Study of Fungal Communities in Dry Red Wine Fermentation in Linfen Appellation, Shanxi


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Methods
2.2.1. Wine Fermentation Process
2.2.2. Gene DNA Extraction from Wine Samples
2.2.3. PCR Amplification and MiSeq Sequencing of Wine Samples
2.3. Data Processing
3. Results
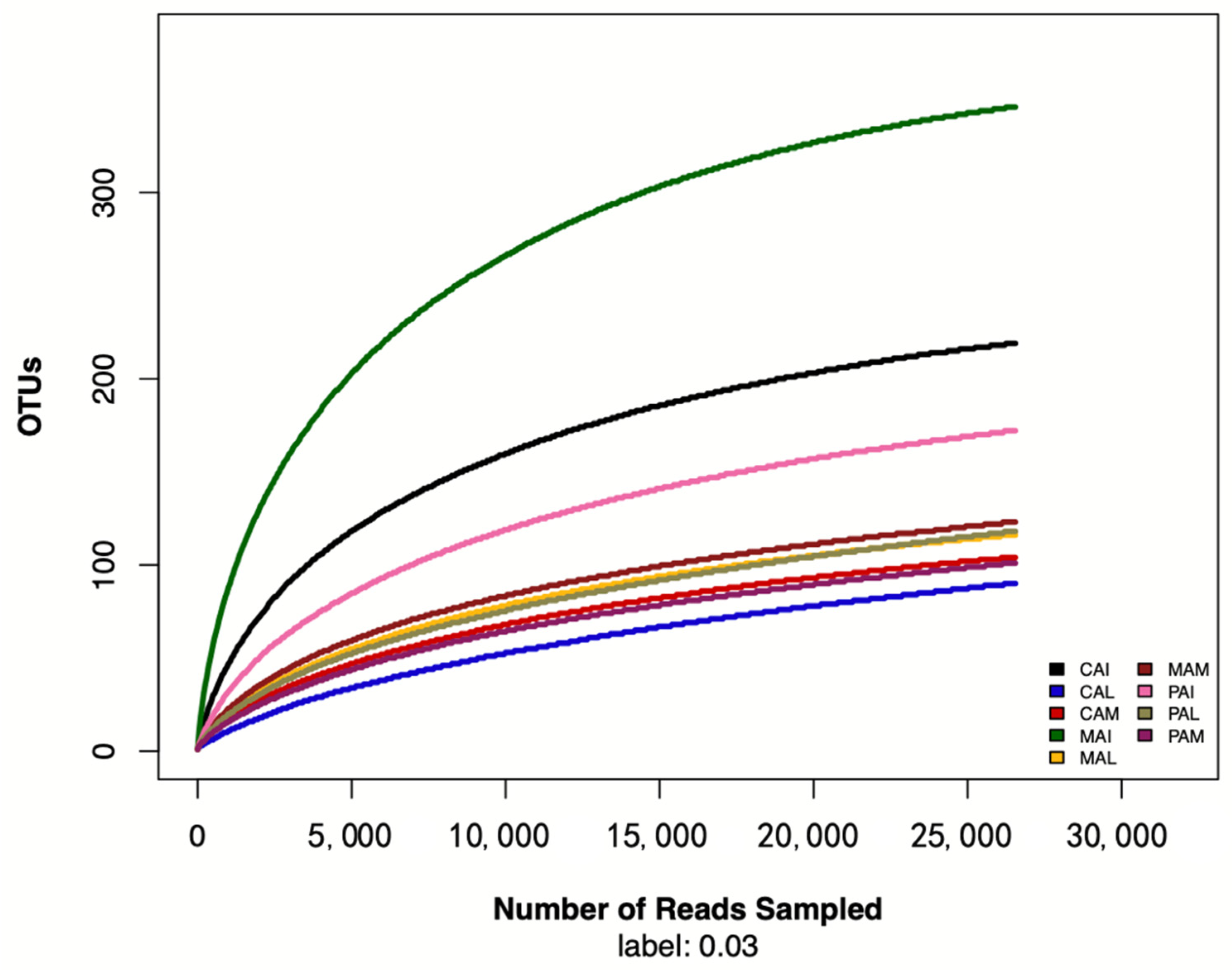
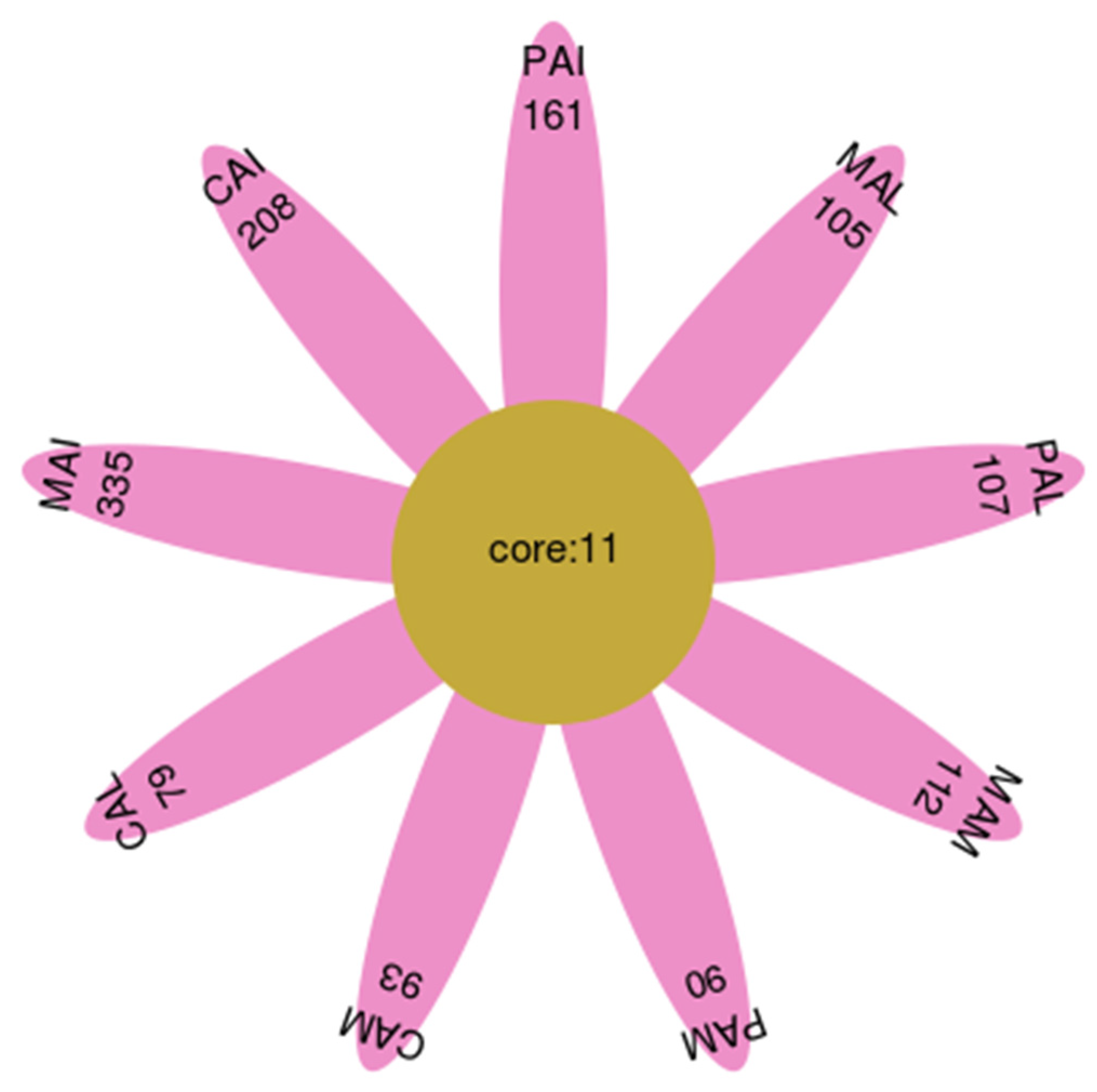
3.1. Sequence Data and OTUs Analysis
3.2. Alpha Diversity Analysis of Fungal Flora during Wine Fermentation
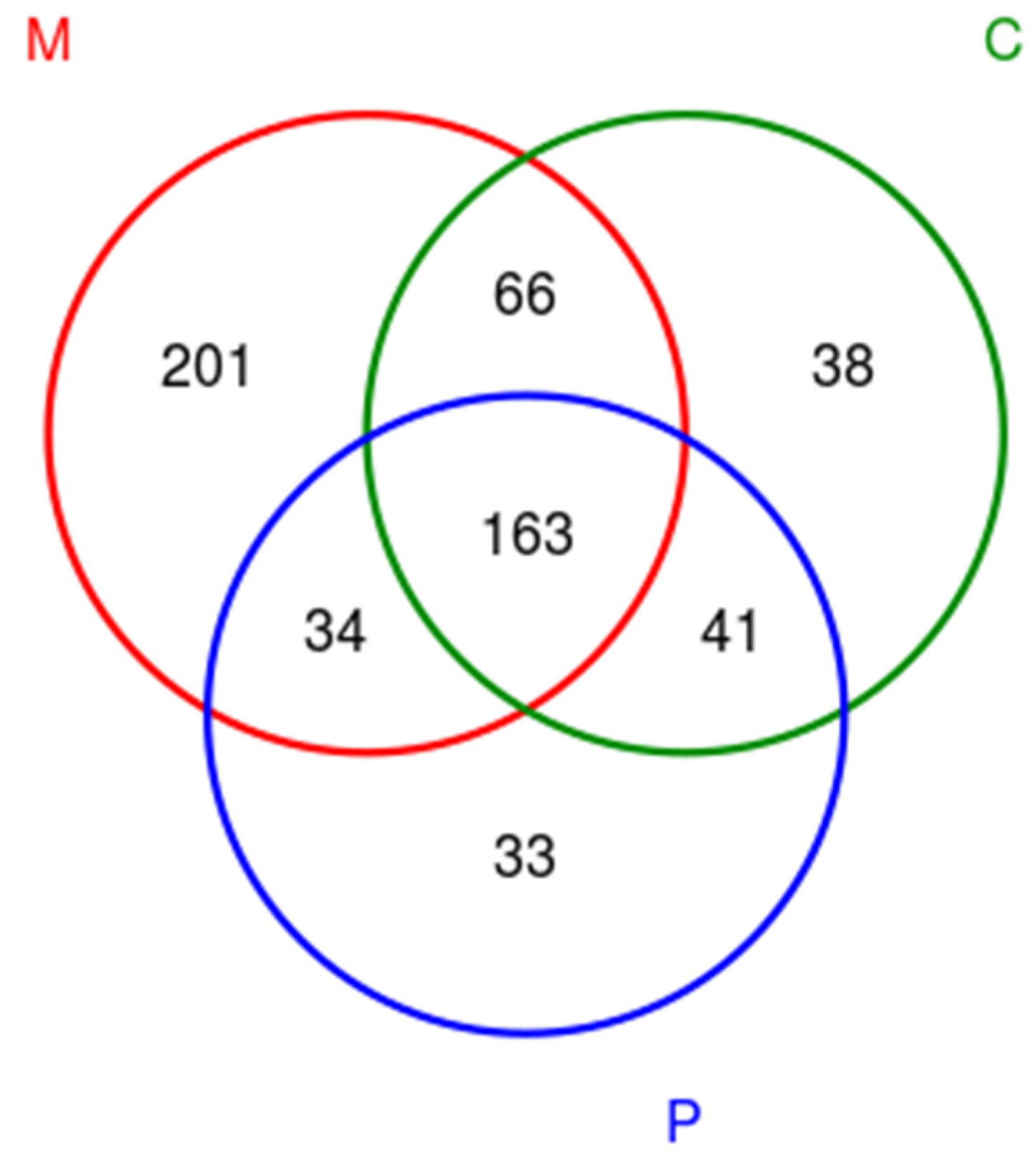
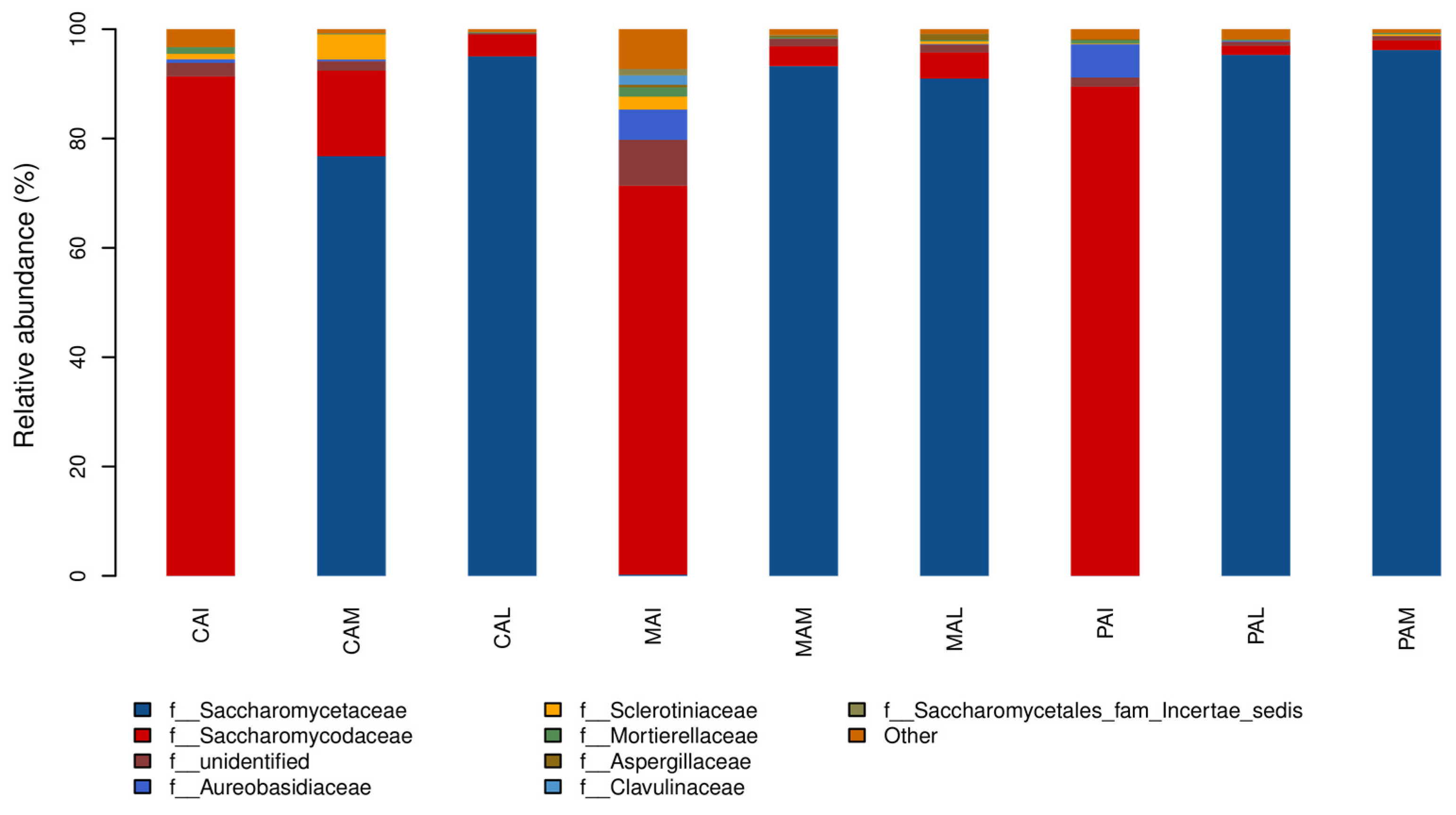
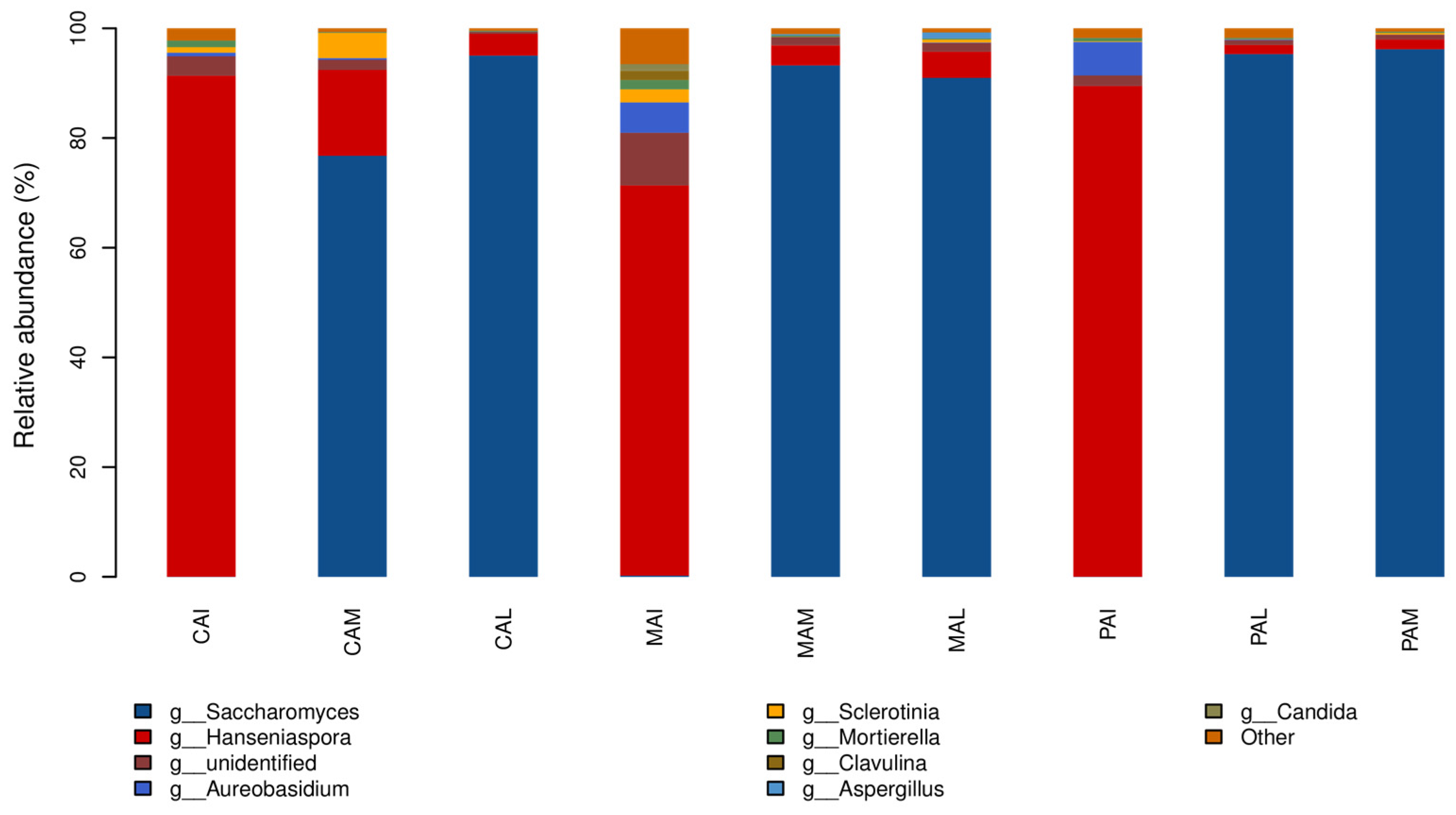
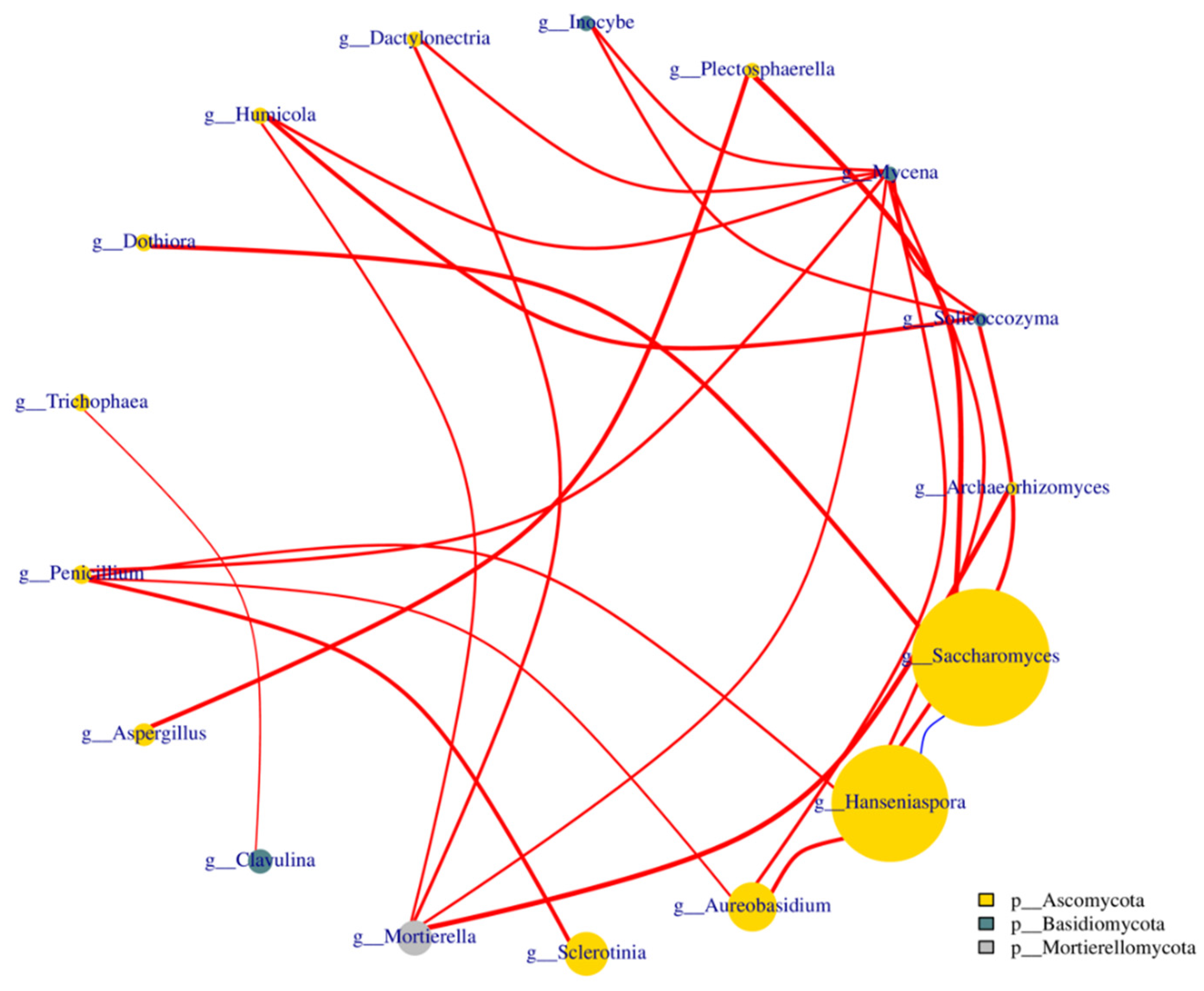
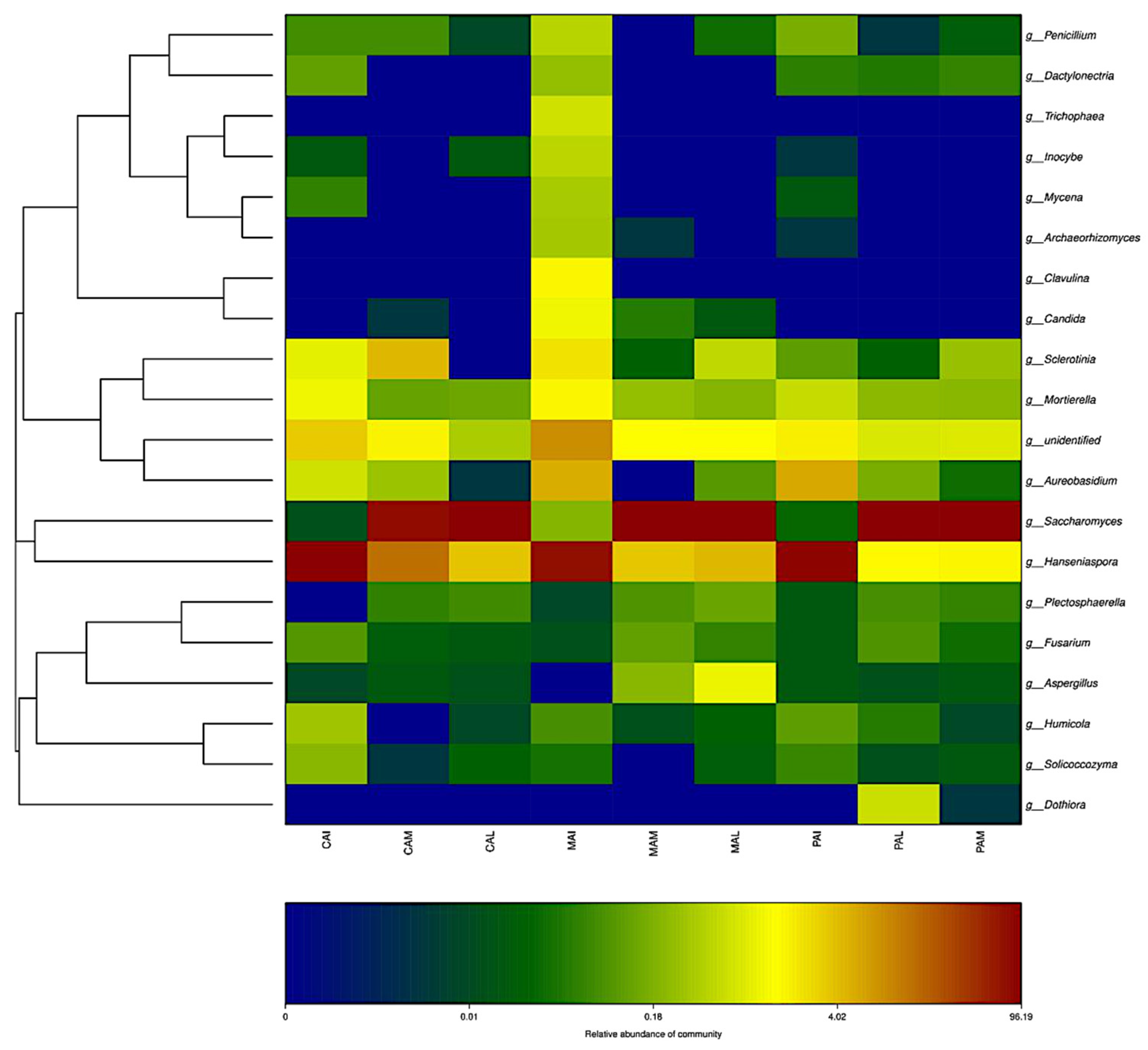
3.3. Taxonomic Distribution and Correlation Analysis of Fungi
3.4. Dynamic Changes of Fungal Flora
3.5. Beta Diversity Analysis of Fungal Flora during Wine Fermentation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maicas, S. Advances in Wine Fermentation. Fermentation 2021, 7, 187. [Google Scholar] [CrossRef]
- Knoll, C.; Fritsch, S.; Schnell, S.; Grossmann, M.; Krieger-Weber, S.; du Toit, M.; Rauhut, D. Impact of different malo-lactic fermentation inoculation scenarios on Riesling wine aroma. World J. Microbiol. Biotechnol. 2012, 28, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.T.; Ding, Y.T.; Gao, F.F.; Zhang, L.; Wang, L.; Li, H.; Wang, H. Community succession of the grape epidermis microbes of cabernet sauvignon (Vitis vinifera L.) from different regions in China during fruit development. Int. J. Food Microbiol. 2022, 362, 109475. [Google Scholar] [CrossRef] [PubMed]
