
Viability, Storage Stability and In Vitro Gastrointestinal Tolerance of Lactiplantibacillus plantarum Grown in Model Sugar Systems with Inulin and Fructooligosaccharide Supplementation



Abstract
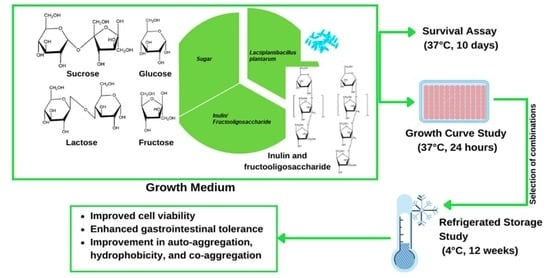
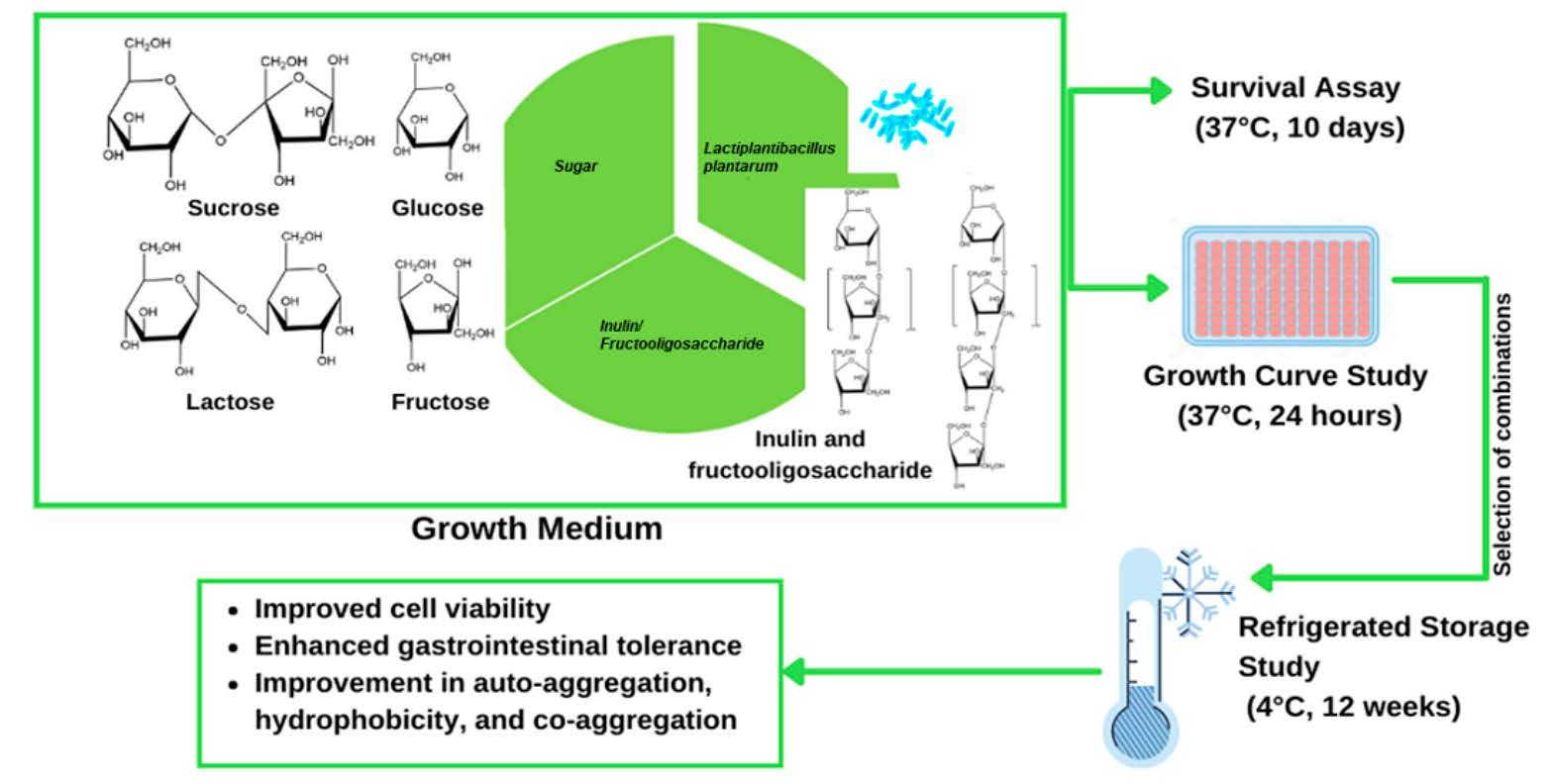
:
1. Introduction
2. Materials and Methods
2.1. Microbial Culture and Materials
2.2. Reactivation of Probiotic Culture and Preparation of Inoculum
2.3. Preparation of MRS Broth for L. plantarum
2.4. Growth Curve Study
2.5. Survival Study
2.6. Refrigerated Storage Study
2.6.1. Storage Viability and pH
2.6.2. Sugar and Organic Acid Analysis
2.6.3. Ethanol Analysis
2.6.4. Acid Tolerance Assay
2.6.5. Cell Auto-Aggregation and Co-Aggregation
2.6.6. Hydrophobicity of Bacteria
2.6.7. Gastrointestinal Tolerance Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. Growth Study
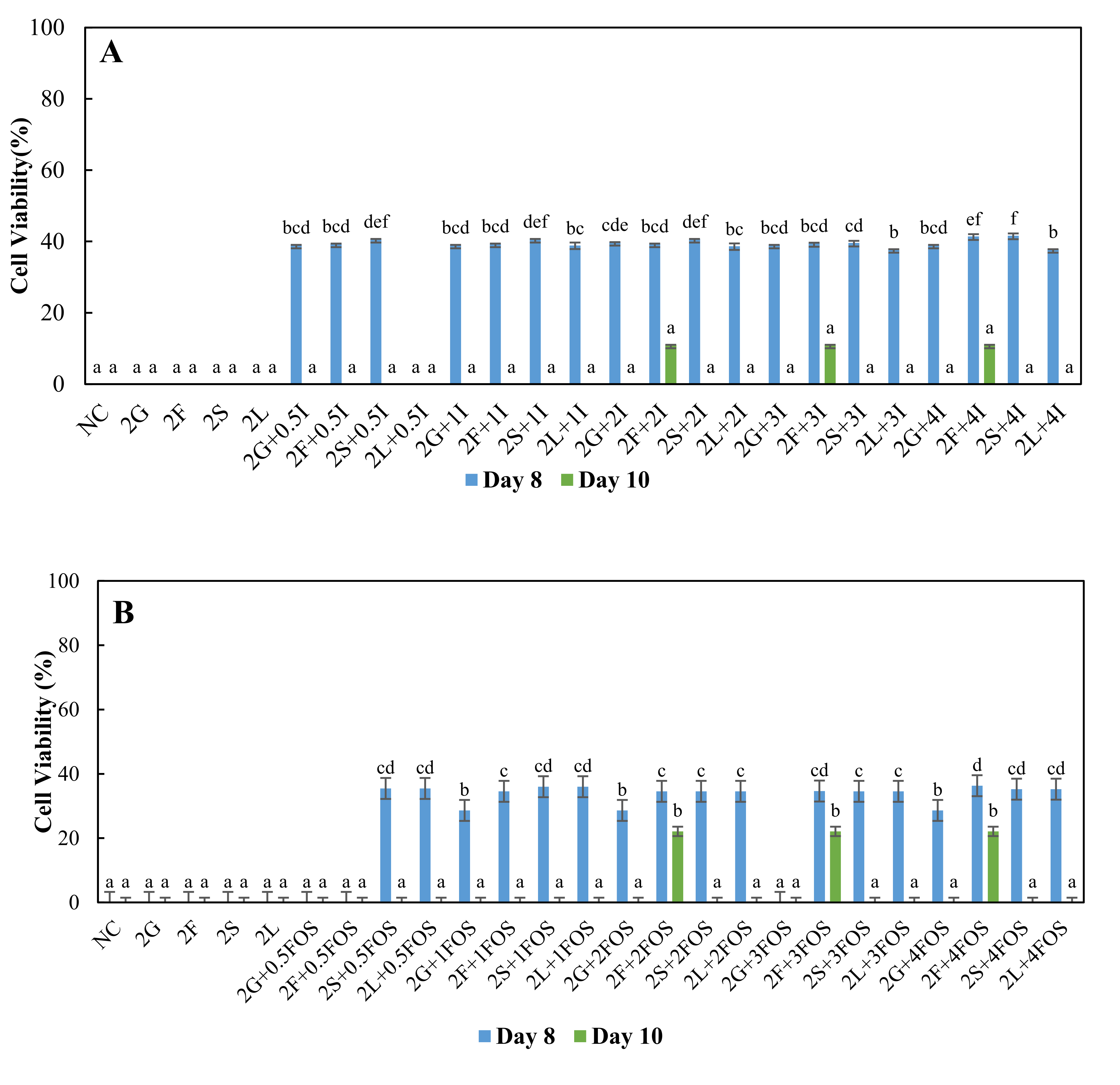
3.2. Comparison of the Survival of L. plantarum in Different Model Sugar Systems with Inulin and FOS Supplementation on Day-8 and Day-10
3.3. Effect of Inulin and FOS Supplementation on Storage Viability (log CFU/mL) and pH during 12 Weeks of Storage at 4 °C

{kind=link}
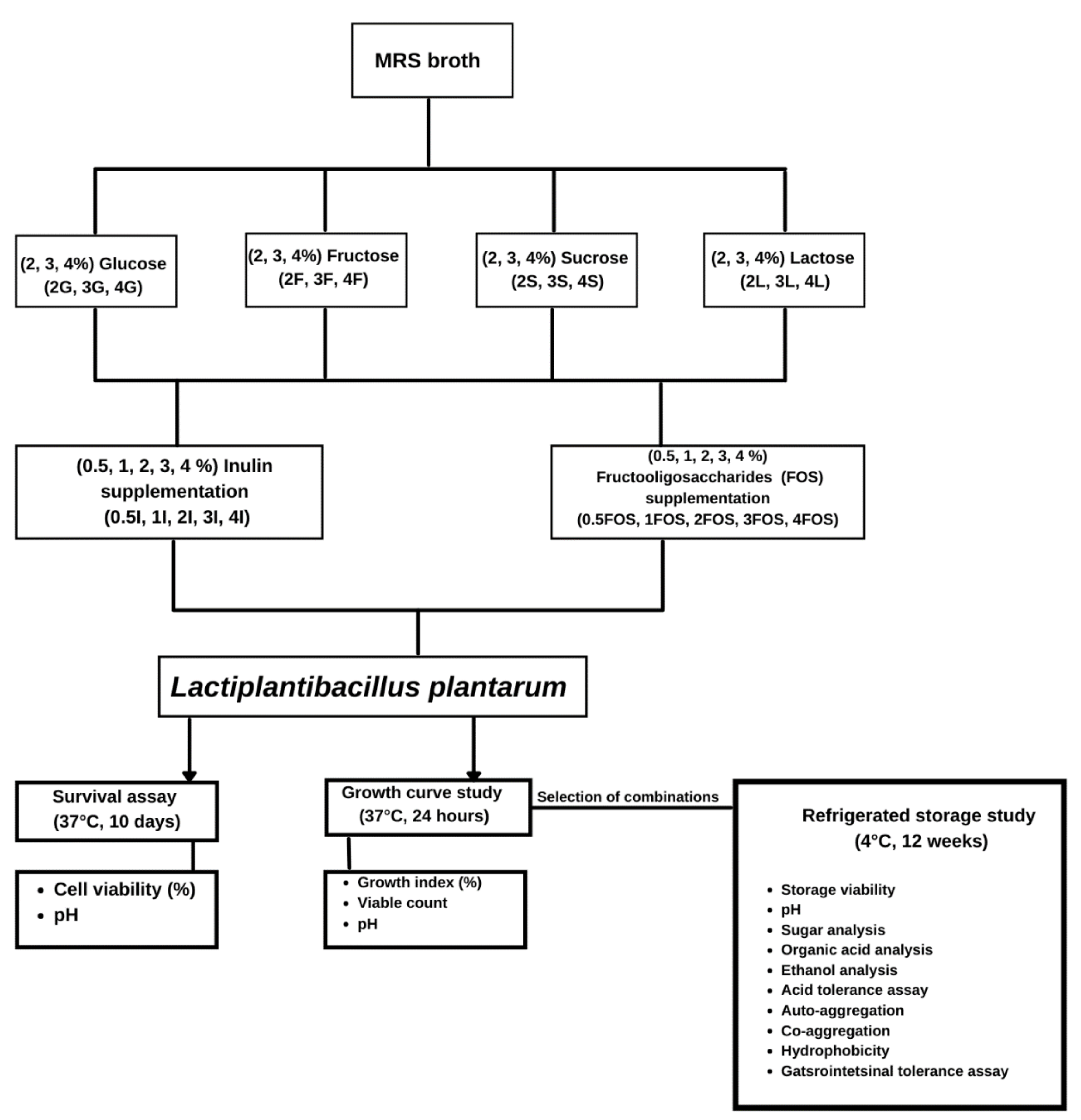{kind=link}
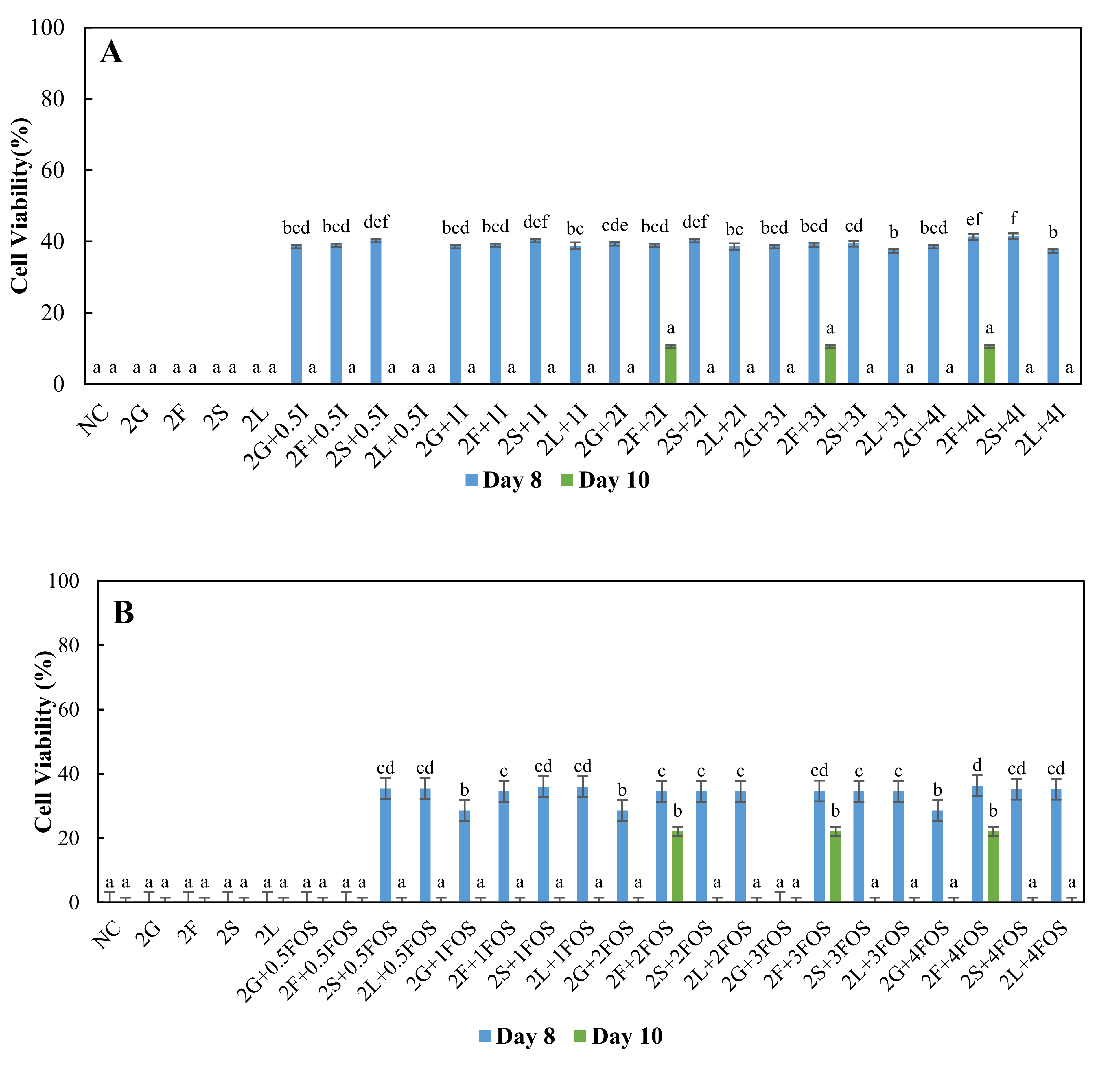{kind=link}
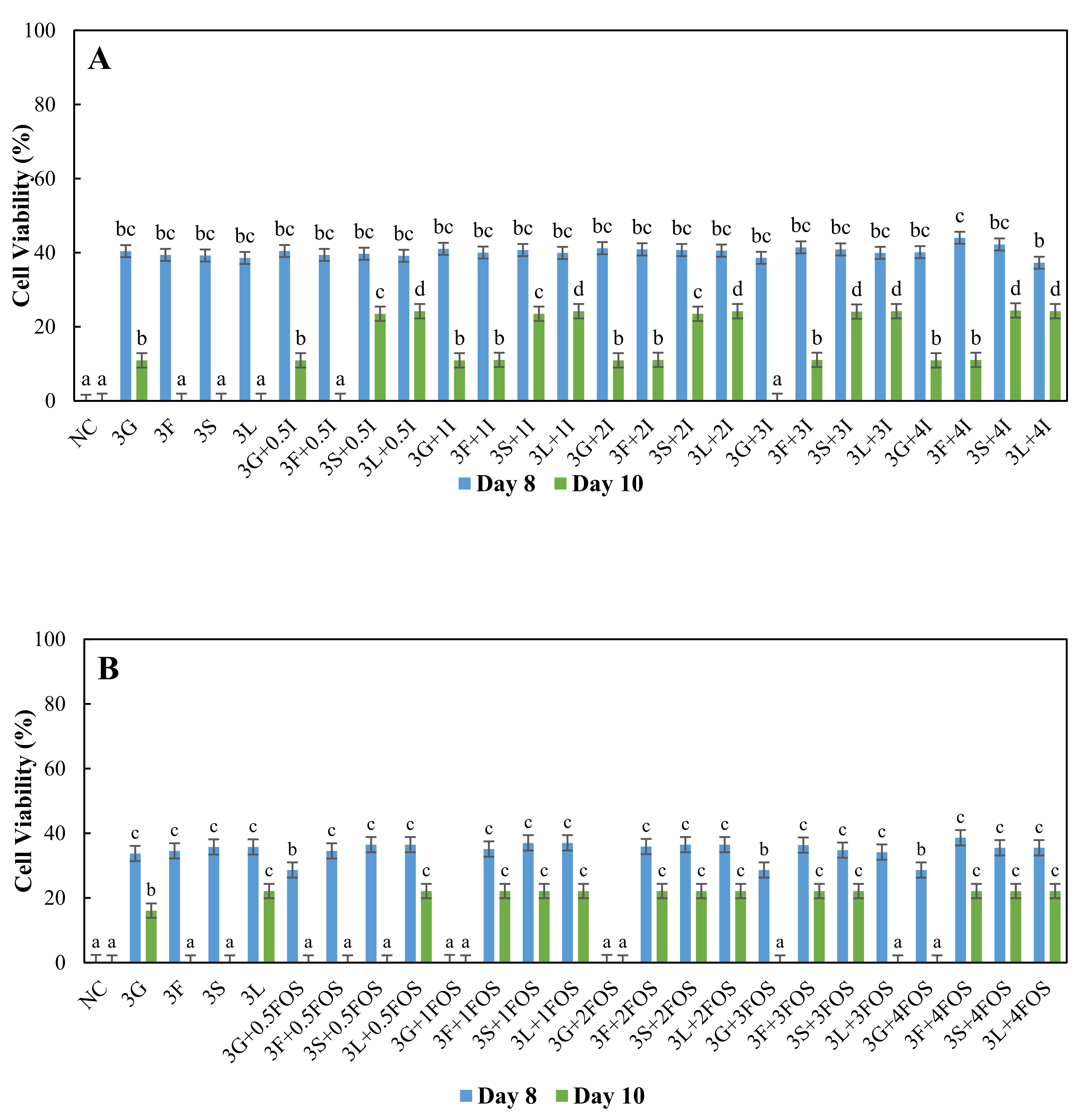{kind=link}
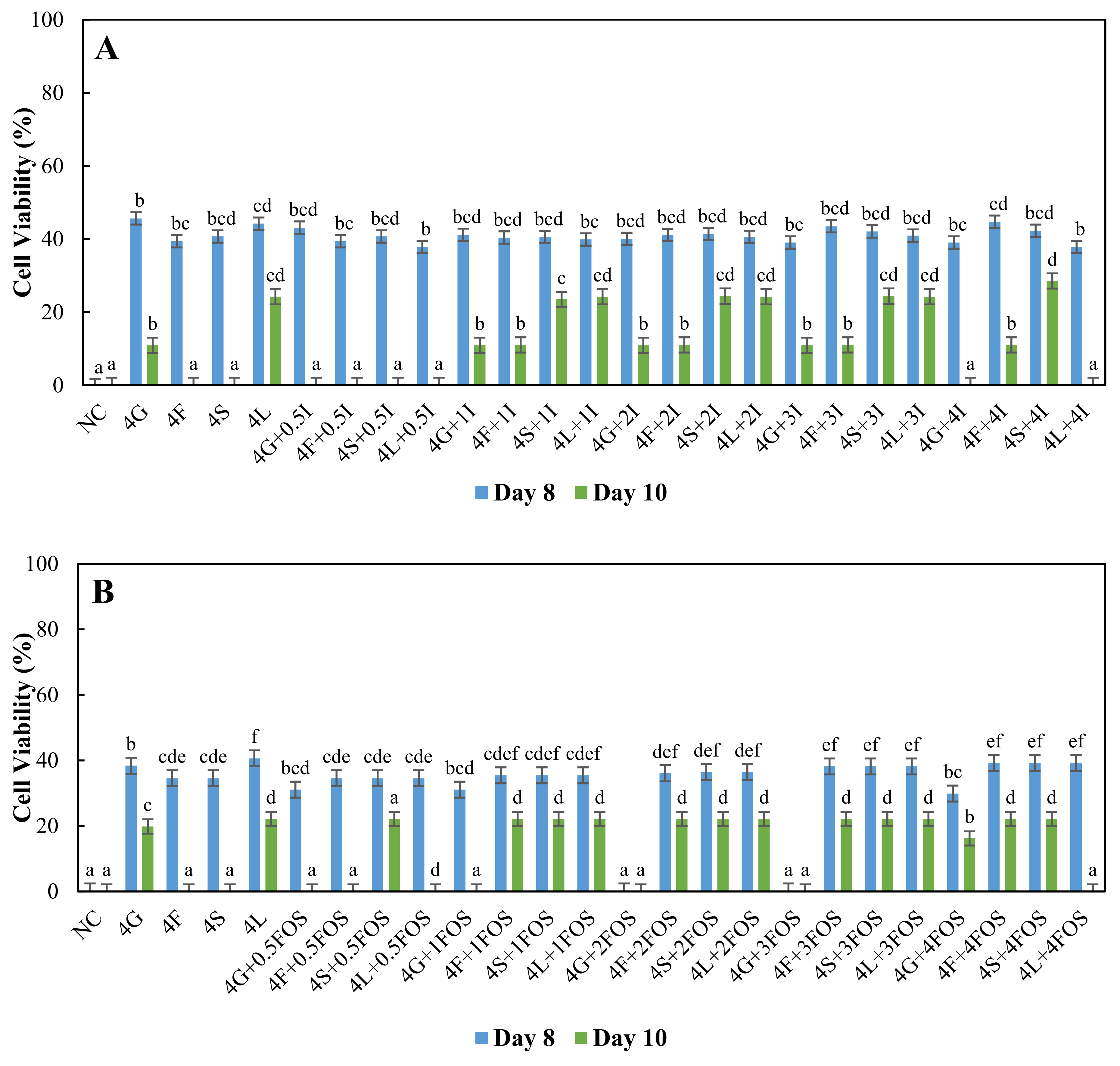{kind=link}
| Storage Viability (log CFU/mL) | pH Week 12 | |||
|---|---|---|---|---|
| Week 0 | Week 12 | Week 0 | Week 12 | |
| NC | Aa 7.6 ± 0.1 | Aa 0.0 ± 0.0 | BCDa 6.5 ± 0.1 | Db 7.1 ± 0.1 |
| 3S | Ca 9.3 ± 0.1 | BCb 5.6 ± 0.0 | CDa 6.7 ± 0.1 | Cb 6.7 ± 0.1 |
| 2L | Ca 9.2 ± 0.1 | Bb 4.6 ± 0.0 | CDa 6.6 ± 0.1 | Db 7.2 ± 0.1 |
| 3L | Ca 9.1 ± 0.5 | Bb 4.8 ± 0.0 | Ea 6.8 ± 0.1 | Db 7.2 ± 0.1 |
| 3I | Ba 8.2 ± 0.3 | Db 6.9 ± 0.0 | BCa 6.2 ± 0.2 | BCb 6.9 ± 0.1 |
| 4I | Ba 8.7 ± 0.2 | Cb 6.7 ± 0.1 | Ba 5.9 ± 0.2 | ABb 6.4 ± 0.2 |
| 2FOS | Ba 8.7 ± 0.2 | Cb 6.7 ± 0.1 | ABa 5.9 ± 0.1 | BCb 6.6 ± 0.2 |
| 4FOS | Ba 8.3 ± 0.1 | Db 7.1 ± 0.0 | Aa 5.5 ± 0.2 | ABb 6.4 ± 0.0 |
| 3S+4I | Da 10.4 ± 0.1 | Eb 8.1 ± 0.0 | Aa 5.6 ± 0.3 | Ab 5.9 ± 0.4 |
| 2L+3I | Da 10.4 ± 0.1 | Eb 8.1 ± 0.0 | ABa 5.6 ± 0.1 | ABb 6.4 ± 0.0 |
| 3S+2FOS | Da 10.2 ± 0.0 | Eb 8.2 ± 0.0 | Aa 5.3 ± 0.1 | ABb 6.5 ± 0.1 |
| 3L+4FOS | Da 10.4 ± 0.1 | Eb 8.2 ± 0.7 | Aa 5.8 ± 0.1 | ABb 6.3 ± 0.0 |


3.4. Sugar and Organic Acids
3.5. Auto-Aggregation, Hydrophobicity, and Co-Aggregation
3.6. Gastrointestinal Tolerance Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stewart, M.L.; Timm, D.A.; Slavin, J.L. Fructooligosaccharides exhibit more rapid fermentation than long-chain inulin in an in vitro fermentation system. Nutr. Res. 2008, 28, 329–334. [Google Scholar] [CrossRef]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2017, 137, 830S–837S. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Bayarri, S.; Tárrega, A.; Costell, E. Inulin as texture modifier in dairy products. Food Hydrocoll. 2011, 25, 1881–1890. [Google Scholar] [CrossRef]
- Van Den Ende, W.; Peshev, D.; De Gara, L. Disease prevention by natural antioxidants and prebiotics acting as ROS scavengers in the gastrointestinal tract. Trends Food Sci. Technol. 2011, 22, 689–697. [Google Scholar] [CrossRef]
- Peshev, D.; Van Den Ende, W. Fructans: Prebiotics and immunomodulators. J. Funct. Foods 2014, 8, 348–357. [Google Scholar] [CrossRef]
- Oliveira, R.P.; Florence, A.C.; Silva, R.C.; Perego, P.; Converti, A.; Gioielli, L.A.; Oliveira, M.N. Effect of different prebiotics on the fermentation kinetics, probiotic survival and fatty acids profiles in nonfat symbiotic fermented milk. Int. J. Food Microbiol. 2009, 128, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Wu, L.; Wu, Z.; Pan, D.; Zeng, X.; Guo, Y.; Lian, L. Effects of oligosaccharides on the fermentation properties of Lactobacillus plantarum. J. Dairy Sci. 2019, 102, 2863–2872. [Google Scholar] [CrossRef]
