deep ocean water (DOW) is rich in mineral nutrients, and our previous studies have confirmed that it promotes fungal growth and metabolite production. Cordycepin is a functional metabolite of Cordyceps with anti-inflammatory and antitumor properties. Although Cordyceps militaris can produce a large amount of cordycepin when DOW is used as the culture water, no study has explored the relevant regulatory mechanism through which DOW increases the adenosine and cordycepin contents. Therefore, in this study, we used 2D electrophoresis and proteomic analysis to investigate the effects of DOW with different mineral compositions on proteins in the fruiting bodies of C. militaris and to explore how DOW regulates cordycepin biosynthesis. The results indicated that the use of DOW with high magnesium and calcium contents can increase the production of cordycepin from 6.62 mg/g to 11.40 mg/g by C. militaris fruiting bodies. According to the 2D electrophoresis and proteomic analysis results of the fruiting body proteins of C. militaris cultured with DOW with different concentrations, the expression of 17 and 9 proteins was proportional to the cordycepin content and the concentration of DOW, respectively. Magnesium, sulfate, and other minerals of DOW stimulated the production of cordycepin by promoting the glycolytic pathway, producing more ATP and adenosine. It also increased the production of nucleic acids and adenosine by promoting the pentose phosphate pathway and methionine cycle, thereby promoting the growth and cordycepin production of fruiting bodies.
1. Introduction
Deep ocean water (DOW) is low-temperature, clean water and is rich in minerals and nutrients. According to previous studies, DOW has several health benefits. For example, it has hypoglycemic [1], hypolipidemic [2], and anti-fatty liver [3] properties. DOW also promotes biomass growth and increases the production of metabolites. The rich minerals of DOW were proven as the important factor for stimulating biomass production and secondary metabolites in fungi, such as ankaflavin and monascin in Monascus purpureus-fermented rice [4], and triterpinoids and polysaccharides of Antrodia cinnamomea [5,6]. M. purpureus-fermented product cultured with DOW performed higher hypolipidemic effects and anti-adipogenesis effects in rats fed a high-fat diet [4,7,8]. A. cinnamomeas-fermented product cultured with DOW performed higher protection against liver fibrosis in the thioacetamide-injected rat model. The mycelium of Cordyceps cicadae absorbed and chelated the DOW magnesium. The fungal-bound magnesium can perform higher absorption capacity in the brain in the Alzheimer’s disease rat model [9]. In the Cordyceps militaris mycelium study, the magnesium, sodium, calcium, iron, and nitrate present in DOW can promote the production of cordycepin in Cordyceps militaris mycelium [10]. Furthermore, treatment of mice with C. militaris mycelium cultured with DOW ameliorated the development of liver fibrosis [11]. However, the effect of DOW on the growth and metabolism of a fruiting body by C. militaris were still unclear. Therefore, in this study, we investigated the effects of C. militaris cultured with DOW on the protein synthesis in and metabolic changes of fruiting bodies.
C. militaris is used as a Chinese medicine in Asia and mainly contains adenosine, cordycepin, and polysaccharides, among other components. Cordycepin is a unique active substance of Cordyceps, and its production in C. militaris is higher than that in other Cordyceps species. The production of cordycepin by the fruiting bodies of C. militaris can be further increased through artificial cultivation. Several experimental results have confirmed that C. militaris can regulate the oxidative stress and inflammatory response of the liver because of its high cordycepin content. Generally, the cordycepin content is threefold higher in the fruiting bodies of C. militaris than in the mycelia [12]. Previous studies have indicated that cordycepin can inhibit the lipopolysaccharide-induced production of tumor necrosis factor-α [13], increase the concentration or activity of antioxidants in rats, and ameliorate the oxidative damage caused by reactive oxygen species [14].
According to previous studies exploring the biosynthetic pathway of cordycepin, adenosine is first metabolized to 3′-AMP and then to 2′-C-3′-dA and finally to cordycepin [15]. Another biosynthetic pathway involves the formation of 2′,3′-cAMP phosphatase after RNA degradation, which is then converted into 3′-AMP and finally into cordycepin [16]. However, these studies have investigated only the chemical aspect of the biosynthetic process from adenosine to cordycepin and have not addressed the overall biochemical pathways and protein regulation mechanisms in proteomics, which remain unclear.
Proteomics is mainly used in protein identification, structure analysis, and posttranslational modification structure prediction and is also used in the analysis and comparison of a large number of proteins with different expression patterns. Therefore, proteomics can be used to explore how DOW regulates the production of cordycepin and biomass by the fruiting bodies of C. militaris. In this study, we used 2D electrophoresis and proteomic analysis to explore how DOW regulates biomass and cordycepin production by the fruiting bodies of C. militaris.
2. Materials and Methods
2.1. Chemicals
Yeast mold (YM) agar and broth were purchased from Difco Laboratories (Detroit, MI, USA). Magnesium chloride was purchased from Showa Chemical Industry (Tokyo, Japan). Ethanol (95%) was purchased from the Taiwan Tobacco and Liquor Corp. (Taipei, Taiwan). All other chemicals were purchased from Sigma Chemical (St. Louis, MO, USA).
2.2. Preparation of Various Deep Ocean Water Concentrations
Various concentrations of desalinated DOW (D1, D2, D3, and D4), provided by the Eastern Taiwan Deep Sea Water Innovation and Research Center (Taitung, Taiwan), were pumped from a depth of 670 m in the Pacific Ocean near eastern Taiwan and processed through electrodeionization. According to our previous study, we defined DOW with 20.65 mg/L Mg2+ as 1× DOW. For this study, we prepared 5× DOW (103.25 mg/L Mg2+), 10× DOW (206.5 mg/L Mg2+), 15× DOW (309.75 mg/L Mg2+), and 30× DOW (619.5 mg/L Mg2+) by diluting each concentration of DOW (D1, D2, D3, or D4) with ultrapure water (UPW).
2.3. Microorganism and Seed Cultures
Briefly, C. militaris CM027, provided by Acro Biomedical Co (Fishers, IN, USA), was maintained on potato dextrose agar at 28 °C and then transferred to fresh medium for 10 days. Seed cultures were then prepared by transferring a loopful of the colony from the YM agar slant into a 500 mL Hinton flask containing 100 mL of medium (3 g/L yeast extract, 5 g/L malt extract, 10 g/L peptone, and 3 g/L dextrose). Finally, the cultures were incubated at 20 °C with rotation shaking at 100 rpm for 72 h, and inoculum sizes of 5% were transferred to a submerged or solid cultured substrate.