- Luyt, N.A.; Beaufort, S.; Divol, B.; Setati, M.E.; Taillandier, P.; Bauer, F.F. Phenotypic characterization of cell-to-cell interactions between two yeast species during alcoholic fermentation. World J. Microbiol. Biotechnol. 2021, 37, 186. [Google Scholar] [CrossRef]
- Böhmer, M.; Smoľak, D.; Ženišová, K.; Čaplová, Z.; Pangallo, D.; Puškárová, A.; Bučková, M.; Cabicarová, T.; Budiš, J.; Šoltýs, K.; et al. Comparison of microbial diversity during two different wine fermentation processes. FEMS Microbiol. Lett. 2020, 367, fnaa150. [Google Scholar] [CrossRef]
- Hu, B.R.; Li, H. Analysis of aromatic composition in the different wine-grapes by gas chromatography-mass spectrometry. Food Ferment. Ind. 2005, 31, 89–92. [Google Scholar]
- Hu, B.R.; Yang, X.Y.; Wang, Z.J.; Hua, L. Analysis of aromatic composition in different mono-varietal dry wines from eastern region of Ningxia Helan mountain. Trans. Chin. Soc. Agric. Mach. 2005, 36, 87–90, 113. [Google Scholar]
- Jing, Y.; Zhu, Y.; Xu, Y. Mystery behind Chinese liquor fermentation. Trends Food Sci. Technol. 2017, 63, 18–28. [Google Scholar]
- Boran, H.U.; Jie, L.U.; Wenbiao, X.U.; Fengmin, Z. Comparative Study on the Changes of Aroma Components in the Grape and Dry Red Wine of Cabernet Sauvignon. Anim. Plant Sci. 2015, 25, 240–246. [Google Scholar]
- Wu, C.; Wang, C.; Wang, X.; Zhou, H.; Huang, Y.; Qiu, S. A review of the application of high-throughput sequencing technology in analysis of the diversity of microbial communities involved in the fermentation of alcoholic beverages. Food Sci. 2019, 40, 348–355. [Google Scholar]