- Delavari, M.; Pourahmad, R.; Sokutifar, R. Production of low fat synbiotic yogurt containing Lactobacillus plantarum and inulin. Adv. Environ. Biol. 2014, 8, 17–24. [Google Scholar]
- Kariyawasam, K.M.G.M.M.; Lee, N.K.; Paik, H.D. Synbiotic yoghurt supplemented with novel probiotic Lactobacillus brevis KU200019 and fructooligosaccharides. Food Biosci. 2021, 39, 100835. [Google Scholar] [CrossRef]
- Bao, Y.; Zhang, Y.; Li, H.; Liu, Y.; Wang, S.; Dong, X.; Su, F.; Yao, G.; Sun, T.; Zhang, H. In vitro screen of Lactobacillus plantarum as probiotic bacteria and their fermented characteristics in soymilk. Ann. Microbiol. 2021, 62, 1311–1320. [Google Scholar] [CrossRef]
- Amanda, E.; Choo, W.S. Effect of refrigerated storage on the physicochemical characteristics and viability of Lactobacillus plantarum in fermented watermelon juice with or without supplementation with inulin or fructooligosaccharide. J. Food Process. Preserv. 2018, 42, 13831. [Google Scholar] [CrossRef]
- Russo, P.; de Chiara, M.L.V.; Capozzi, V.; Arena, M.P.; Amodio, M.L.; Rascón, A.; Dueñas, M.T.; López, P.; Spano, G. Lactobacillus plantarum strains for multifunctional oat-based foods. LWT-Food Sci. Technol. 2016, 68, 288–294. [Google Scholar] [CrossRef]
- Speranza, B.; Campaniello, D.; Monacis, N.; Bevilacqua, A.; Sinigaglia, M.; Corbo, M.R. Functional cream cheese supplemented with Bifidobacterium animalis subsp. lactis DSM 10140 and Lactobacillus reuteri DSM 20016 and prebiotics. Food Microbiol. 2018, 72, 16–22. [Google Scholar] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.C.; Vaughan, E.E.; Kleerebezem, M.; de Vos, W.M. Lactobacillus plantarum—survival, functional and potential probiotic properties in the human intestinal tract. Int. Dairy J. 2006, 16, 1018–1028. [Google Scholar] [CrossRef]
- Didari, T.; Solki, S.; Mozaffari, S.; Nikfar, S.; Abdollahi, M. A systematic review of the safety of probiotics. Expert Opin. Drug Saf. 2014, 13, 227–239. [Google Scholar] [CrossRef]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.; Santiago, A.; Teixeira, J.A.; Venâncio, A.; Abrunhosa, L. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Ren, L.Q.; Zhou, Y.; Ye, B.C. Characterization of antimicrobial activity of three Lactobacillus plantarum strains isolated from Chinese traditional dairy food. Food Sci. Nutr. 2019, 7, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.V.; Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Chobert, J.M.; Haertlé, T.; Franco, B.D.G.M. Characterization of a two-peptide plantaricin produced by Lactobacillus plantarum MBSa4 isolated from Brazilian salami. Food Control 2016, 60, 103–112. [Google Scholar] [CrossRef]
- Kwak, M.K.; Liu, R.; Kang, S.O. Antimicrobial activity of cyclic dipeptides produced by Lactobacillus plantarum LBP-K10 against multidrug-resistant bacteria, pathogenic fungi, and influenza A virus. Food Control 2018, 85, 223–234. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, B.; Tomar, S.K.; Chauhan, A. Techno-functional differentiation of two vitamin B 12 producing Lactobacillus plantarum strains: An elucidation for diverse future use. Appl. Microbiol. Biotechnol. 2017, 101, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Hugenschmidt, S.; Schwenninger, S.M.; Lacroix, C. Concurrent high production of natural folate and vitamin B12 using a co-culture process with Lactobacillus plantarum SM39 and Propionibacterium freudenreichii DF13. Process. Biochem. 2011, 46, 1063–1070. [Google Scholar] [CrossRef]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and characterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef]