2.4. Solid Fermentation of C. militaris in Deep Ocean Water or Ultrapure Water
Solid fermentation was performed using a 500 mL glass bottle containing a rice substrate (50 g of rice, 35 mL of UPW, various concentrations of DOW, and a magnesium chloride solution). After sterilization (at 121 °C for 20 min), the seed culture medium was inoculated and incubated at 20 °C in a dark room for 7 days. Subsequently, the C. militaris culture was incubated at 20 °C and relative humidity of 80–90% in a room with light (light for 24 h). Next, the fruiting bodies were sampled on days 30 and 60. After the solid culture, the fruiting bodies were collected, dried in a freeze dryer, and weighed. Finally, dried fruiting body powder was used for proteomic analysis and the analysis of cordycepin and adenosine.
2.5. Determination of Cordycepin and Adenosine
The dried fruiting body powder (0.1 g) was extracted with 1 mL of methanol at 50 °C for 1 h. The extracts (10%, w/v) were then filtered using a filter with a pore size of 0.45 μm and analyzed through high-performance liquid chromatography (HPLC) (model L-2130; Hitachi, Tokyo, Japan) on a C18 column (25 cm × 4.6 mm i.d., 5 μm; Phenomenex LUNA) with gradient elution. HPLC was performed in triplicate according to a previously described method [10]. Cordycepin and adenosine were separated through gradient elution with a mobile phase consisting of water/methanol (95.0/5.0 to 58.4/41.6 in 20 min, v/v). The flow rate was set at 0.8 mL/min. Finally, cordycepin and adenosine were detected using a photodiode array detector (model L-2455 DAD; Hitachi) set at 260 nm and the full wavelength.
2.6. Protein Extraction
Fresh fruiting bodies were obtained by grinding wet biomass in liquid nitrogen. After cell disruption, the biomass was resuspended in 25 mL of methanol at −20 °C for 24 h. The mixture was then incubated for at least 1 h at 4 °C, and the precipitated proteins were collected by centrifugation at 22,000× g for 1 h at 4 °C. After centrifugation, the supernatant was discarded. Subsequently, methanol (25 mL) was added again to the pellet, and the supernatant was collected by centrifugation (16,000× g at 4 °C for 10 min). This process was repeated three times, and acetone (25 mL) was then added and reacted at −20 °C for 15 min. Next, after centrifugation (16,000× g at 4 °C for 10 min), the supernatant was discarded, and this process was repeated twice. The final protein precipitate was dried in a hood, and the protein precipitate was redissolved in a solubilization buffer (7 M urea, 2 M thiourea, 4% CHAPS, 2% DTT, 2% pharmalyte, and 1% SERDOLIT MB-1) with shaking for 2 h. After centrifugation (12,000× g at 4 °C for 15 min), the solubilized proteins were stored at −70 °C until further 2D gel electrophoresis analysis.
2.7. 2D Gel Electrophoresis
The first dimension with isoelectric focusing (IEF) was performed on an IPGphor Isoelectric Focusing System (Amersham Biosciences, Buckinghamshire, UK) at 20 °C with a current of 45 μA per strip. Next, 200 μL of each protein sample (2.5 mg/g) was loaded onto Immobiline DryStrip gel at a pH of 3–10 (IPG strips; Amersham Biosciences) by in-gel rehydration. IEF was then performed under the following conditions: 30 V × 12 h, 500 V × 3 h, 1000 V × 2 h, 2000 V × 2 h, 4000 V × 3 h, and 8000 V × 9 h. Before the second dimension (sodium dodecyl sulfate–polyacrylamide gel electrophoresis), the IPG strips were equilibrated for 15 min in 6 M urea, 30% glycerol, 3% sodium dodecyl sulfate, 2% bromophenol blue, and 50 mM Tris-HCl (pH 8.8) with 1% (w/v) DTT and then for 15 min in the same buffer with 2.5% (w/v) iodoacetamide. For the second dimension, the equilibrated strips were then transferred onto lab-cast sodium dodecyl sulfate–polyacrylamide gels, and the proteins were separated on a 12–16% linear gradient and 12% uniform gels. The second dimension was performed using a Hoefer vertical separation unit (Amersham Biosciences) at 10 °C in a constant working voltage mode as follows: 80 V for 1 h and then 125 V overnight until the bromophenol blue dye front reached the bottom of the gel.
The gel was then stained with colloidal Coomassie blue G-250 and scanned at 300 dpi (Epson Perfection V800 Photo; Epson, Jakarta, Indonesia). Image analysis was then performed using PDQuest 2-D analysis software (Bio-Rad, Hercules, CA, USA). The MASCOT 2.1.0 search program (Matrix Science, London, UK) was applied for protein identification by using the annotated C. militaris genome (http://www.ebi.ac.uk/genomes/eukaryota.html, access on 13 August 2022) as the database. All annotations were based on the sequenced genome of C. militaris and the National Center for Biotechnology Information (NCBI) reference sequence database.
2.8. Statistical Analysis
Data are expressed as mean ± standard deviation (SD). Analysis of variance with Duncan’s test and Pearson’s product-moment correlation coefficient test was performed using SPSS version 10.0 (SPSS, Chicago, IL, USA). Differences with p < 0.05 were considered statistically significant.
3. Results
3.1. Mineral Composition of Deep Ocean Water
The mineral compositions of the four concentrations of desalinated DOW (D1, D2, D3, and D4) are presented in Table 1. All four DOW solutions had the same magnesium content but different contents of other minerals. D1 and D2 had higher contents of calcium, potassium, sodium, and other minerals than D3 and D4, but their sulfate content was lower. In addition, D1 had a higher content of copper than D2, and D4 had the lowest chloride content among the four concentrations of DOW. Therefore, we adjusted the magnesium content in the DOW solutions to be the same. We then investigated the effects of DOW minerals other than magnesium on the growth of C. militaris fruiting bodies and the metabolism of cordycepin, and we further explored the main potential mechanisms by using proteomics.

Table 1.
Mineral concentrations in various DOW solutions.