- del Carmen Portillo, M.; Franquès, J.; Araque, I.; Reguant, C.; Bordons, A. Bacterial diversity of Grenache and Carignan grape surface from different vine-yards at Priorat wine region (Catalonia, Spain). Int. J. Food Microbiol. 2016, 219, 56–63. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, X.; Zhong, Q.; Zhuang, X.; Bai, Z. Microbial community analyses associated with nine varieties of wine grape carposphere based on high-throughput sequencing. Microorganisms 2019, 7, 668. [Google Scholar] [CrossRef] [PubMed]
- Setati, M.E.; Acobson, D.; Bauer, F.F. Sequence-based analysis of the Vitis vinifera L. cv Cabernet Sauvignon grape must mycobiome in three South African vineyards employing distinct agronomic systems. Front. Microbiol. 2015, 6, 1358. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.R.; Gao, J.; Xu, S.C.; Zhu, J.Y.; Fan, X.M.; Zhou, X.Y. Quality evaluation of different varieties of dry red wine based on nuclear magnetic resonance metabolomics. Appl. Biol. Chem. 2020, 63, 24. [Google Scholar] [CrossRef]
- Tang, X.Y. Analysis of Microbial Diversity of Naturally Fermented Soybean Paste Sauce Grains. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2017. [Google Scholar]
- Zhang, S.W.; Chen, X.; Zhong, Q.D.; Huang, Z.; Meng, Z.; Luo, J.; Shi, L.; Bai, Z. Analysis of microbial community diversity of different varieties of wine grape skins. Biotechnol. Bull. 2017, 33, 128–137. [Google Scholar]