- Alexander, C.; Swanson, K.S.; Fahey, G.C., Jr.; Garleb, K.A. Perspective: Physiologic Importance of Short-Chain Fatty Acids from Nondigestible Carbohydrate Fermentation. Adv. Nutr. 2019, 10, 576–589. [Google Scholar] [CrossRef]
- Cebeci, A.; Gürakan, C. Properties of potential probiotic Lactobacillus plantarum strains. Food Microbiol. 2003, 20, 511–518. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365s–373s. [Google Scholar] [CrossRef] [PubMed]
- Plumed-Ferrer, C.; Koistinen, K.M.; Tolonen, T.L.; Lehesranta, S.J.; Karenlampi, S.O.; Makimattila, E.; Joutsjoki, V.; Virtanen, V.; Von Wright, A. Comparative study of sugar fermentation and protein expression patterns of two Lactobacillus plantarum strains grown in three different media. Appl. Environ. Microbiol. 2008, 74, 5349–5358. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhou, F.; Ren, J.; Ai, L.; Dong, Y.; Wu, Z.; Liu, Z.; Chen, W.; Guo, B. Cloning, expression and functional validation of a β-fructofuranosidase from Lactobacillus plantarum. Process. Biochem. 2014, 49, 758–767. [Google Scholar] [CrossRef]
- Goh, Y.J.; Lee, J.H.; Hutkins, R.W. Functional analysis of the fructooligosaccharide utilization operon in Lactobacillus paracasei 1195. Appl. Environ. Microbiol. 2007, 73, 5716–5724. [Google Scholar] [CrossRef]
- Barrangou, R.; Altermann, E.; Hutkins, R.; Cano, R.; Klaenhammer, T.R. Functional and comparative genomic analyses of an operon involved in fructooligosaccharide utilization by Lactobacillus acidophilus. Proc. Natl. Acad. Sci. USA 2003, 100, 8957–8962. [Google Scholar] [CrossRef]
- Kaplan, H.; Hutkins, R.W. Metabolism of fructooligosaccharides by Lactobacillus paracasei 1195. Appl. Environ. Microbiol. 2003, 69, 2217–2222. [Google Scholar] [CrossRef]
- Saulnier, D.M.; Molenaar, D.; de Vos, W.M.; Gibson, G.R.; Kolida, S. Identification of prebiotic fructooligosaccharide metabolism in Lactobacillus plantarum WCFS1 through microarrays. Appl. Environ. Microbiol. 2007, 73, 1753–1765. [Google Scholar] [CrossRef]
- Parhi, P.; Song, K.P.; Choo, W.S. Effect of inulin and fructooligosaccharide supplementation on the growth and survival of Lactobacillus casei in model sugar systems. J. Food Process. Preserv. 2021, 45, 15228. [Google Scholar] [CrossRef]
- Ramos, C.L.; Thorsen, L.; Schwan, R.F.; Jespersen, L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 2013, 36, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Sada, A.; Orlando, P. Synbiotic potential of carrot juice supplemented with Lactobacillus spp. and inulin or fructooligosaccharides. J. Sci. Food Agric. 2008, 88, 2271–2276. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Perricone, M.; Cannarsi, M.; Corbo, M.R.; Sinigaglia, M. Technological and spoiling characteristics of the yeast microflora isolated from Bella di Cerignola table olives. Int. J. Food Sci. Technol. 2009, 44, 2198–2207. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Sinigaglia, M.; Speranza, B.; Altieri, C. Effect of prebiotic compounds on the growth and survival of bifidobacteria in a laboratory medium. Adv. J. Food Sci. Technol. 2016, 11, 770–774. [Google Scholar] [CrossRef]
- Kos, B.V.Z.E.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Kimoto-Nira, H.; Suzuki, C.; Sasaki, K.; Kobayashi, M.; Mizumachi, K. Survival of a Lactococcus lactis strain varies with its carbohydrate preference under in vitro conditions simulated gastrointestinal tract. Int. J. Food Microbiol. 2010, 143, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Valero-Cases, E.; Frutos, M.J. Effect of different types of encapsulation on the survival of Lactobacillus plantarum during storage with inulin and in vitro digestion. LWT-Food Sci. Technol. 2015, 64, 824–828. [Google Scholar] [CrossRef]
- Da Silva Sabo, S.; Converti, A.; Todorov, S.D.; Domínguez, J.M.; de Souza Oliveira, R.P. Effect of inulin on growth and bacteriocin production by Lactobacillus plantarum in stationary and shaken cultures. Int. J. Food Sci. Technol. 2015, 50, 864–870. [Google Scholar] [CrossRef]