3.2. Effect of Deep Ocean Water on the Biomass of C. militaris Fruiting Bodies and the Content of Functional Components
C. militaris fruiting bodies were obtained by culturing C. militaris with the different DOW solutions or a magnesium chloride solution. For 1× DOW, we used a magnesium concentration of 20.65 mg/L in each group. The fruiting body dry weight results for each group are presented in Figure 1. On day 40 of fermentation, the fruiting-body biomass significantly increased with the DOW concentration as follows: D1 (15× and 30×, p < 0.01), D2 (30×, p < 0.05), D3 (5× and 30×, p < 0.01), and D4 (5× and 15×, p < 0.05)—along with a significant increase in biomass with the magnesium chloride content (5×, p < 0.01; 15×, p < 0.05). From day 40 to day 60, the fruiting-body biomass significantly increased in all groups. On day 60 of fermentation, the fruiting-body biomass significantly increased with the DOW concentration as follows: D2 (15× and 30×, p < 0.05; 5×, p < 0.01), D3 (30×, p < 0.05; 5× and 15×, p < 0.01), and D4 (5×, 15×, and 30×, p < 0.05)—along with a significant increase in biomass with the magnesium chloride content (5× and 15×, p < 0.01; 30×, p < 0.05). The fruiting-body biomass exhibited the most significant increase with D2 (5×) and magnesium chloride (5× and 15×) compared with UPW (p < 0.01). These results indicated that magnesium chloride is the main component in DOW causing the increase in the biomass of fruiting bodies. However, although the four DOW had an equal content of magnesium, they also showed different amounts of biomass. This suggested that the biomass may also be regulated by other minerals in DOW.
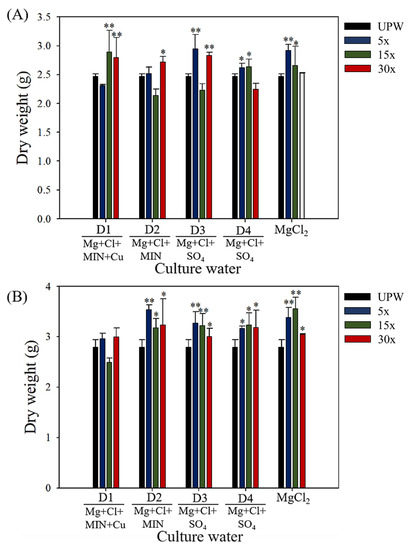
Figure 1.
Biomass dry weight of C. militaris fruiting bodies cultured with different concentrations and mineral compositions of DOW, magnesium chloride solution, and UPW: (A) day 40, (B) day 60. Data are presented as mean ± SD (n = 3). * and ** indicate significant differences (p < 0.05, p < 0.01, and p < 0.001, respectively) as compared with UPW.
The adenosine content of the fruiting bodies of each group is presented in Figure 2. On culture day 40, the adenosine content significantly increased with the DOW concentration as follows: D1 (5× and 15×, p < 0.01; 30×, p < 0.01), D2 (5× and 15×, p < 0.01), D3 (5×, p < 0.001; 15×, p < 0.05; 30×, p < 0.01), and D4 (5×, 15×, and 30×, p < 0.001)—along with a significant increase in the adenosine content with the magnesium chloride content (5×, p < 0.05; 15×, p < 0.01; and 30×, p < 0.001). However, on day 60, the adenosine content did not exceed that observed on day 40. On culture day 60, the adenosine content remained significantly high in the D1 (15× and 30×, p < 0.05), D3 (5×, p < 0.05; 15× and 30×, p < 0.001), and D4 (5×, 15×, and 30×, p < 0.01) groups, along with a significant increase in the adenosine content with the magnesium chloride content (5×, 15×, and 30×, p < 0.01). These results indicated that the magnesium chloride content in DOW is one of the critical factors increasing the adenosine content in fruiting bodies, and the mineral components of D3 had a greater effect on increasing the adenosine content than UPW.
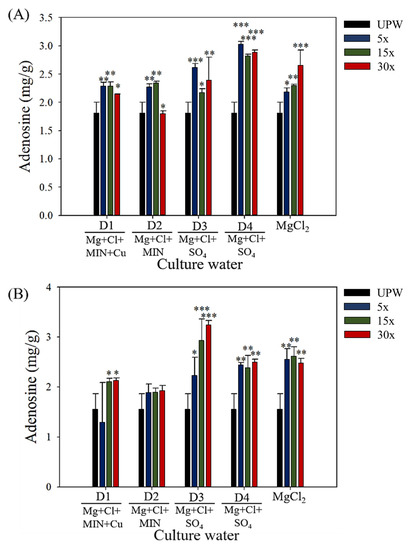
Figure 2.
Production of adenosine in C. militaris fruiting bodies cultured with different concentrations and mineral compositions of DOW, magnesium chloride solution, and UPW: (A) day 40, (B) day 60. Data are presented as mean ± SD (n = 3). *, **, and *** indicate significant differences (p < 0.05, p < 0.01, and p < 0.001, respectively) as compared with UPW.
The cordycepin content in the fruiting bodies of each group is presented in Figure 3. When cultured for 40 days, C. militaris fruiting bodies produced less than 2.5 mg/g of cordycepin. However, after 60 days of culture, cordycepin production reached 6–12 mg/g. This means that the conversion of cordycepin mainly occurs within 40–60 days. To obtain high cordycepin production, a cultivation period of up to 60 days was required. After culture day 60, cordycepin production significantly increased with the DOW concentration: D3 (5×, p < 0.05; 15×, p < 0.01) and D4 (30×, p < 0.05). However, cordycepin production did not increase with D1, D2, or magnesium chloride. This means that magnesium chloride is not the main nutrient salt promoting the bioconversion and production of cordycepin. However, the mineral content and composition of D3 played a potential role in the cordycepin production.
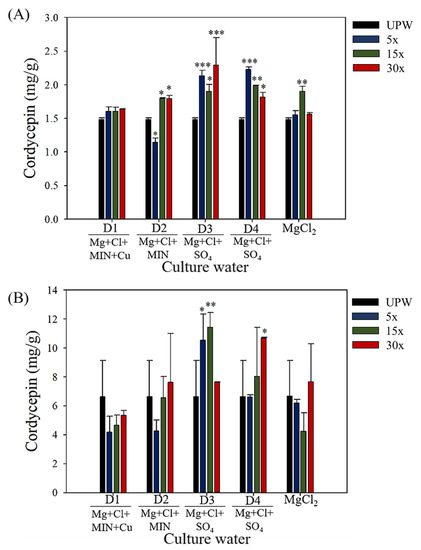
Figure 3.