- Mendoza, L.M.; Merín, M.G.; Morata, V.I.; Farías, M.E. Characterization of wines produced by mixed culture of autochthonous yeasts and Oenococcus oeni from the northwest region of Argentina. J. Ind. Microbiol. Biotechnol. 2011, 38, 1777–1785. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qi, G.M.; Wang, C.P. The role of non-enological yeasts in wine production and research progress. Chin. Foreign Grape Wine 2019, 1, 52–56. [Google Scholar]
- Liang, Z.; Lin, X.; He, Z.; Su, H.; Li, W.; Ren, X. Amino acid and microbial community dynamics during the fermentation of Hong Qu glutinous rice wine. Food Microbiol. 2020, 90, 103467. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Andorrà, I.; Landi, S.; Mas, A.; Esteve-Zarzoso, B.; Guillamón, J.M. Effect of fermentation temperature on microbial population evolution using culture-independent and dependent techniques. Food Res. Int. 2010, 43, 773–779. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Z.; Sun, J.; Ni, L. The dynamics of volatile compounds and their correlation with the microbial succession during the traditional solid-state fermentation of Gutian Hong Qu glutinous rice wine. Food Microbiol. 2020, 86, 103347. [Google Scholar] [CrossRef] [PubMed]
- Gabriela, M.M.; Inés, M.A.V. Application of a grape surface majority pectinolytic species, Aureobasidium pullulans, to low-temperature red winemaking: Development and stability of wine color. J. Wine Res. 2020, 31, 1816534. [Google Scholar]
- Mao, X.Y.; Chen, T.; Tian, H.; He, D. Metabolic study of arachidonic acid production by Persea Alpina. China Fats Oils 2011, 36, 42–44. [Google Scholar]
