
Microbial Composition of Fermented Korean Soy Paste (Doenjang) Prepared by Adding Different Herbs during Fermentation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Soy Paste Samples
2.2. Metagenomic DNA Extraction, Sequencing, and Bioinformatic Analysis
2.3. Statistical Analysis
3. Results
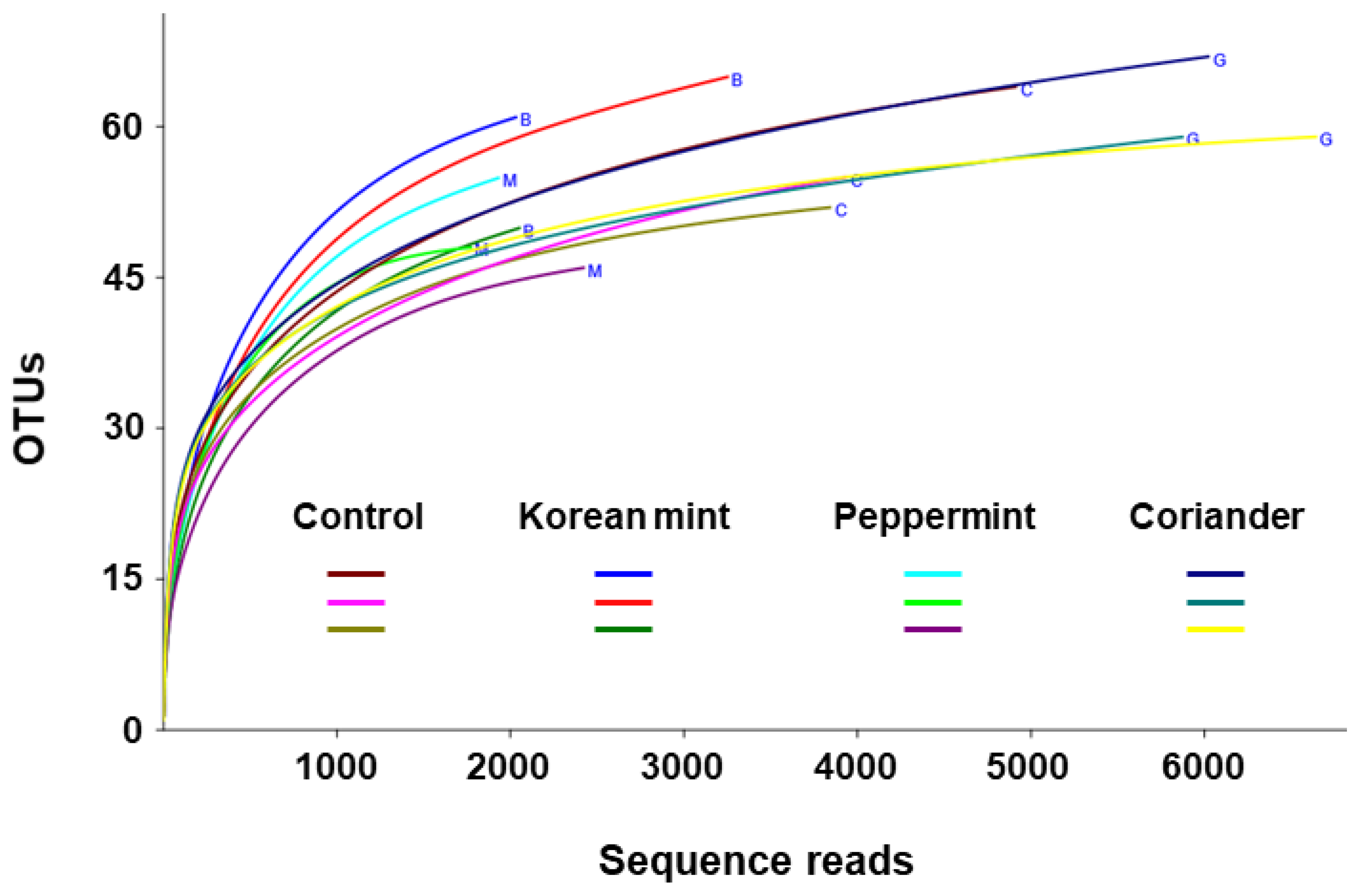
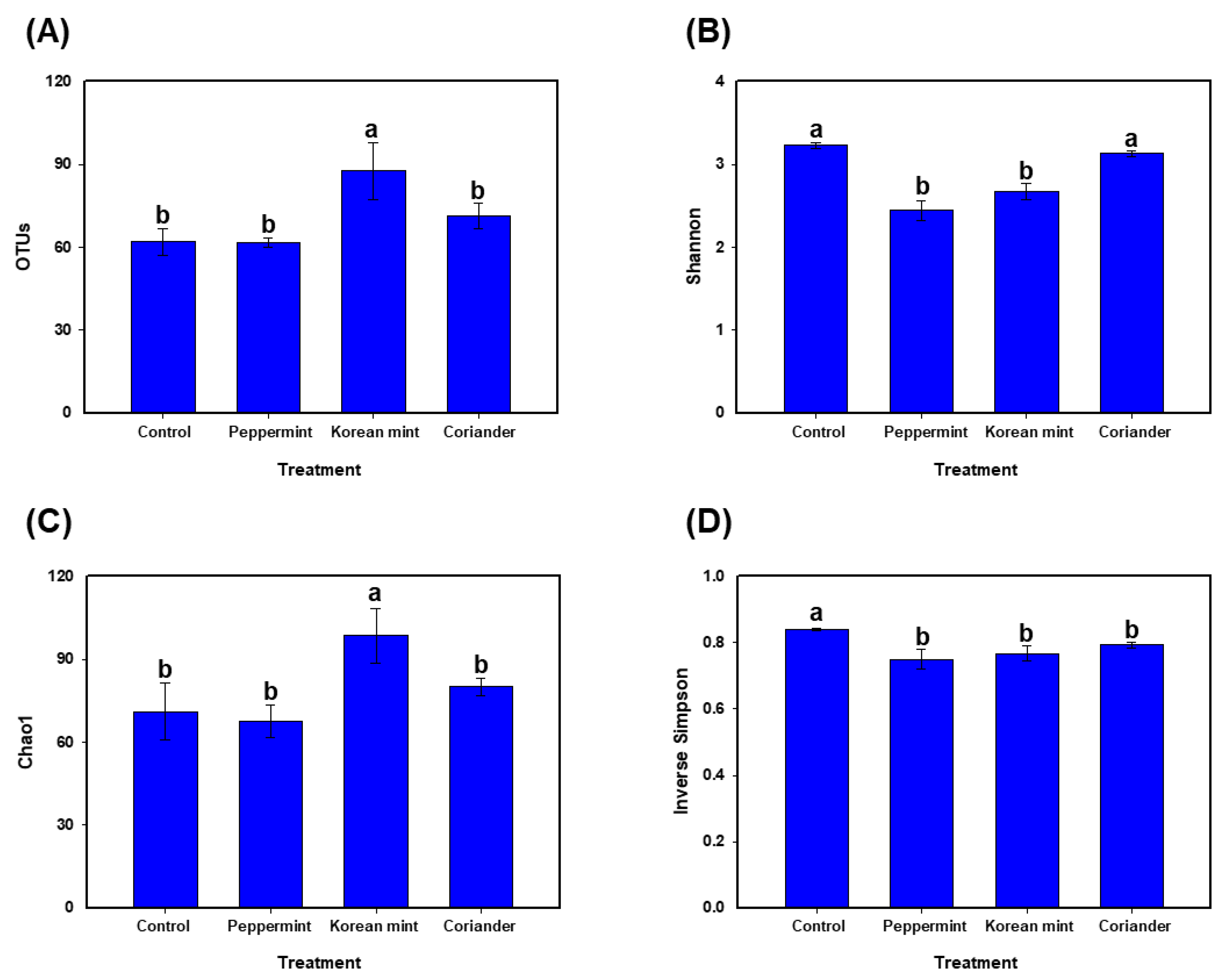
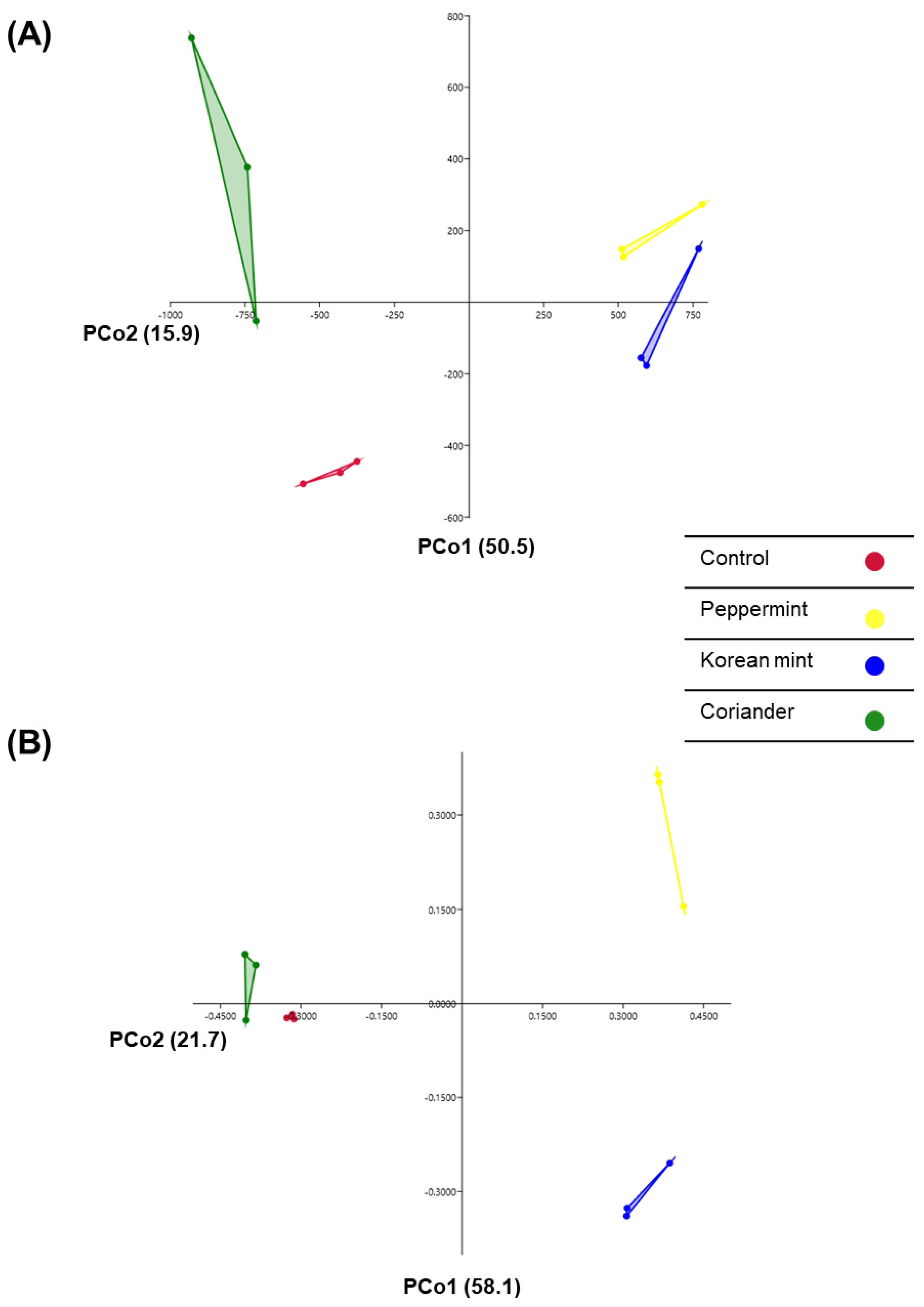
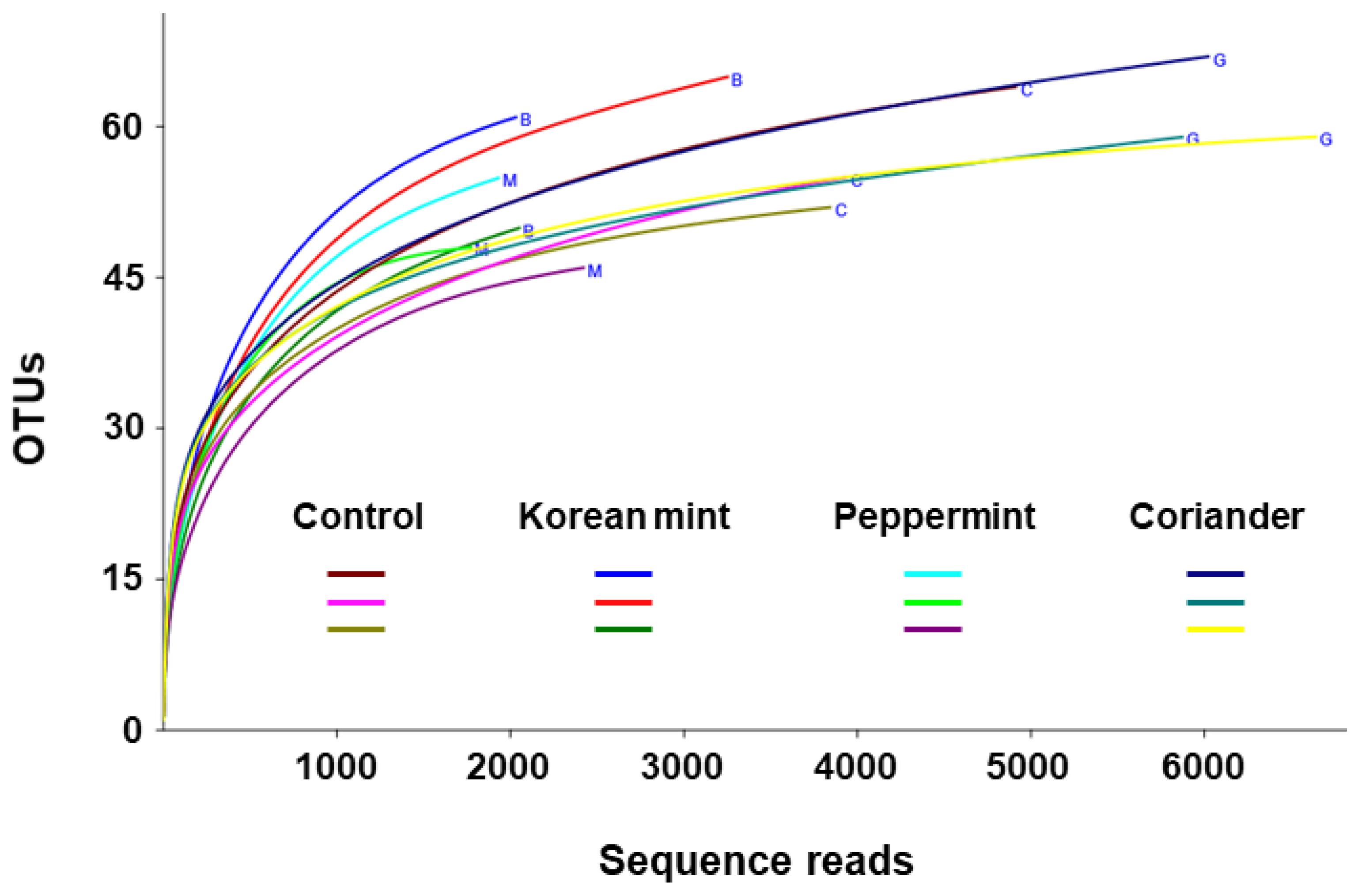
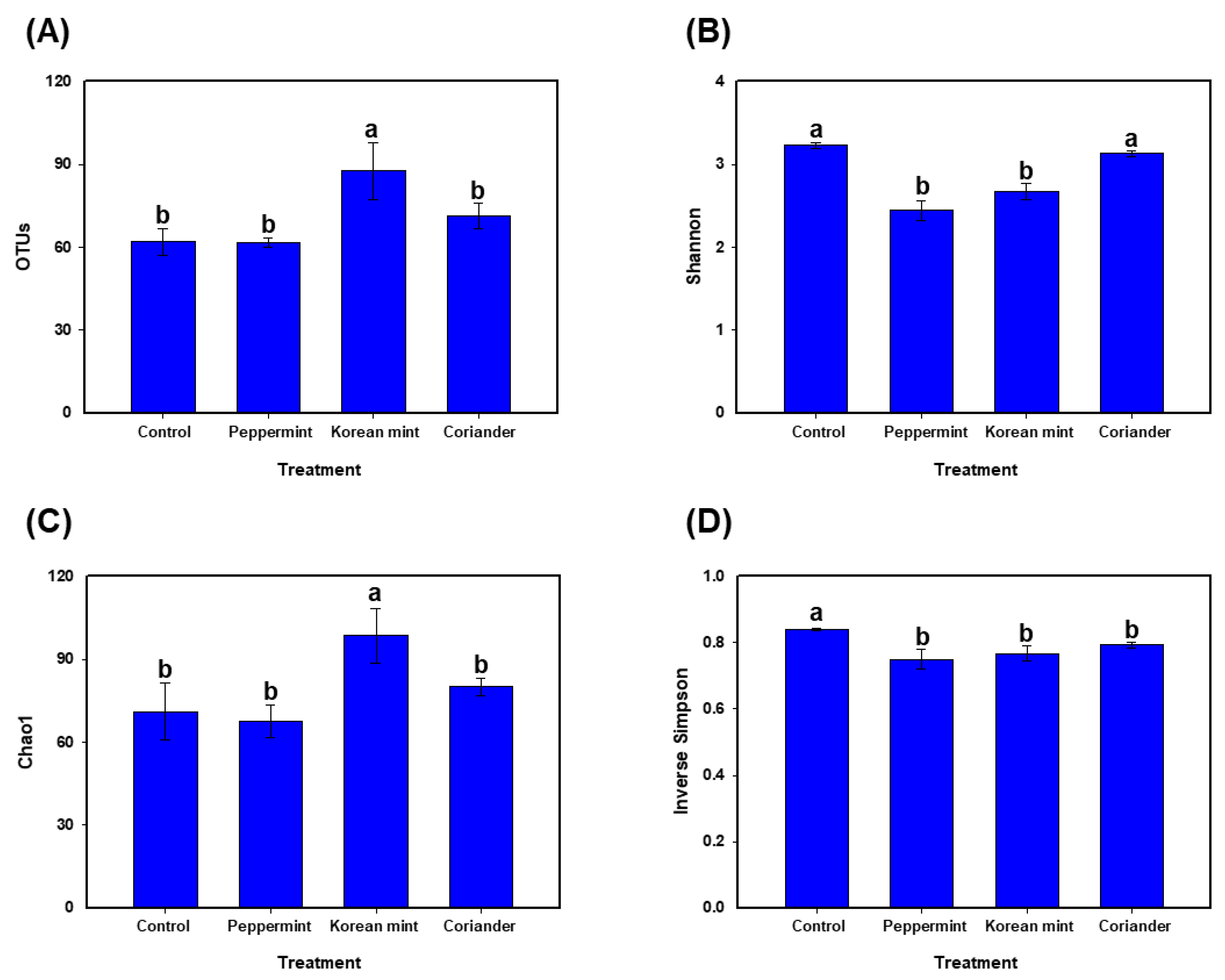
3.1. Diversity of Fermented Soy Paste Microbiota
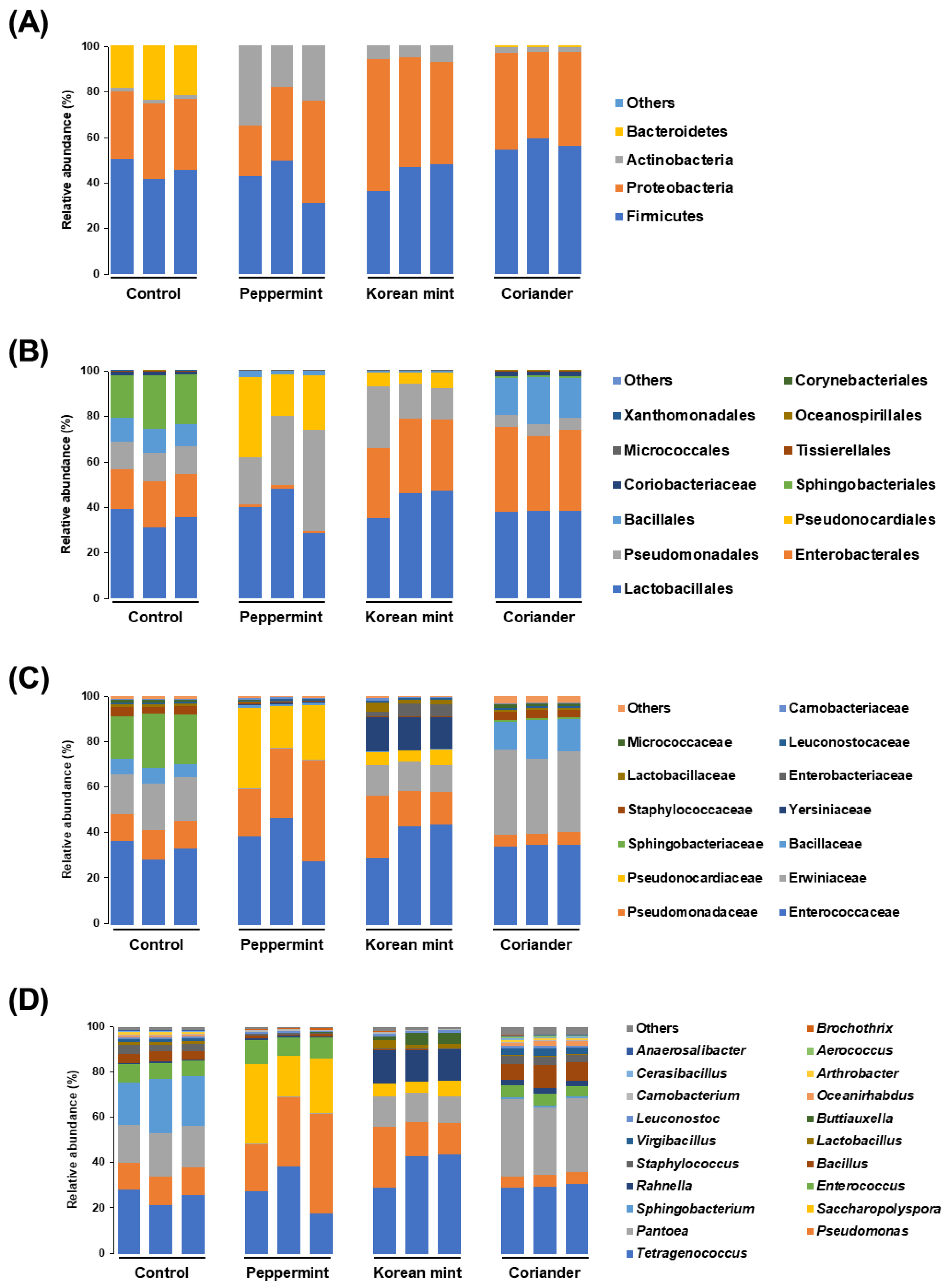
3.2. Effects of Peppermint, Korean Mint, and Coriander on Microbial Composition and Dominant Taxa in Doenjang
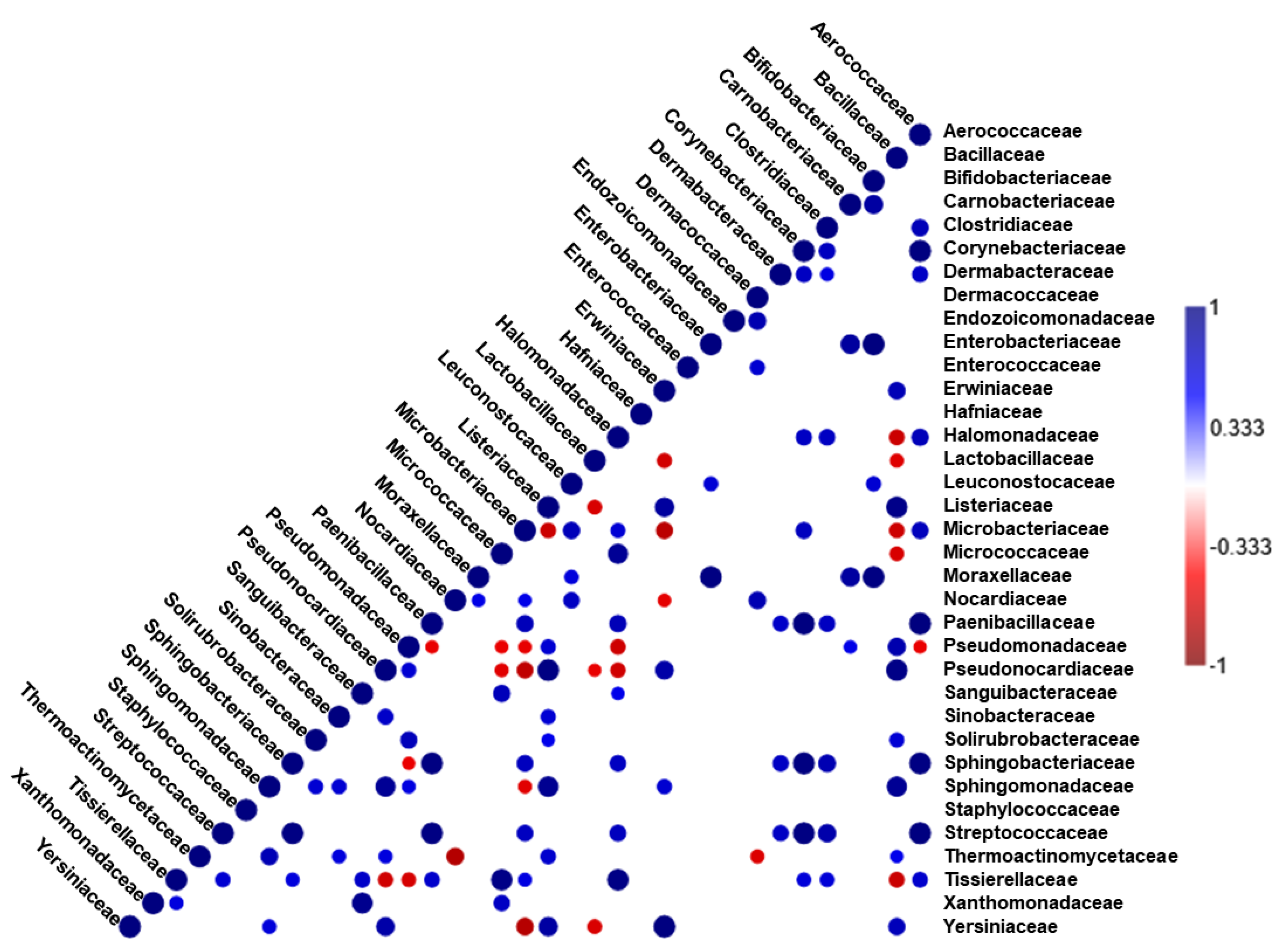
3.3. Correlation Analysis of Relative Abundance of Bacterial Families in Doenjang Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, Z.-H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.-Y. Bioactivity of soy-based fermented foods: A review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef]
- Rolle, R.; Satin, M. Basic requirements for the transfer of fermentation technologies to developing countries. Int. J. Food Microbiol. 2002, 75, 181–187. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, S.; Lee, S.; Oh, J.Y.; Jeon, E.J.; Ryu, H.S.; Lee, C.H. Primary and secondary metabolite profiling of doenjang, a fermented soybean paste during industrial processing. Food Chem. 2014, 165, 157–166. [Google Scholar] [CrossRef]