Production of cordycepin in C. militaris fruiting bodies cultured with different concentrations and mineral compositions of DOW, magnesium chloride, and UPW: (A) day 40, (B) day 60. Data are presented as mean ± SD (n = 3). *, **, and *** indicate significant differences (p < 0.05, p < 0.01, and p < 0.001, respectively) as compared with UPW.
3.3. Effects of Deep Ocean Water with Different Mineral Compositions on the Protein Expression of C. militaris Fruiting Bodies
We used 2D electrophoresis and proteomic analysis to investigate the effect of various DOW and magnesium chloride concentrations on the expression of the fruiting body proteins of C. militaris. After separation through 2D electrophoresis, protein extraction from fruiting bodies cultured with DOW with different mineral compositions and concentrations yielded 800–1200 protein spots. We therefore investigated the effect of the 15× concentration of D1, D2, D3, D4, and magnesium chloride solutions on C. militaris proteins. The 2D electrophoresis patterns of all groups are presented in Figure 4. We further analyzed the expression of protein spots and selected those with significant differences in protein expression among the DOW groups as the target protein spots, obtaining a total of 20 target protein spots. Among all groups, the fruiting bodies of D3 (15×) had the highest cordycepin content. Therefore, we further analyzed protein spots with the highest expression of D3. The 2D electropherogram results of the D3 group indicated that protein spots 1001, 1303, 1304, 3301, 3403, 3503, 3506, 4403, 4409, 4604, 4705, 4707, 4801, 4802, 5301, 5404, 6503, 6706, 7504, and 8701 exhibited higher expression levels than those of UPW and other DOW groups.
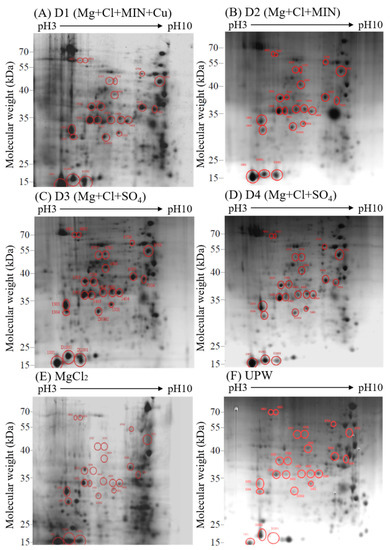
Figure 4.
2D electrophoresis of the fruiting body proteins of C. militaris cultured with (A–E) magnesium chloride solution, and (F).
Liquid chromatography with tandem mass spectrometry (LC-MS/MS) was performed for protein spot expression analysis and peptide identification. Differences in protein spot expression across the samples are presented in Figure 5. As shown in the figure, D3 (15×) not only increased the production of cordycepin but also resulted in high protein expression levels in various protein spots. These protein spots were identified as Woronin body major protein (WB), glyceraldehyde-3-phosphate dehydrogenase (G3PD), serine protease, formate dehydrogenase, 6-phosphogluconate dehydrogenase (6PGD, decarboxylating), heat shock 70 kDa protein (Hsp70), β-hexosaminidase, transaldolase, cyanide hydratase, enolase, carbohydrate-binding WSC, and heat shock 90 kDa protein (Hsp90). D2 (15×) also promoted the expression levels of various protein spots, including Hsp70, glycoside superfamily, T-complex 1 subunit alpha, and 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase.
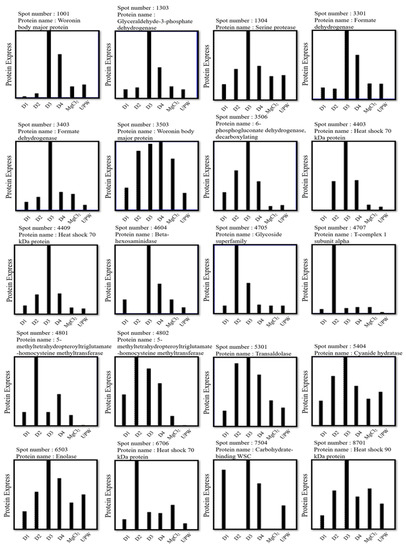
Figure 5.
2D electrophoresis results of the effect of various DOW solutions (D1, D2, D3, and D4) on the protein expression levels of C. militaris fruiting bodies.
3.4. Effects of Different Concentrations of Deep Ocean Water on the Expression of C. militaris Fruiting Body Proteasome
We compared the effect of the DOW concentration on protein performance. The results obtained indicated that the D4 dose dependently increased the production of cordycepin. Therefore, we analyzed and compared the protein expression levels of fruiting bodies cultured with various concentrations of D4 (0×, 5×, 15×, and 30×) and explored the relationship between DOW-regulated protein expression levels and cordycepin biosynthesis. As shown in Figure 6, the 2D electrophoresis results were different because of the increased concentration of D4. Hence, using a 2D electropherogram, we analyzed the expression of protein spots (Figure 7). We also screened all protein spots whose expression increased with the concentration of DOW and cordycepin. A total of 11 target protein spots were obtained: 1001, 1303, 1304, 3301, 3503, 5301, 5404, 6503, D1001, D2001, and D3302. These protein spots were identified by LC-MS/MS as WB, G3PD, serine protease, formate dehydrogenase, transaldolase, cyanide hydratase, enolase, superoxide dismutase, and fructose-bisphosphate aldolase (FBPA).
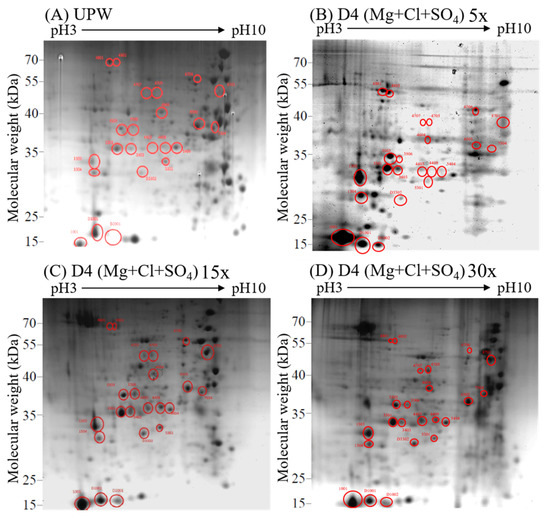
Figure 6.
2D electrophoresis of C. militaris fruiting body proteins cultured with different concentrations of D4: (A–D) 30× D4.