- Munoz, M.; Mosquera, A.; Almeciga-Diaz, C.J.; Melendez, A.P.; Sanchez, O.F. Fructooligosaccharides metabolism and effect on bacteriocin production in Lactobacillus strains isolated from ensiled corn and molasses. Anaerobe 2012, 18, 321–330. [Google Scholar] [CrossRef]
- Oliveira, R.P.D.S.; Perego, P.; De Oliveira, M.N.; Converti, A. Effect of inulin as a prebiotic to improve growth and counts of a probiotic cocktail in fermented skim milk. LWT-Food Sci. Technol. 2011, 44, 520–523. [Google Scholar] [CrossRef]
- Perrin, S.; Warchol, M.; Grill, J.P.; Schneider, F. Fermentations of fructo-oligosaccharides and their components by Bifidobacterium infantis ATCC 15697 on batch culture in semi-synthetic medium. J. Appl. Microbiol. 2001, 90, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Leh, M.B.; Charles, M. Lactic acid production by batch fermentation of whey permeate: A mathematical model. J. Ind. Microbiol. Biotechnol. 1989, 4, 65–70. [Google Scholar] [CrossRef]
- Zhang, X.; Hao, X.; Wang, H.; Li, X.; Liu, L.; Yang, W.; Zhao, M.; Wang, L.; Bora, A.F.M. The effects of Lactobacillus plantarum combined with inulin on the physicochemical properties and sensory acceptance of low-fat Cheddar cheese during ripening. Int. Dairy J. 2021, 115, 104947. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of Probiotic Lactobacilli in Acidic Environments Is Enhanced in the Presence of Metabolizable Sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef]
- Hedberg, M.; Hasslöf, P.; Sjöström, I.; Twetman, S.; Stecksén-Blicks, C. Sugar fermentation in probiotic bacteria—An in vitro study. Oral Microbiol. Immunol. 2008, 23, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Kneifel, W. In vitro growth behaviour of probiotic bacteria in culture media with carbohydrates of prebiotic importance. Microb. Ecol. Health Dis. 2000, 12, 27–34. [Google Scholar]
- Deutscher, J.; Galinier, A.; Martin-Verstraete, I. Carbohydrate uptake and metabolism. In Bacillus Subtilis and Its Closest Relatives: From Genes to Cells; Sonenshein, A.L., Hoch, J.A., Losick, R., Eds.; ASM Press: Washington, DC, USA, 2001; pp. 129–150. [Google Scholar]
- Titgemeyer, F.; Hillen, W. Global control of sugar metabolism: A gram-positive solution. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Siezen, R.J., Kok, J., Abee, T., Schasfsma, G., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 59–71. [Google Scholar]
- Vadeboncoeur, C.; Pelletier, M. The phosphoenolpyruvate: Sugar phosphotransferase system of oral streptococci and its role in the control of sugar metabolism. FEMS Microbiol. Rev. 1997, 19, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Huebner, J.; Wehling, R.L.; Hutkins, R.W. Functional activity of commercial prebiotics. Int. Dairy J. 2007, 17, 770–775. [Google Scholar] [CrossRef]
- Krasaekoopt, W.; Watcharapoka, S. Effect of addition of inulin and galactooligosaccharide on the survival of microencapsulated probiotics in alginate beads coated with chitosan in simulated digestive system, yogurt and fruit juice. LWT-Food Sci. Technol. 2014, 57, 761–766. [Google Scholar] [CrossRef]
- Livingston, D.P., III; Henson, C.A. Apoplastic sugars, fructans, fructan exohydrolase, and invertase in winter oat: Responses to second-phase cold hardening. Plant Physiol. 1998, 116, 403–408. [Google Scholar] [CrossRef]
- Vereyken, I.J.; Chupin, V.; Islamov, A.; Kuklin, A.; Hincha, D.K.; de Kruijff, B. The effect of fructan on the phospholipid organization in the dry state. Biophys. J. 2003, 85, 3058–3065. [Google Scholar] [CrossRef]
- Honda, H.; Yajima, N.; Saito, T. Characterization of lactose utilization and β-Galactosidase in Lactobacillus brevis KB290, the hetero-fermentative lactic acid bacterium. Curr. Microbiol. 2012, 65, 679–685. [Google Scholar] [CrossRef]