- Kim, M.; Im, S.; Yoo, Y.; Kim, G.; Lee, J.H. Antioxidative materials in domestic Meju and Doenjang-(4)-Separation of phenolic compounds and their antioxidative activity. J. Korean Soc. Food Sci. Nutr. 1994, 23, 792–798. [Google Scholar]
- Nam, Y.R.; Won, S.B.; Chung, Y.-S.; Kwak, C.S.; Kwon, Y.H. Inhibitory effects ofDoenjang, Korean traditional fermented soybean paste, on oxidative stress and inflammation in adipose tissue of mice fed a high-fat diet. Nutr. Res. Pr. 2015, 9, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Ra, K.; Oh, S.; Kim, J.; Suh, H.J. Isolation of fibrinolytic enzyme and β-glucosidase producing strains from Doenjang and op-timum conditions of enzyme production. J. Korean Soc. Food Sci. Nutr. 2004, 33, 439–442. [Google Scholar]
- Cha, Y.-S.; Yang, J.-A.; Back, H.-I.; Kim, S.-R.; Kim, M.-G.; Jung, S.-J.; O Song, W.; Chae, S.-W. Visceral fat and body weight are reduced in overweight adults by the supplementation ofDoenjang, a fermented soybean paste. Nutr. Res. Pract. 2012, 6, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.-O.; Park, S.-Y.; Park, K.-Y. Longer aging time increases the anticancer and antimetastatic properties of doenjang. Nutrition 2006, 22, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Kwak, C.S.; Park, S.C.; Song, K.Y. Doenjang, a Fermented Soybean Paste, Decreased Visceral Fat Accumulation and Adipocyte Size in Rats Fed with High Fat Diet More Effectively Than Nonfermented Soybeans. J. Med. Food 2012, 15, 1–9. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Daily, J.W., III; Kim, H.J.; Park, S. Antidiabetic effects of fermented soybean products on type 2 diabetes. Nutr. Res. 2010, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Kim, S.H.; Joo, H.K.; Lee, G.S.; Yim, M.H. Isolation and identification of microorganisms producing the soy pro-tein-hydrolyzing enzyme from traditional mejus. Appl. Biol. Chem. 2000, 43, 86–94. [Google Scholar]
- Kwon, D.Y.; Hong, S.M.; Ahn, I.S.; Kim, M.J.; Yang, H.J.; Park, S. Isoflavonoids and peptides from meju, long-term fermented soybeans, increase insulin sensitivity and exert insulinotropic effects in vitro. Nutrition 2011, 27, 244–252. [Google Scholar] [CrossRef]
- Jung, W.Y.; Jung, J.Y.; Lee, H.J.; Jeon, C.O. Functional Characterization of Bacterial Communities Responsible for Fermentation of Doenjang: A Traditional Korean Fermented Soybean Paste. Front. Microbiol. 2016, 7, 827. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Song, K.-M.; Kim, H.-J.; Kim, B.-S.; Rhee, Y.K.; Lee, M.-K. Characterization and use of microbial communities in Doenjang to control the unpleasant odor of Ginkgo epicarp. Food Sci. Biotechnol. 2014, 23, 1959–1964. [Google Scholar] [CrossRef]
- Park, H.-K.; Gil, B.; Kim, J.K. Characteristics of taste components of commercial soybean paste. Food Sci. Biotechnol. 2002, 11, 376–379. [Google Scholar]
- Lee, S.S.; Park, K.H.; Choi, K.J.; Won, S. Identification and isolation of zygomycetous fungi found on maeju, a raw material of Korean traditional soysources. Korean J. Mycol. 1993, 21, 172–187. [Google Scholar]