- Liao, P.; Wang, W.M.; Wang, W.C.; Kryczek, I.; Li, X.; Bian, Y.; Sell, A.; Wei, S.; Grove, S.; Johnson, J.K.; et al. CD8(+) T cells and fatty acids orchestrate tumor ferroptosis and immunity via ACSL4. Cancer Cell 2022, 40, 365. [Google Scholar] [CrossRef]
- Bosma, K.J.; Kaiser, C.E.; Kimple, M.E.; Gannon, M. Effects of Arachidonic Acid and Its Metabolites on Functional Beta-Cell Mass. Metabolites 2022, 12, 342. [Google Scholar] [CrossRef]
- Barbary, F.; Moshirian, N. The Modulation of Arachidonic Acid Metabolism and Blood pressure-lowering Effect of Honokiol in Spontaneously Hypertensive Rats. Molecules 2022, 27, 3396. [Google Scholar]
- Urbina, J.A.S.; García, J.A.A.; Terán, F.R. Yeast species are associated with a spontaneous fermentation of taberna, a traditional palm wine from the southeast of Mexico. Ann. Microbiol. 2015, 65, 287–296. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on using non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef]
- Kitagawa, T.; Tokuhiro, K.; Sugiyama, H.; Kohda, K.; Isono, N.; Hisamatsu, M.; Takahashi, H.; Imaeda, T. Construction of a β-glucosidase expression system using the multi-stress-tolerant yeast Issatchenkia orientalis. Appl. Microbiol. Biotechnol. 2010, 87, 1841–1853. [Google Scholar] [CrossRef]
- Kim, D.H.; Hong, Y.A.; Park, H.D. Co-fermentation of grape must by Issatchenkia Orientalis and Saccharomyces cerevisiae reduces the malic acid content in wine. Biotechnol. Lett. 2008, 30, 1633–1638. [Google Scholar] [CrossRef]