- Srinivas, D.; Mital, B.K.; Garg, S.K. Utilization of sugars by Lactobacillus acidophilus strains. Int. J. Food Microbiol. 1990, 10, 51–57. [Google Scholar] [CrossRef]
- Goldin, B.R.; Gorbach, S.L.; Saxelin, M.; Barakat, S.; Gualtieri, L.; Salminen, S. Survival of Lactobacillus species (strain GG) in human gastrointestinal tract. Dig. Dis. Sci. 1992, 37, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Haukioja, A.; Söderling, E.; Tenovuo, J. Acid production from sugars and sugar alcohols by probiotic lactobacilli and bifidobacteria in vitro. Caries Res. 2008, 42, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl. Environ. Microbiol. 2014, 80, 2206–2215. [Google Scholar] [CrossRef]
- Neysens, P.; Messens, W.; Gevers, D.; Swings, J.; De Vuyst, L. Biphasic kinetics of growth and bacteriocin production with Lactobacillus amylovorus DCE 471 occur under stress conditions. Microbiology 2003, 149, 1073–1082. [Google Scholar] [CrossRef]
- De Vuyst, L. Technology aspects related to the application of functional starter cultures. Food Technol. Biotechnol. 2000, 38, 105–112. [Google Scholar]
- Sodini, I.; Lucas, A.; Oliveira, M.N.; Remeuf, F.; Corrieu, G. Effect of milk base and starter culture on acidification, texture, and probiotic cell counts in fermented milk processing. J. Dairy Sci. 2002, 85, 2479–2488. [Google Scholar] [CrossRef]
- Buntin, N.; Hongpattarakere, T.; Ritari, J.; Douillard, F.P.; Paulin, L.; Boeren, S.; Shetty, S.A.; de Vos, W.M. An inducible operon is involved in inulin utilization in Lactobacillus plantarum strains, as revealed by comparative proteogenomic and metabolic profiling. Appl. Environ. Microbiol. 2017, 83, e02402-16. [Google Scholar] [CrossRef]
- Desai, A.B.; Powell, I.B.; Shah, N.P. Survival and activity of probiotic lactobacilli in skim milk containing prebiotics. J. Funct. Foods 2004, 69, 57–60. [Google Scholar] [CrossRef]
- Bruno, F.A.; Lankaputhra, W.E.V.; Shah, N.P. Growth, viability and activity of Bifidobacterium spp. in skim milk containing prebiotics. J. Funct. Foods 2002, 67, 2740–2744. [Google Scholar] [CrossRef]
- Fugelsang, K.C. The lactic acid bacteria. In Wine Microbiology; Springer: Boston, MA, USA, 1997; pp. 3–47. [Google Scholar]
- Alander, M.; Satokari, R.; Korpela, R.; Saxelin, M.; Vilpponen-Salmela, T.; Mattila-Sandholm, T.; von Wright, A. Persistence of colonization of human colonic mucosa by a probiotic strain, Lactobacillus rhamnosus GG, after oral consumption. Appl. Environ. Microbiol. 1999, 65, 351–354. [Google Scholar] [CrossRef] [PubMed]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; de Cadiñanos, L.P.G.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial co-aggregation: An integral process in the development of multi-species biofilms. Trends Microbiol. 2003, 11, 94–100. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Kourelis, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Yiangou, M. Evaluation of adhesion capacity, cell surface traits and immunomodulatory activity of presumptive probiotic Lactobacillus strains. Int. J. Food Microbiol. 2010, 140, 154–163. [Google Scholar] [CrossRef]
- Pan, M.; Kumaree, K.K.; Shah, N.P. Physiological changes of surface membrane in Lactobacillus with prebiotics. J. Funct. Foods 2017, 82, 744–750. [Google Scholar]
- Li, H.; Lu, M.; Guo, H.; Li, W.; Zhang, H. Protective effect of sucrose on the membrane properties of Lactobacillus casei Zhang subjected to freeze-drying. J. Food Prot. 2010, 73, 715–719. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO (Food and Agriculture Organization/World Health Organization). Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; WHO: London, ON, Canada, 2002. [Google Scholar]