- Yoo, S.; Cho, W.; Kang, S.; Lee, S.H. Isolation and identification of microorganisms in Korean traditional soybena paste and soybean sauce. Korean J. Microbiol. Biotechnol. 1999, 27, 113–117. [Google Scholar]
- Jeong, D.-W.; Kim, H.-R.; Jung, G.; Han, S.; Kim, C.-T.; Lee, J.-H. Bacterial community migration in the ripening of doenjang, a traditional Korean fermented soybean food. J. Microbiol. Biotechnol. 2014, 24, 648–660. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.M.; Seo, W.T. Bacterial diversity in a Korean traditional soybean fermented foods (doenjang and ganjang) by 16S rRNA gene sequence analysis. Food Sci. Biotechnol. 2007, 16, 320–324. [Google Scholar]
- Kim, T.-W.; Lee, J.-H.; Kim, S.-E.; Park, M.-H.; Chang, H.C.; Kim, H.-Y. Analysis of microbial communities in doenjang, a Korean fermented soybean paste, using nested PCR-denaturing gradient gel electrophoresis. Int. J. Food Microbiol. 2009, 131, 265–271. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Microbial community dynamics during fermentation of doenjang-meju, traditional Korean fermented soybean. Int. J. Food Microbiol. 2014, 185, 112–120. [Google Scholar] [CrossRef]
- Kim, M.J.; Kwak, H.S.; Jung, H.Y.; Kim, S.S. Microbial communities related to sensory attributes in Korean fermented soy bean paste (doenjang). Food Res. Int. 2016, 89, 724–732. [Google Scholar] [CrossRef]
- Song, D.; Chun, B.; Lee, S.; Son, S.; Reddy, C.; Mun, H.; Jeon, C.; Lee, C. Comprehensive Metabolite Profiling and Microbial Communities of Doenjang (Fermented Soy Paste) and Ganjang (Fermented Soy Sauce): A Comparative Study. Foods 2021, 10, 641. [Google Scholar] [CrossRef]