- Huang, Z.R.; Guo, W.L.; Zhou, W.B.; Li, L.; Xu, J.X.; Hong, J.L.; Liu, H.P.; Zeng, F.; Bai, W.D.; Liu, B.; et al. Microbial communities and volatile metabolites in different traditional fermentation starters used for Hong Qu glutinous rice wine. Food Res. Int. 2019, 121, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Martino, A.; Schiraldi, C.; Di Lazzaro, A.; Fiume, I.; Spagna, G.; Pifferi, P.G.; De Rosa, M. Improvement of the flavor of Falanghina white wine using a purified glycosidase preparation from Aspergillus niger. Process Biochem. 2000, 36, 93–102. [Google Scholar] [CrossRef]
- Hu, K.; Zhu, X.L.; Mu, H.; Ma, Y.; Ullah, N.; Tao, Y.S. A novel extracellular glycosidase activity from Rhodotorula mucilaginosa: Its application potential in wine aroma enhancement. Lett. Appl. Microbiol. 2016, 62, 169–176. [Google Scholar] [CrossRef]
- Abdeljalil, S.; Ben Hmad, I.; Saibi, W.; Amouri, B.; Maalej, W.; Kaaniche, M.; Koubaa, A.; Gargouri, A. Investigations on Hydrolytic Activities from Stachybotrys microspora and Their Use as an Alternative in Yeast DNA Extraction. Appl. Biochem. Biotechnol. 2014, 172, 1599–1611. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.L.; Wu, Y.; He, Q.; Xu, S.; Chen, Y. Analysis of microbial community diversity in traditional Korean rice wine and its liquor beverages based on high-throughput sequencing technology. Food Sci. 2019, 40, 107–114. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Length Gradient | Number of Sequencs |
|---|---|
| 0–200 | 1036 |
| 200–260 | 20,918 |
| 260–320 | 3463 |
| 320–360 | 432 |
| 360–380 | 92,895 |
| 380–400 | 31 |
| 400–420 | 45 |
| 420–440 | 242,204 |
| 440–460 | 271 |
| 460–480 | 21 |
| 480–500 | 26 |
| 500–520 | 28 |
| 520–540 | 15 |