- Pan, X.; Wu, T.; Zhang, L.; Cai, L.; Song, Z. Influence of oligosaccharides on the growth and tolerance capacity of lactobacilli to simulated stress environment. Lett. Appl. Microbiol. 2009, 48, 362–367. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S.; Coppola, R. Bile salt and acid tolerance of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. FEMS Microbiol. Lett. 2005, 244, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, R.N.; Iliev, I.N.; Chipeva, V.A.; Dimitonova, S.P.; Samelis, J.; Danova, S.T. Identification and in vitro characterisation of Lactobacillus plantarum strains from artisanal Bulgarian white brined cheeses. J. Basic Microbiol. 2008, 48, 234–244. [Google Scholar] [CrossRef]
- Buriti, F.C.; Castro, I.A.; Saad, S.M. Viability of Lactobacillus acidophilus in synbiotic guava mousses and its survival under in vitro simulated gastrointestinal conditions. Int. J. Food Microbiol. 2010, 137, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Mascaraque, L.G.; Morfin, R.C.; Pérez-Masiá, R.; Sanchez, G.; Lopez-Rubio, A. Optimization of electrospraying conditions for the microencapsulation of probiotics and evaluation of their resistance during storage and in-vitro digestion. LWT-Food Sci. Technol. 2016, 69, 438–446. [Google Scholar] [CrossRef]
- Schell, D.; Beermann, C. Fluidized bed microencapsulation of Lactobacillus reuteri with sweet whey and shellac for improved acid resistance and in-vitro gastrointestinal survival. Food Res. Int. 2014, 62, 308–314. [Google Scholar] [CrossRef]
- Gardiner, G.; Stanton, C.; Lynch, P.B.; Collins, J.K.; Fitzgerald, G.; Ross, R.P. Evaluation of cheddar cheese as a food carrier for delivery of a probiotic strain to the gastrointestinal tract. J. Dairy Sci. 1999, 82, 1379–1387. [Google Scholar] [CrossRef]
- Hernández-Hernández, O.; Muthaiyan, A.; Moreno, F.J.; Montilla, A.; Sanz, M.L.; Ricke, S.C. Effect of prebiotic carbohydrates on the growth and tolerance of Lactobacillus. Food Microbiol. 2012, 30, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.S.; Verruck, S.; Vieira, C.R.; Prudêncio, E.S.; Amante, E.R.; Amboni, R.D. Influence of microencapsulation with sweet whey and prebiotics on the survival of Bifidobacterium-BB-12 under simulated gastrointestinal conditions and heat treatments. LWT-Food Sci. Technol. 2015, 64, 1004–1009. [Google Scholar] [CrossRef]
| Inulin | |||||||
|---|---|---|---|---|---|---|---|
| 0% | 0.5% | 1% | 2% | 3% | 4% | ||
| Negative Control | 0% | Aa 35.5 ± 2.3 | Dc 68.0 ± 0.4 | Dd 72.2 ± 0.4 | Bd 74.0 ± 0.6 | Ac 67.6 ± 0.5 | Ab 61.7 ± 1.0 |
| Glucose | 2% | Fb 84.7 ± 0.7 | GHc 87.8 ± 0.6 | Gd 91.0 ± 0.4 | Fd 91.7 ± 0.7 | Ca 81.7 ± 0.5 | Ca 82.2 ±1.2 |
| 3% | Fb 86.1 ± 0.7 | IJc 90.9 ± 1.2 | Gc 91.9 ± 1.3 | Fc 92.9 ± 0.7 | Db 85.3 ± 1.2 | Ca 81.2 ± 1.1 | |
| 4% | Ge 100.0 ± 0.5 | Jd 93.6 ± 0.9 | Gc 90.9 ± 0.7 | Eb 87.2 ± 0.7 | Db 85.4 ± 0.2 | Ca 82.4 ± 1.1 | |
| Fructose | 2% | Ea 73.9 ± 0.5 | Fbc 81.2 ± 0.5 | Fc 83.5 ± 1.3 | Ed 87.2 ± 0.4 | Cbc 82.0 ± 0.3 | BCb 79.5 ± 1.0 |
| 3% | Fb 85.0 ± 1.8 | GHIbc 88.0 ± 1.4 | Gc 90.7 ± 1.8 | Ebc 88.8 ± 1.0 | BCa 80.3 ± 0.6 | BCa 80.0 ± 0.7 | |
| 4% | Gc 96.3 ± 1.4 | Gb 86.2 ± 0.8 | Fa 82.9 ± 0.7 | Da 82.9 ± 0.1 | Ca 80.9 ± 1.0 | BCa 80.2 ± 1.0 | |
| Sucrose | 2% | Ec 71.1 ± 0.4 | CDa 65.1 ± 0.4 | BCb 68.1 ± 0.3 | Bd 73.3 ± 1.0 | Ce 81.4 ± 0.6 | Ce 82.1 ± 0.4 |
| 3% | Da 62.9 ± 0.6 | Eb 75.9 ± 0.3 | Eb 76.1 ± 0.4 | Dc 82.1 ± 0.3 | Dd 86.1 ± 1.0 | De 93.9 ± 0.5 | |
| 4% | Da 60.3 ± 0.4 | HIe 89.7 ± 0.9 | Fd 86.5 ± 0.8 | Dc 82.1 ± 0.6 | Cc 82.7 ± 0.4 | BCb 79.6 ± 0.4 | |
| Lactose | 2% | Ba 49.1 ± 1.9 | Aa 50.0 ± 0.6 | Ab 64.2 ± 0.2 | Ac 68.6 ± 0.9 | Cd 81.2 ± 0.4 | BCd 80.7 ± 0.7 |
| 3% | Ca 54.5 ± 0.4 | Cb 63.3 ± 1.2 | ABb 65.4 ± 2.1 | Cc 78.7 ± 0.7 | BCc 80.2 ± 0.7 | Cc 80.9 ± 0.2 | |
| 4% | Ca 55.6 ± 0.9 | Ba 58.4 ± 0.8 | CDb 69.3 ± 0.9 | Ab 69.4 ± 1.6 | Bc 78.1 ± 0.9 | Bc 77.8 ± 0.8 | |