- Tapsell, L.C.; Hemphill, I.; Cobiac, L.; Sullivan, D.R.; Fenech, M.; Patch, C.S.; Roodenrys, S.; Keogh, J.B.; Clifton, P.; Williams, P.G.; et al. Health benefits of herbs and spices: The past, the present, the future. Med. J. Aust. 2006, 185, S1–S24. [Google Scholar] [CrossRef] [Green Version]
- Areias, F.; Valentão, P.; Andrade, P.; Ferreres, F.; Seabra, R. Phenolic fingerprint of peppermint leaves. Food Chem. 2001, 73, 307–311. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Baskar, T.B.; Park, Y.E.; Park, J.S.; Lee, S.Y.; Park, S.U. In Vitro Antioxidant and Antimicrobial Properties of Flower, Leaf, and Stem Extracts of Korean Mint. Antioxidants 2019, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Wangensteen, H.; Samuelsen, A.B.; Malterud, K.E. Antioxidant activity in extracts from coriander. Food Chem. 2004, 88, 293–297. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2011, 6, 610–618. [Google Scholar] [CrossRef]
- Li, W.; Chang, Y. CD-HIT-OTU-MiSeq, an improved approach for clustering and analyzing paired end MiSeq 16S rRNA sequences. BioRxiv 2017, 153783. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Hammer, O.; Harper, D.A.; Ryan, P.D. Palaeontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Bahuguna, A.; Shukla, S.; Lee, J.S.; Bajpai, V.K.; Kim, S.-Y.; Huh, Y.S.; Han, Y.-K.; Kim, M. Garlic augments the functional and nutritional behavior of Doenjang, a traditional Korean fermented soybean paste. Sci. Rep. 2019, 9, 5436. [Google Scholar] [CrossRef]