| 540–560 | 0 |
| 560–600 | 0 |
| Name of Samples | Chao1 | Coverage | Observed_Species | PD_Whole_Tree | Shannon | Simpson |
|---|---|---|---|---|---|---|
| CAI | 245.56 | 1.00 | 219.00 | 48.51 | 0.96 | 0.17 |
| CAM | 129.62 | 1.00 | 104.00 | 26.94 | 1.20 | 0.38 |
| CAL | 133.00 | 1.00 | 90.00 | 25.79 | 0.38 | 0.10 |
| MAI | 371.54 | 1.00 | 346.00 | 76.67 | 2.50 | 0.49 |
| MAM | 173.17 | 1.00 | 123.00 | 34.22 | 0.60 | 0.13 |
| MAL | 153.05 | 1.00 | 116.00 | 27.52 | 0.73 | 0.17 |
| PAI | 210.15 | 1.00 | 172.00 | 37.31 | 0.86 | 0.20 |
| PAM | 144.05 | 1.00 | 101.00 | 26.12 | 0.38 | 0.07 |
| PAL | 194.56 | 1.00 | 118.00 | 26.48 | 0.47 | 0.09 |
| Name of Strans | CAI | CAM | CAL | MAI | MAM | MAL | PAI | PAM | PAL |
|---|---|---|---|---|---|---|---|---|---|
| H. uvarum | 91.4% | 15.6% | 3.96% | 71.2% | 3.66% | 4.78% | 89.5% | 1.79% | 1.67% |
| S. cerevisiae | 0.0113% | 76.8% | 95.0% | 0.207% | 93.2% | 91.0% | 0.0264% | 96. 2% | 95.3% |
| A. pullulans | 0.678% | 0.313% | 0.0038% | 5.54% | - | 0.102% | 6.10% | 0.0301% | 0.173% |
| M. alpina | 0.264% | 0.0113% | 0.0075% | 0.117% | - | 0.0226% | 0.0942% | 0.0226% | 0.0640% |
| C. tropicalis | - | - | - | 1.21% | 0.0226% | 0.0038% | - | - | - |
| I. orientalis | - | - | - | 0.0038% | 0.0113% | - | - | - | - |
| A. niger | - | 0.0038% | 0.0038% | - | 0.0603% | 0.478% | - | 0.0075% | - |
| R. mucilaginosa | - | - | - | - | 0.0113% | 0.0188% | - | - | - |
| R. sp | 0.0038% | - | - | 0.0414% | - | - | 0.0038% | - | - |
| R. diobovata | - | - | - | - | 0.0075% | 0.0414% | - | - | 0.0038% |
| S. microspora | - | - | - | - | 0.0001% | 0.0113% | - | - | - |