| Fructooligosaccharide (FOS) | |||||||
|---|---|---|---|---|---|---|---|
| 0% | 0.5% | 1% | 2% | 3% | 4% | ||
| Negative Control | 0% | Aa 35.7 ± 1.9 | Ab 69.0 ± 0.8 | ABc 75.3 ± 0.2 | Ad 80.0 ± 0.5 | ABd 79.4 ± 0.4 | Ac 73.2 ± 0.1 |
| Glucose | 2% | Fab 83.7 ± 0.1 | EFb 85.2 ± 0.4 | DEc 88.3 ± 1.1 | Fc 90.8 ± 1.1 | Dab 84.1 ± 1.5 | DEa 82.0 ± 0.6 |
| 3% | Gc 86.7 ± 0.4 | FGc 87.2 ± 0.6 | Cb 83.0 ± 0.7 | ABb 83.0 ± 1.6 | Aa 77.9 ± 0.1 | Ba 76.9 ± 0.8 | |
| 4% | Id 100.0 ± 0.5 | Hc 92.5 ± 1.2 | Eb 88.9 ± 1.2 | DEFb 88.3 ± 0.6 | DEa 84.7 ± 0.5 | EFa 83.7 ± 0.9 | |
| Fructose | 2% | Ea 78.3 ± 0.7 | Dab 80.7 ± 0.9 | Cbc 81.4 ± 0.3 | BCDd 85.3 ± 0.9 | DEFd 85.2 ± 0.7 | EFcd 83.3 ± 1.0 |
| 3% | Fb 83.7 ± 0.2 | Gd 93.2 ± 0.2 | Dc 85.8 ± 0.7 | BCb 83.7 ± 0.8 | CDbc 83.7 ± 0.9 | BCDa 79.5 ± 0.6 | |
| 4% | Hd 97.1 ± 0.6 | Hc 92.1 ± 1.3 | Eb 88.6 ± 1.2 | CDEb 86.8 ± 0.8 | BCa 81.3 ± 0.2 | BCa 78.4 ± 0.9 | |
| Sucrose | 2% | Da 65.7 ± 0.5 | Hc 87.7 ± 0.2 | Fd 93.1 ± 0.1 | EFc 89.1 ± 0.9 | FGc 87.3 ± 1.0 | EFb 84.0 ± 0.5 |
| 3% | Da 65.0 ± 0.8 | Ebc 84.3 ± 0.6 | Fd 92.5 ± 0.4 | Ge 95.2 ± 0.1 | DEFGc 86.0 ± 1.1 | EFb 83.5 ± 0.5 | |
| 4% | Ca 61.4 ± 0.6 | EFGc 86.0 ± 0.5 | DEe 88 ± 0.7 | CDEcd 86.4 ± 0.5 | DEc 84.5 ± 0.1 | CDb 80.6 ± 0.9 | |
| Lactose | 2% | Ba 54.9 ± 0.3 | BCb 77.0 ± 0.6 | Bb 77.5 ± 0.4 | Bc 83.1 ± 0.8 | DEFGd 85.4 ± 0.6 | Ge 88.8 ± 0.7 |
| 3% | Ba 53.6 ± 1.1 | CDb 78.4 ± 0.3 | Bb 77.7 ± 0.3 | BCDc 85.7 ± 1.2 | EFGc 86.9 ± 0.1 | Hd 93.1 ± 0.9 | |
| 4% | Ba 53.4 ± 0.2 | Bb 75.2 ± 0.5 | Ab 74.3 ± 0.5 | ABc 82.8 ± 0.6 | Gd 87.9 ±0.8 | Fe 84.7 ± 0.3 | |
| Sucrose | Lactose | Fructose | Lactic Acid | |||||
|---|---|---|---|---|---|---|---|---|
| Week 0 | Week 12 | Week 0 | Week 12 | Week 0 | Week 12 | Week 0 | Week 12 | |
| NC | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 |
| 3S | Ca 1.0 ± 0.0 | Bb 0.0 ± 0.0 | - | - | - | - | Ca 4.3 ± 0.0 | Eb 10.8 ± 0.0 |
| 2L | - | - | Ba 1.4 ± 0.0 | Ab 0.1 ± 0.1 | - | - | Aa 0.0 ± 0.0 | BCb 1.7 ± 0.0 |
| 3L | - | - | Da 2.5 ± 0.0 | Ab 0.2 ± 0.2 | - | - | Ba 1.6 ± 0.1 | Db 4.7 ± 0.0 |
| 3I | - | - | - | - | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | Aa 0.0 ± 0.0 | BCb 1.7 ± 0.0 |
| 4I | - | - | - | - | Aa 0.0 ± 0.0 | Aa 0.1 ± 0.1 | Aa 0.0 ± 0.0 | Cb 2.4 ± 0.2 |
| 2FOS | - | - | - | - | Aa 0.0 ± 0.0 | Aa 0.1 ± 0.1 | Aa 0.0 ± 0.0 | Cb 2.7 ± 0.2 |
| 4FOS | - | - | - | - | Aa 0.1 ± 0.0 | Aa 0.2 ± 0.2 | Aa 0.0 ± 0.0 | Bb 1.3 ± 0.0 |
| 3S+4I | Da 1.4 ± 0.0 | Bb 0.1 ± 0.1 | - | - | BCa 0.2 ± 0.0 | Aa 0.3 ± 0.4 | Ca 4.9 ± 0.1 | Eb 10.8 ± 0.0 |
| 2L+3I | - | - | Ca 1.8 ± 0.0 | Ab 0.3 ± 0.0 | Ca 0.3 ± 0.1 | Aa 0.1 ± 0.0 | Aa 0.2 ± 0.1 | BCb 2.1 ± 0.0 |
| 3S+2FOS | Ba 0.5 ± 0.1 | Bb 0.1 ± 0.0 | - | - | BCa 0.2 ± 0.0 | Aa 0.2 ± 0.2 | Da 6.6 ± 0.0 | Fb 12.5 ± 0.0 |
| 3L+4FOS | - | - | Ea 2.8 ± 0.0 | Bb 0.7 ± 0.1 | Da 0.4 ± 0.1 | Aa 0.1 ± 0.0 | Ba 1.3 ± 0.1 | Db 4.2 ± 0.1 |
| Auto-Aggregation (%) | Hydrophobicity (%) | Co-Aggregation (%) with E. coli | Co-Aggregation (%) with E. faecalis | |||||
|---|---|---|---|---|---|---|---|---|
| Week 0 | Week 12 | Week 0 | Week 12 | Week 0 | Week12 | Week 0 | Week 12 | |
| NC | Aa 46.8 ± 0.1 | Ab 0.0 ± 0.1 | Aa 52.6 ± 0.1 | Ab 0.5 ± 0.1 | Aa 11.6 ± 0.0 | Ab 0.3 ± 0.0 | Aa 16.1 ± 0.0 | Ab 0.8 ± 0.0 |
| 3S | Ba 59.4 ± 0.0 | Bb 66.2 ± 0.0 | Ba 57.7 ± 0.0 | Ab 39.6 ± 0.0 | Ca 34.0 ± 0.1 | Cb 47.4 ± 0.0 | Ba 27.3 ± 0.0 | Cb 54.0 ± 0.7 |
| 2L | Ba 52.3 ± 0.0 | Ab 57.0 ± 0.0 | Ca 68.4 ± 0.0 | Bb 51.0 ± 0.0 | Ba 17.4 ± 0.0 | Bb 34.9 ± 0.0 | Aa 17.8 ± 0.0 | Bb 34.6 ± 0.1 |
| 3L | Ca 63.1 ± 0.1 | Bb 67.2 ± 0.2 | Ca 69.3 ± 0.3 | Bb 54.0 ± 0.0 | Ba 17.6 ± 0.2 | Bb 38.0 ± 0.0 | Aa 17.9 ± 0.0 | Bb 34.6 ± 0.3 |
| 3I | Ca 64.3 ± 0.2 | Ba 63.7 ± 0.2 | Ca 69.5 ± 0.2 | Cb 60.0 ± 0.2 | Da 47.6 ± 0.2 | Cb 51.1 ± 0.0 | Ba 28.5 ± 0.0 | Cb 55.1 ± 0.3 |