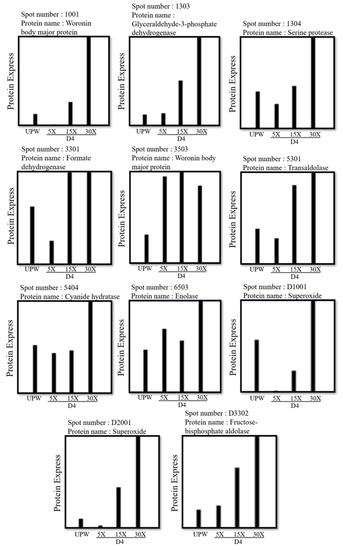
Figure 7.
2D electrophoresis results of the effect of various concentrations of D4 on the protein expression levels of C. militaris fruiting bodies.
A total of 23 protein spots were identified, and their protein identities are presented in Table 2. These proteins were divided into three categories: (1) DOW-regulated carbohydrate and energy metabolism proteins; (2) DOW-regulated and protein synthesis, folding, and protective functional proteins; and (3) other functional proteins.
Table 2.
Identities of the C. militaris fruiting body proteins identified in 2D electrophoresis.
As shown in Table 3, the carbohydrate and energy metabolism proteins regulated by DOW included G3PD, 6PGD (decarboxylating), β-hexosaminidase, glycoside superfamily, transaldolase, enolase, and FBPA. On the other hand, the protein synthesis, folding, and protective functional proteins regulated by DOW included serine protease, Hsp70, T-complex 1 subunit alpha, 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase, and Hsp90. Other functional proteins included WB, formate dehydrogenase, cyanide hydratase, carbohydrate-binding WSC, and superoxide dismutase.
Table 3.
Fruiting body proteins that are regulated by DOW and that demonstrated a substantial change in protein quality and cordycepin content.
4. Discussions
During the growth cycle of C. militaris fruiting bodies, mycelia are first formed in the cereal medium. During this mycelial growth phase, adenosine is produced. With continuous culturing, fruiting bodies form after day 30. During the first 30 days of culture, the amount of cordycepin produced is small. However, its production starts to increase from day 30 to day 60 of culture. This means that the mycelial growth phase and adenosine production phase occur simultaneously, and that the conversion of adenosine into cordycepin occurs only after the complete formation of fruiting bodies. In this study, we investigated how the mineral composition of DOW can enhance cordycepin biosynthesis by fruiting bodies and the adenosine and cordycepin contents of C. militaris. Our findings indicated that the composition and concentration of DOW minerals should be important factors for affecting the biosynthesis of cordycepin. Therefore, we investigated the relationship between cordycepin biosynthesis and the composition of DOW minerals by using proteomics.
DOW contains a large amount of magnesium chloride. The four DOW solutions with different mineral compositions used in this study not only had higher contents of sodium, potassium, strontium, barium, and sulfate than that of magnesium chloride but also contained several trace elements. Therefore, we used the same magnesium content across all groups and investigated the effects of different mineral compositions on the growth of C. militaris fruiting bodies and the production of cordycepin. The results indicated that the mineral compositions of D1 and D2 were similar. Compared with D3 and D4, both D1 and D2 had higher contents of sodium, potassium, calcium, strontium, and barium and a lower content of sulfate. However, D1 had a higher copper content than D2, and D3 had a higher sulfate content than D4, with the chloride content of D4 being the lowest among all four DOW solutions.
According to our results, magnesium chloride can promote the formation of fruiting bodies and adenosine production, but it does not affect the production of cordycepin. This means that magnesium ions are mainly associated with the growth of fungi and nucleotide biosynthesis but do not affect the conversion process of cordycepin, although chloride ions may inhibit the production of cordycepin. According to previous studies, high concentrations of chloride ions can reduce the biosynthesis of cordycepin, and the cordycepin content can be reduced by culturing C. militaris with a chloride salt solution [10]. These results indicate that magnesium can considerably increase the biomass of fruiting bodies, promote the biosynthesis of adenosine, and offset the inhibitory effect caused by chloride ions.
The results obtained indicated that the biomass and adenosine content of the fruiting bodies of C. militaris cultured with D1 and D2 were higher than those of C. militaris cultured with RO water. Both D3 and D4 considerably increased the cordycepin content, suggesting that the cordycepin content is related to the composition of DOW minerals. This is why we sought to clarify the relationship between mineral composition and cordycepin production. From the results obtained, we concluded that magnesium ions can increase biomass and the biosynthesis of adenosine but do not considerably affect the production of cordycepin.
The sulfate content of both D3 and D4 was more than twice that of D1 and D2, but D3 and D4 had lower contents of sodium, potassium, calcium, strontium, iron, copper, and barium than D1 and D2. This finding means that sulfate may be regarded as an important salt in DOW that enhances the production of cordycepin in fruiting bodies. The biomass and adenosine production in fruiting bodies cultured with D3 and D4 were not considerably higher than those with D1, D2, and magnesium chloride solutions, indicating that sulfate may not directly affect the formation of fruiting bodies and the production of adenosine. D3 demonstrated its highest capability to increase the cordycepin content at 5× and 15×, whereas D4 demonstrated its highest capability at 30×. In addition, the concentration of D4 was proportional to the cordycepin content, and both D3 and D4 had a high sulfate content. According to previous studies on submerged cultures, magnesium, sodium, calcium, iron, and NO3− promote the production of cordycepin by C. militaris mycelia treated with DOW, whereas sulfate promotes the biosynthesis of adenosine [10]. Previous studies have also indicated that adenosine is a precursor in the production of cordycepin [28]. This means that sulfate may promote the biosynthesis of adenosine and its conversion into cordycepin.
According to previous studies, in the biosynthesis of cordycepin, adenosine is first metabolized into 3′-AMP and then into 2′-C-3′-dA and finally converted into cordycepin [15]. Another pathway of cordycepin biosynthesis involves the formation of 2′,3′-cAMP after RNA degradation, which is then converted into 3′-AMP by a phosphatase enzyme and finally into cordycepin [16]. Various catalytic enzymes are involved in these two pathways, such as nucleotide kinase, adenylate cyclase (AC), phosphodiesterase (PDE), phosphoesterase, and phosphatase. Several studies have also indicated that strontium and calcium regulate adenylate cyclase to limit the production of cAMP [29,30] while regulating the calcium-dependent phosphodiesterase [31]. However, copper prevents the binding and dissociation of GTP and gas, inhibiting the activation of AC by GTP and reducing the production of cAMP. Copper also inhibits the activity of phosphodiesterase [32]. In addition to strontium and calcium, D1 and D2 contained several mineral elements. D1 also had a high copper content, which may reduce the expression of cAMP and inhibit the increase in the cordycepin production. Both D3 and D4 had low contents of copper, strontium, and calcium, but this did not affect the production of cordycepin. Therefore, we used 2D electrophoresis to explore the differences in the proteomic performance of C. militaris fruiting bodies cultured in DOW with different mineral compositions.