| G. pullulans | 0.0151% | 0.0113% | 0.0188% | - | 0.0188% | - | 0.0113% | 0.0075% | 0.0527% |
| Number of Yeast Colonies Formed Per mL of Fermentation Broth (108 CFU/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Yeast Species | CAI | CAM | CAL | MAI | MAM | MAL | PAI | PAM | PAL |
| H. uvarum | 5.18 | 0.887 | 0.225 | 4.03 | 0.207 | 0.271 | 5.07 | 0.101 | 0.0946 |
| S. cerevisiae | 0.0006 | 4.35 | 5.39 | 0.0117 | 5.29 | 5.16 | 0.0015 | 5.45 | 5.40 |
| C. tropicalis | - | - | - | 0.0683 | 0.0013 | 0.0002 | - | - | - |
| I. orientalis | - | - | - | 0.0002 | 0.0006 | - | - | - | - |
| R. sp | 0.0002 | - | - | 0.0023 | - | - | 0.0002 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, B.; Zhou, M.; Su, J.; Lin, L.; Xu, S. Study of Fungal Communities in Dry Red Wine Fermentation in Linfen Appellation, Shanxi. Fermentation 2022, 8, 475. https://doi.org/10.3390/fermentation8100475
Hu B, Zhou M, Su J, Lin L, Xu S. Study of Fungal Communities in Dry Red Wine Fermentation in Linfen Appellation, Shanxi. Fermentation. 2022; 8(10):475. https://doi.org/10.3390/fermentation8100475
Chicago/Turabian StyleHu, Boran, Min Zhou, Jinghao Su, Lan Lin, and Shaochen Xu. 2022. "Study of Fungal Communities in Dry Red Wine Fermentation in Linfen Appellation, Shanxi" Fermentation 8, no. 10: 475. https://doi.org/10.3390/fermentation8100475
APA StyleHu, B., Zhou, M., Su, J., Lin, L., & Xu, S. (2022). Study of Fungal Communities in Dry Red Wine Fermentation in Linfen Appellation, Shanxi. Fermentation, 8(10), 475. https://doi.org/10.3390/fermentation8100475