- Hwang, K.-M.; Jung, K.-O.; Song, C.-H.; Park, K.-Y. Increased Antimutagenic and Anticlastogenic Effects of Doenjang (Korean Fermented Soybean Paste) Prepared with Bamboo Salt. J. Med. Food 2008, 11, 717–722. [Google Scholar] [CrossRef]
- Shim, J.M.; Lee, K.W.; Yao, Z.; Kim, H.-J.; Kim, J.H. Properties of Doenjang (Soybean Paste) Prepared with Different Types of Salts. J. Microbiol. Biotechnol. 2016, 26, 1533–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.; Lee, J.S.; Bajpai, V.K.; Khan, I.; Huh, Y.S.; Han, Y.-K.; Kim, M. Toxicological evaluation of lotus, ginkgo, and garlic tailored fermented Korean soybean paste (Doenjang) for biogenic amines, aflatoxins, and microbial hazards. Food Chem. Toxicol. 2019, 133, 110729. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.K.; Kang, S.M.; Noh, Y.S. Quality properties on soy bean pastes made with microorganisms isolated from traditional soy bean pastes. Korean J. Microbiol. Biotechnol. 2000, 32, 1266–1270. [Google Scholar]
- Masuda, S.; Yamaguchi, H.; Kurokawa, T.; Shirakami, T.; Tsuji, R.; Nishimura, I. Immunomodulatory effect of halophilic lactic acid bacterium Tetragenococcus halophilus Th221 from soy sauce moromi grown in high-salt medium. Int. J. Food Microbiol. 2008, 121, 245–252. [Google Scholar] [CrossRef]