The results obtained in the present study indicated that adding magnesium chloride at 5–15× substantially promoted the formation of C. militaris fruiting bodies. The results also indicated that magnesium chloride content more than 5× substantially increased the production of adenosine. However, magnesium chloride did not have a considerable effect on the production of cordycepin. According to the protein expression results, 15× magnesium chloride enhanced the expression of WB, Hsp70, formate dehydrogenase, β-hexosaminidase, and 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase. This finding means that magnesium chloride may increase the formation of fruiting bodies by increasing the expression of these proteins. Although D1 did not affect the formation of fruiting bodies, it increased the production of adenosine when added at 10× and 15×. However, this effect was not reflected in the final generation of cordycepin and exhibited a downward trend. Compared with the magnesium chloride group, the D1 group demonstrated lower expression levels of WB, enolase, Hsp70, and Hsp90. In addition, although the D1 and magnesium chloride groups had the same concentration of magnesium, D1 eventually inhibited the formation of fruiting bodies, presumably because some mineral elements may have inhibited the expression of these proteins.
Although the compositions of D2 and D1 were similar, D2 considerably increased the formation of fruiting bodies, whereas D1 did not. In addition, unlike D1, D2 did not considerably promote the biosynthesis of adenosine. However, none of these DOW solutions increased the production of cordycepin. D2 demonstrated considerably higher expression levels of glycoside superfamily, T-complex 1 subunit alpha, and 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase. The glycoside superfamily is a family of proteases that hydrolyze glycosidic bonds. A glycosidic bond is the bond between the base and a ribose or deoxyribose in a nucleoside or the bond between the sugar groups. Bacteria with increased expression of this enzyme can more effectively degrade sugars, nucleosides, and other substances and can convert them into raw materials for biomass growth and energy metabolism.
The glycoside superfamily includes fungal chitinases, which are involved in the breakdown and release of polysaccharides. T-complex 1 subunit alpha is a major element of the chaperonin-containing T-complex (TRiC), a molecular chaperone complex that assists in the folding of proteins upon ATP hydrolysis. This TRiC plays a role in the folding of actin and tubulin [19], which are speculated to be components of the somatic cytoskeletal proteins of C. militaris. Finally, 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase is involved in protein modification and methyl transfer. These proteins play a crucial role in protein modification and polysaccharide formation and are released during growth.
Among all groups, only D3 at 15× considerably increased the formation of fruiting bodies and the production of adenosine and cordycepin. D3 also enhanced the expression of WB, formate dehydrogenase, G3PD, serine protease, Hsp70, enolase, β-hexosaminidase, cyanide hydratase, enolase, and carbohydrate-binding WSC. These results indicate that D3 can increase the fruiting-body biomass and the production of adenosine and cordycepin by increasing the expression of these proteins.
Although D4 did not substantially increase the production of cordycepin, the amount produced gradually increased with the increase in the concentration of D4. Therefore, we used D4 to explore the relationship between the concentration of deep seawater and the expression of each protein. As shown in Table 1, Table 2 and Table 3, increasing the concentration of D4 from 5× to 15× increased the formation of fruiting bodies and the production of adenosine but did not considerably increase the production of cordycepin. However, when the concentration of D4 reached 30×, the growth of fruiting bodies and the production of adenosine and cordycepin considerably increased. As shown in Figure 7, D4 at 30× considerably increased the expression levels of WB, G3PD, serine protease, cyanide hydratase, enolase, superoxide dismutase, and FBPA. D4 also enhanced the growth of fruiting bodies and increased the production of adenosine and cordycepin by regulating the expression of these proteins. For instance, D4 at 15× increased the expression levels of WB, formate dehydrogenase, and transaldolase, which is a similar trend to that observed for the production of mycelia and adenosine. These results indicate that these three proteins are key proteins underlying the enhanced production of mycelia and adenosine by D4.
According to the aforementioned results, DOW can promote the production of adenosine and the growth of fruiting bodies. Thus, the trend observed in the formation of fruiting bodies is similar to that observed in the production of adenosine. These results indicate that magnesium is the main mineral that, along with other mineral elements, enhances the expression of the key proteins of Cordyceps and promotes the formation of fruiting bodies and synthesis of adenosine. These proteins include (1) mycelial structure stabilization and protection proteins (e.g., WB, Hsp70, Hsp90, and β-hexosaminidase), (2) amino acid and nucleotide biosynthesis–related proteins (e.g., 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase), and (3) detoxification enzymes (e.g., formate dehydrogenase). Carbohydrate-binding WSC is a protein that is required to locate WB in cells [26]. WB is a specialized, fungus-specific organelle that enables the closure of septal pores after injury to protect the hyphae against excessive cytoplasmic bleeding [23], facilitating the rest of the damaged cells to continue to grow. Increased WB expression may represent robust cell division and hyphal extension of C. militaris fruiting bodies, resulting in increased septa. The inferences described in this section echo the boost of TCP1, indicating that DOW may contribute to the stability of mycelial proteins. In case of mycelial damage, both Hsp90 and Hsp70 assist in the repair process. Hsp70 is involved in protein folding, synthesis, and assembly and protects proteins against oxidation, hydrophobicity, and high-temperature denaturation. Hsp70 is also involved in the inhibition of apoptosis [21]. Hsp90 is a critical component of the Hsp90–Hsp70 chaperone protein. After its translation in the ribosome, Hsp70 causes initial folding, and then Hsp90 is used in refolding or structure maintenance. Overall, Hsp70 is an extremely valuable protein in maintaining the growth of fungal cells [33].