| 4I | Ca 65.5 ± 0.1 | Bb 67.3 ± 0.2 | Da 74.5 ± 0.2 | Cb 60.6 ± 0.2 | Ea 52.8 ± 0.2 | CDb 57.6 ± 0.1 | Ba 28.7 ± 0.1 | Cb 55.3 ± 0.6 |
| 2FOS | Ca 67.0 ± 0.2 | Ba 67.9 ± 0.1 | Da 75.9 ± 0.2 | CDb 62.8 ± 0.2 | Ea 57.8 ± 0.2 | Db 61.8 ± 0.1 | BCa 29.6 ± 0.1 | Cb 58.6 ± 0.7 |
| 4FOS | Da 87.4 ± 0.1 | Bb 69.4 ± 0.1 | Da 77.9 ± 0.3 | Db 68.4 ± 0.0 | Ea 57.9 ± 0.2 | Db 62.9 ± 0.0 | Ca 31.3 ± 0.0 | Db 60.7 ± 0.9 |
| 3S+4I | Da 87.5 ± 0.1 | Cb 82.5 ± 0.2 | Fa 89.6 ± 0.1 | Eb 77.6 ± 0.1 | Ga 70.4 ± 0.0 | Eb 76.4 ± 0.0 | Ca 33.6 ± 0.0 | Db 62.5 ± 0.1 |
| 2L+3I | Da 89.6 ± 0.1 | Cb 83.5 ± 0.0 | Fa 88.3 ± 0.2 | Eb 78.1 ± 0.1 | Fa 63.7 ± 0.2 | Gb 89.8 ± 0.0 | Ca 33.8 ± 0.0 | Db 64.3 ± 0.3 |
| 3S+2FOS | Ea 95.0 ± 0.0 | Cb 85.8 ± 0.0 | Fa 90.0 ± 0.0 | Fb 81.1 ± 0.0 | Ea 58.6 ± 0.0 | Eb 78.1 ± 0.0 | Da 38.7 ± 0.0 | Eb 68.1 ± 0.1 |
| 3L+4FOS | Ea 95.1 ± 0.1 | Cb 85.9 ± 0.1 | EFa 86.0 ± 0.1 | Fb 82.4 ± 0.1 | Fa 64.5 ± 0.1 | Fb 80.1 ± 0.0 | Ea 41.8 ± 0.1 | Eb 69.4 ± 0.1 |
| Week 0 | Week 2 | Week 4 | Week 6 | Week 8 | Week 10 | Week 12 | |
|---|---|---|---|---|---|---|---|
| NC | Ac 65.0 ± 1.8 | Ab 55.3 ± 1.1 | Aa 53.7 ± 1.5 | Aa 52.0 ± 1.0 | Aa 51.3 ± 0.6 | Aa 51.0 ± 0.0 | Aa 51.0 ± 0.0 |
| 3S | Cbc 70.0 ± 1.0 | Db 67.2 ± 1.5 | Db 65.7 ± 1.2 | Db 65.2 ± 1.3 | Db 64.7 ± 1.5 | Da 62.7 ± 1.2 | Da 60.3 ± 0.6 |
| 2L | Bd 67.5 ± 0.1 | Cc 63.0 ± 1.7 | Bb 59.3 ± 1.2 | Bb 58.3 ± 1.2 | Db 58.0 ± 1.0 | Ca 56.0 ± 1.0 | Ba 55.3 ± 0.6 |
| 3L | Bd 68.7 ± 0.5 | Dd 67.6 ± 1.5 | Cc 62.0 ± 1.0 | Cb 60.7 ± 1.2 | Dab 59.7 ± 1.2 | Ca 58.3 ± 1.2 | Ca 57.3 ± 0.6 |
| 3I | Ad 65.5 ± 1.5 | Bc 58.7 ± 1.1 | Bc 57.3 ± 0.6 | Bb 55.7 ± 1.5 | Ba 53.3 ± 1.5 | Ba 53.0 ± 1.0 | Aa 52.7 ± 0.6 |
| 4I | Dc 72.7 ± 1.2 | Ec 71.3 ± 1.5 | Dbc 69.7 ± 1.5 | Eb 68.3 ± 1.3 | Db 66.7 ± 1.5 | Da 63.8 ± 2.0 | Da 61.9 ± 0.3 |
| 2FOS | Ed 75.7 ± 0.6 | Fd 75.0 ± 1.7 | Ed 74.0 ± 2.0 | Fc 72.7 ± 1.1 | Ec 72.3 ± 1.3 | Eb 69.3 ± 0.6 | Ea 63.0 ± 1.4 |
| 4FOS | Fd 79.7 ± 1.5 | Gcd 78.3 ± 0.6 | Fcd 78.0 ± 1.0 | Gc 77.3 ± 1.2 | Fc 77.0 ± 1.0 | Fb 72.7 ± 1.2 | Fa 66.6 ± 0.5 |
| 3S+4I | He 91.5 ± 1.1 | Id 86.0 ± 1.6 | Gc 83.7 ± 0.7 | Hc 83.7 ± 0.8 | Gb 80.7 ± 1.5 | Gb 79.0 ± 1.0 | Ga 74.7 ± 0.6 |
| 2L+3I | Hd 90.8 ± 1.1 | Hbc 84.3 ± 1.2 | Gbc 84.4 ± 1.0 | Hb 83.0 ± 1.0 | Gb 82.7 ± 0.6 | Hb 83.7 ± 0.9 | Ha 76.7 ± 0.6 |
| 3S+2FOS | Id 93.7 ± 0.1 | Ic 88.0 ± 1.7 | Hbc 86.7 ± 0.6 | Ibc 86.3 ± 0.6 | Hb 85.3 ± 1.2 | Ibc 87.7 ± 0.6 | Ia 80.3 ± 0.6 |
| 3L+4FOS | Gc 89.0 ± 0.0 | Ic 87.7 ± 1.2 | Ic 88.7 ± 0.6 | Ic 88.7 ± 0.6 | Ic 88.7 ± 0.6 | Ib 86.0 ± 1.5 | Ja 82.6 ± 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parhi, P.; Song, K.P.; Choo, W.S. Viability, Storage Stability and In Vitro Gastrointestinal Tolerance of Lactiplantibacillus plantarum Grown in Model Sugar Systems with Inulin and Fructooligosaccharide Supplementation. Fermentation 2021, 7, 259. https://doi.org/10.3390/fermentation7040259
Parhi P, Song KP, Choo WS. Viability, Storage Stability and In Vitro Gastrointestinal Tolerance of Lactiplantibacillus plantarum Grown in Model Sugar Systems with Inulin and Fructooligosaccharide Supplementation. Fermentation. 2021; 7(4):259. https://doi.org/10.3390/fermentation7040259
Chicago/Turabian StyleParhi, Priyanka, Keang Peng Song, and Wee Sim Choo. 2021. "Viability, Storage Stability and In Vitro Gastrointestinal Tolerance of Lactiplantibacillus plantarum Grown in Model Sugar Systems with Inulin and Fructooligosaccharide Supplementation" Fermentation 7, no. 4: 259. https://doi.org/10.3390/fermentation7040259
APA StyleParhi, P., Song, K. P., & Choo, W. S. (2021). Viability, Storage Stability and In Vitro Gastrointestinal Tolerance of Lactiplantibacillus plantarum Grown in Model Sugar Systems with Inulin and Fructooligosaccharide Supplementation. Fermentation, 7(4), 259. https://doi.org/10.3390/fermentation7040259