- Nishimura, I.; Igarashi, T.; Enomoto, T.; Dake, Y.; Okuno, Y.; Obata, A. Clinical Efficacy of Halophilic Lactic Acid Bacterium Tetragenococcus halophilus Th221 from Soy Sauce Moromi for Perennial Allergic Rhinitis. Allergol. Int. 2009, 58, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Huang, J.; Jin, Y.; Wu, C.; Shen, D.; Zhang, S.; Zhou, R. Aflatoxin B1 degradation by salt tolerant Tetragenococcus halophilus CGMCC 3792. Food Chem. Toxicol. 2018, 121, 430–436. [Google Scholar] [CrossRef]
- Manivasagan, P.; Kang, K.-H.; Sivakumar, K.; Li-Chan, E.C.; Oh, H.-M.; Kim, S.-K. Marine actinobacteria: An important source of bioactive natural products. Environ. Toxicol. Pharmacol. 2014, 38, 172–188. [Google Scholar] [CrossRef]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Marine actinobacterial metabolites and their pharmaceutical po-tential. In Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1371–1386. [Google Scholar]
- Nie, Z.; Zheng, Y.; Xie, S.; Zhang, X.; Song, J.; Xia, M.; Wang, M. Unraveling the correlation between microbiota succession and metabolite changes in traditional Shanxi aged vinegar. Sci. Rep. 2017, 7, 9240. [Google Scholar] [CrossRef] [Green Version]
- Marzban, A.; Ebrahimipour, G.; Danesh, A. Bioactivity of a Novel Glycolipid Produced by a Halophilic Buttiauxella sp. and Improving Submerged Fermentation Using a Response Surface Method. Molecules 2016, 21, 1256. [Google Scholar] [CrossRef] [Green Version]
- Lambiase, A.; Rossano, F.; Del Pezzo, M.; Raia, V.; Sepe, A.; De Gregorio, F.; Catania, M.R. Sphingobacterium respiratory tract infection in patients with cystic fibrosis. BMC Res. Notes 2009, 2, 262. [Google Scholar] [CrossRef] [Green Version]