Cyanide is a common toxic substance for living organisms; its degradation is a critical part of metabolism. In fungi, cyanide is metabolized through a two-step hydrolytic mechanism. Cyanide is first converted by cyanide hydratase into formamide, which in turn is converted into formate [25]. Next, formate is further degraded into CO2 and NADH by formate dehydrogenase. Our results indicated that the expression levels of cyanide hydratase and formate dehydrogenase in the fruiting bodies of C. militaris cultured in DOW were much higher than in UPW, which may indicate that these two enzymes can effectively degrade cyanide, reduce cyanide residues, and protect cells. N-acetylglucosamine is a common hexosamine that exists in the chitin of fungal cell walls, glycoproteins of animal and plant cells, mucopolysaccharides, intercellular substances, and tissue-supporting substances. β-Hexosaminidase is an enzyme that hydrolyzes the N-terminal acetyl group of hexosamines and is, therefore, a critical fungal component that plays a role in the degradation of carbohydrates and complex glycans and in the recovery of cellular metabolites [18]. Increased β-hexosaminidase expression in cells may indicate that these cells can break down the matrix more rapidly to obtain nutrients. L-methionine is a sulfur-containing amino acid, and various organisms synthesize S-adenosyl-L-methionine (SAM) to promote the methionine cycle [22]. L-methionine serves as a source of adenosine and methyl groups. To synthesize SAM, 5-methyltetrahydropteroyltriglutamate–homocysteine S-methyltransferase converts L-homocysteine into L-methionine. When SAM is converted into S-adenosyl-L-homocysteine (SAH), methyl groups are released for the synthesis of DNA, RNA, proteins, and phospholipids [22]. SAH is also metabolized into L-homocysteine or adenosine, which can be used as a raw material source for cordycepin.
Generally, DOW can promote the production of cordycepin because of its magnesium content (which increases biomass and adenosine production) and because it contains sufficient amounts of sulfate and other mineral nutrients and low contents of strontium and barium. Magnesium, sulfate, and other minerals promote the production of cordycepin, presumably by enhancing sugar metabolism and nucleotide production for proteins (e.g., G3PD, enolase, FBPA, and 6PGD), and the expression of protein-modification enzymes (e.g., serine protease) and antioxidant and detoxification enzymes (e.g., superoxide dismutase, cyanide hydratase). FBPA, G3PD, and enolase are involved in glycolysis, promote energy metabolism, and play a critical role in the formation of fruiting bodies in fungi [17]. The FBPA and G3PD contents are more than eightfold higher than in other groups. It is speculated that while promoting the conversion of FBP to G3P and DHAP, the faster metabolism of G3P to 1,3-BP and the reduction of G3P to DHAP, another critical enzyme in the pentose phosphorylation pathway (PPP) is 6PGD. In this pathway, glucose is used to generate ribulose 5-phosphate as a raw material for nucleoside biosynthesis. 6PGD participates in the final step of PPP, which is the oxidative decarboxylation of 6-phosphogluconic acid to the five-carbon ribulose 5-phosphate. Simultaneously, NADP+ is converted into NADPH, which serves as a source of coenzymes for the synthesis of lipids and nucleotides by cells. Therefore, promoting the synthesis of nucleoside raw materials can increase the adenosine content, thereby increasing the production of cordycepin.
Author Contributions
Conceptualization, C.-L.L.; methodology, C.-H.L. and H.-L.H.; validation, C.-L.L.; formal analysis, C.-H.L. and H.-L.H.; investigation, C.-H.L. and C.-L.L.; resources, Y.-H.C.; data curation, C.-H.L.; writing—original draft preparation, C.-H.L. and C.-L.L.; writing—review and editing, C.-L.L.; funding acquisition, Y.-H.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Economic Affairs of Taiwan, grant number MOST 109-2218-E-143-001.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data included in this study are available upon request by contacting the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hwang, H.S.; Kim, H.A.; Lee, S.H.; Yun, J.W. Anti-obesity and antidiabetic effects of deep sea water on ob/ob mice. Mar. Biotechnol. 2009, 11, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, S.; Yasukawa, T.; Nakagawa, K.; Miyake, M.; Yamasaki, M.; Katahira, K.; Mohri, M.; Shimizu, T.; Hazama, A. Deep-sea water improves cardiovascular hemodynamics in Kurosawa and Kusanagi-Hypercholesterolemic (KHC) rabbits. Biol. Pharm. Bull. 2008, 31, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Lee, C.L. Comparison of the Improvement Effect of Deep Ocean Water with Different Mineral Composition on the High Fat Diet-Induced Blood Lipid and Nonalcoholic Fatty Liver Disease in a Mouse Model. Nutrients 2021, 13, 1732. [Google Scholar] [CrossRef] [PubMed]
- Lung, T.Y.; Liao, L.Y.; Wang, J.J.; Wei, B.L.; Huang, P.Y.; Lee, C.L. Metals of Deep Ocean Water Increase the Anti-Adipogenesis Effect of Monascus-Fermented Product via Modulating the Monascin and Ankaflavin Production. Mar. Drugs 2016, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yang, S.; Lee, D.Y.; Lee, C. Increasing anti-Abeta-induced neurotoxicity ability of Antrodia camphorata-fermented product with deep ocean water supplementary. J. Sci. Food Agric. 2016, 96, 4690–4701. [Google Scholar] [CrossRef]
- Wang, L.C.; Kuo, I.U.; Tsai, T.Y.; Lee, C.L. Antrodia camphorata-fermented product cultured in deep ocean water has more liver protection against thioacetamide-induced fibrosis. Appl. Microbiol. Biotechnol. 2013, 97, 9955–9967. [Google Scholar] [CrossRef]
- Lee, C.L. The advantages of deep ocean water for the development of functional fermentation food. Appl. Microbiol. Biotechnol. 2015, 99, 2523–2531. [Google Scholar] [CrossRef]
- Wang, L.C.; Lung, T.Y.; Kung, Y.H.; Wang, J.J.; Tsai, T.Y.; Wei, B.L.; Pan, T.M.; Lee, C.L. Enhanced anti-obesity activities of red mold dioscorea when fermented using deep ocean water as the culture water. Mar. Drugs 2013, 11, 3902–3925. [Google Scholar] [CrossRef]