- Tronel, H.; Plesiat, P.; Ageron, E.; Grimont, P. Bacteremia caused by a novel species of Sphingobacterium. Clin. Microbiol. Infect. 2003, 9, 1242–1244. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, D.J.; Leber, A. Enterobacter, Cronobacter, and Pantoea Species. In Principles and Practice of Pediatric Infectious Diseases; Elsevier BV: Amsterdam, The Netherlands, 2018; pp. 824–827.e1. [Google Scholar]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannaa, M.; Cho, S.-S.; Seo, Y.-S.; Park, I. Microbial Composition of Fermented Korean Soy Paste (Doenjang) Prepared by Adding Different Herbs during Fermentation. Fermentation 2021, 7, 93. https://doi.org/10.3390/fermentation7020093
Mannaa M, Cho S-S, Seo Y-S, Park I. Microbial Composition of Fermented Korean Soy Paste (Doenjang) Prepared by Adding Different Herbs during Fermentation. Fermentation. 2021; 7(2):93. https://doi.org/10.3390/fermentation7020093
Chicago/Turabian StyleMannaa, Mohamed, Seong-Soon Cho, Young-Su Seo, and Inmyoung Park. 2021. "Microbial Composition of Fermented Korean Soy Paste (Doenjang) Prepared by Adding Different Herbs during Fermentation" Fermentation 7, no. 2: 93. https://doi.org/10.3390/fermentation7020093
APA StyleMannaa, M., Cho, S.-S., Seo, Y.-S., & Park, I. (2021). Microbial Composition of Fermented Korean Soy Paste (Doenjang) Prepared by Adding Different Herbs during Fermentation. Fermentation, 7(2), 93. https://doi.org/10.3390/fermentation7020093