- Wu, Y.Z.; Lee, C.L. Cordyceps cicadae NTTU 868 Mycelium with The Addition of Bioavailable Forms of Magnesium from Deep Ocean Water Prevents the Aβ40 and Streptozotocin-induced Memory Deficit via Suppressing Alzheimer’s Disease Risk Factors and Increasing Magnesium Uptake of Brain. Fermentation 2021, 7, 39. [Google Scholar]
- Hung, Y.P.; Wang, J.J.; Wei, B.L.; Lee, C.L. Effect of the salts of deep ocean water on the production of cordycepin and adenosine of Cordyceps militaris-fermented product. AMB Express 2015, 5, 140. [Google Scholar] [CrossRef]
- Hung, Y.P.; Lee, C.L. Higher Anti-Liver Fibrosis Effect of Cordyceps militaris-Fermented Product Cultured with Deep Ocean Water via Inhibiting Proinflammatory Factors and Fibrosis-Related Factors Expressions. Mar. Drugs 2017, 15, 168. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.H.; Lee, S.Y.; Sung, G.H.; Kim, S.H.; Choi, H.K. Metabolic profiles and free radical scavenging activity of Cordyceps bassiana fruiting bodies according to developmental stage. PLoS ONE 2013, 8, e73065. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Xu, Y.; Shen, J. Cordycepin inhibits lipopolysaccharide (LPS)-induced tumor necrosis factor (TNF)-alpha production via activating amp-activated protein kinase (AMPK) signaling. Int. J. Mol. Sci. 2014, 15, 12119–12134. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, T.; Yoo, S.K.; Kim, S.W.; Hwang, S.Y.; Sohn, S.H.; Kim, I.W.; Kim, S.K. Cordycepin (3’-deoxyadenosine) attenuates age-related oxidative stress and ameliorates antioxidant capacity in rats. Exp. Gerontol 2012, 47, 979–987. [Google Scholar] [CrossRef]
- Raethong, N.; Laoteng, K.; Vongsangnak, W. Uncovering global metabolic response to cordycepin production in Cordyceps militaris through transcriptome and genome-scale network-driven analysis. Sci. Rep. 2018, 8, 9250. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489.e4. [Google Scholar] [CrossRef]
- Feng, K.; Wang, L.Y.; Liao, D.J.; Lu, X.P.; Hu, D.J.; Liang, X.; Zhao, J.; Mo, Z.Y.; Li, S.P. Potential molecular mechanisms for fruiting body formation of Cordyceps illustrated in the case of Cordyceps sinensis. Mycology 2017, 8, 231–258. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, F.; Wang, T.; Lyu, Y.M.; Alteen, M.G.; Cai, Z.P.; Cui, Z.L.; Liu, L.; Voglmeir, J. Characterization of Stackebrandtia nassauensis GH 20 Beta-Hexosaminidase, a Versatile Biocatalyst for Chitobiose Degradation. Int. J. Mol. Sci. 2019, 20, 1243. [Google Scholar] [CrossRef]
- Seixas, C.; Cruto, T.; Tavares, A.; Gaertig, J.; Soares, H. CCTalpha and CCTdelta chaperonin subunits are essential and required for cilia assembly and maintenance in Tetrahymena. PLoS ONE 2010, 5, e10704. [Google Scholar] [CrossRef]
- Bertels, L.K.; Fernandez Murillo, L.; Heinisch, J.J. The Pentose Phosphate Pathway in Yeasts-More Than a Poor Cousin of Glycolysis. Biomolecules 2021, 11, 725. [Google Scholar] [CrossRef]
- Beere, H.M.; Wolf, B.B.; Cain, K.; Mosser, D.D.; Mahboubi, A.; Kuwana, T.; Tailor, P.; Morimoto, R.I.; Cohen, G.M.; Green, D.R. Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the Apaf-1 apoptosome. Nat. Cell Biol. 2000, 2, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gao, W.; Pan, Y.; Liu, G. Metabolic engineering of Acremonium chrysogenum for improving cephalosporin C production independent of methionine stimulation. Microb. Cell Factories 2018, 17, 87. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.; Gurr, S.J. Fungi, fungicide discovery and global food security. Fungal Genet. Biol. FGB 2020, 144, 103476. [Google Scholar] [CrossRef] [PubMed]
- Tishkov, V.I.; Popov, V.O. Protein engineering of formate dehydrogenase. Biomol Eng. 2006, 23, 89–110. [Google Scholar] [CrossRef]
- Dumestre, A.; Chone, T.; Portal, J.; Gerard, M.; Berthelin, J. Cyanide Degradation under Alkaline Conditions by a Strain of Fusarium solani Isolated from Contaminated Soils. Appl. Environ. Microbiol. 1997, 63, 2729–2734. [Google Scholar] [CrossRef]
- Leonhardt, Y.; Beck, J.; Ebel, F. Functional characterization of the Woronin body protein WscA of the pathogenic mold Aspergillus fumigatus. Int. J. Med. Microbiol. IJMM 2016, 306, 165–173. [Google Scholar] [CrossRef]
- Wang, Z.S.; Gu, Y.X.; Yuan, Q.S. Effect of nutrition factors on the synthesis of superoxide dismutase, catalase, and membrane lipid peroxide levels in Cordyceps militaris mycelium. Curr. Microbiol. 2006, 52, 74–79. [Google Scholar] [CrossRef]
- Raethong, N.; Wang, H.; Nielsen, J.; Vongsangnak, W. Optimizing cultivation of Cordyceps militaris for fast growth and cordycepin overproduction using rational design of synthetic media. Comput. Struct. Biotechnol. J. 2019, 18, 1–8. [Google Scholar] [CrossRef]
- Gu, C.; Cooper, D.M. Ca(2+), Sr(2+), and Ba(2+) identify distinct regulatory sites on adenylyl cyclase (AC) types VI and VIII and consolidate the apposition of capacitative cation entry channels and Ca(2+)-sensitive ACs. J. Biol. Chem. 2000, 275, 6980–6986. [Google Scholar] [CrossRef]
- Chen, X.; Wu, G.; Huang, Z. Structural analysis and antioxidant activities of polysaccharides from cultured Cordyceps militaris. Int. J. Biol. Macromol. 2013, 58, 18–22. [Google Scholar] [CrossRef]
- Yagura, T.; Nagata, I.; Kuma, K.; Uchino, H. Increased cyclic nucleotide phosphodiesterase (PDE) and calmodulin activities in soluble fraction of Graves’ thyroid: Analysis of increase in Ca2+ dependence of PDE activities. J. Clin. Endocrinol. Metab. 1985, 60, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Wang, X.; Wang, S.; Zhu, S.; Rong, R.; Xu, X. Complex effect of zinc oxide nanoparticles on cadmium chloride-induced hepatotoxicity in mice: Protective role of metallothionein. Metallomics 2017, 9, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Leach, M.D.; Klipp, E.; Cowen, L.E.; Brown, A.J. Fungal Hsp90: A biological transistor that tunes cellular outputs to thermal inputs. Nat. Rev..Microbiol. 2012, 10, 693–704. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).