Brewer’s Spent Yeast (BSY), an Underutilized Brewing By-Product



Abstract
1. Introduction
2. Use of Yeast in Brewing and Fermentation—An Overview
2.1. Yeast
2.2. A Brief Overview of Fermentation
2.3. Pitching
2.4. Yeast Viability
3. Formation and Processing of Brewer’s Spent Yeast (BSY)
3.1. Flocculation
3.2. Production of Yeast Extract
3.3. Modification of Yeast Extract: Debittering
4. BSY Composition and Properties
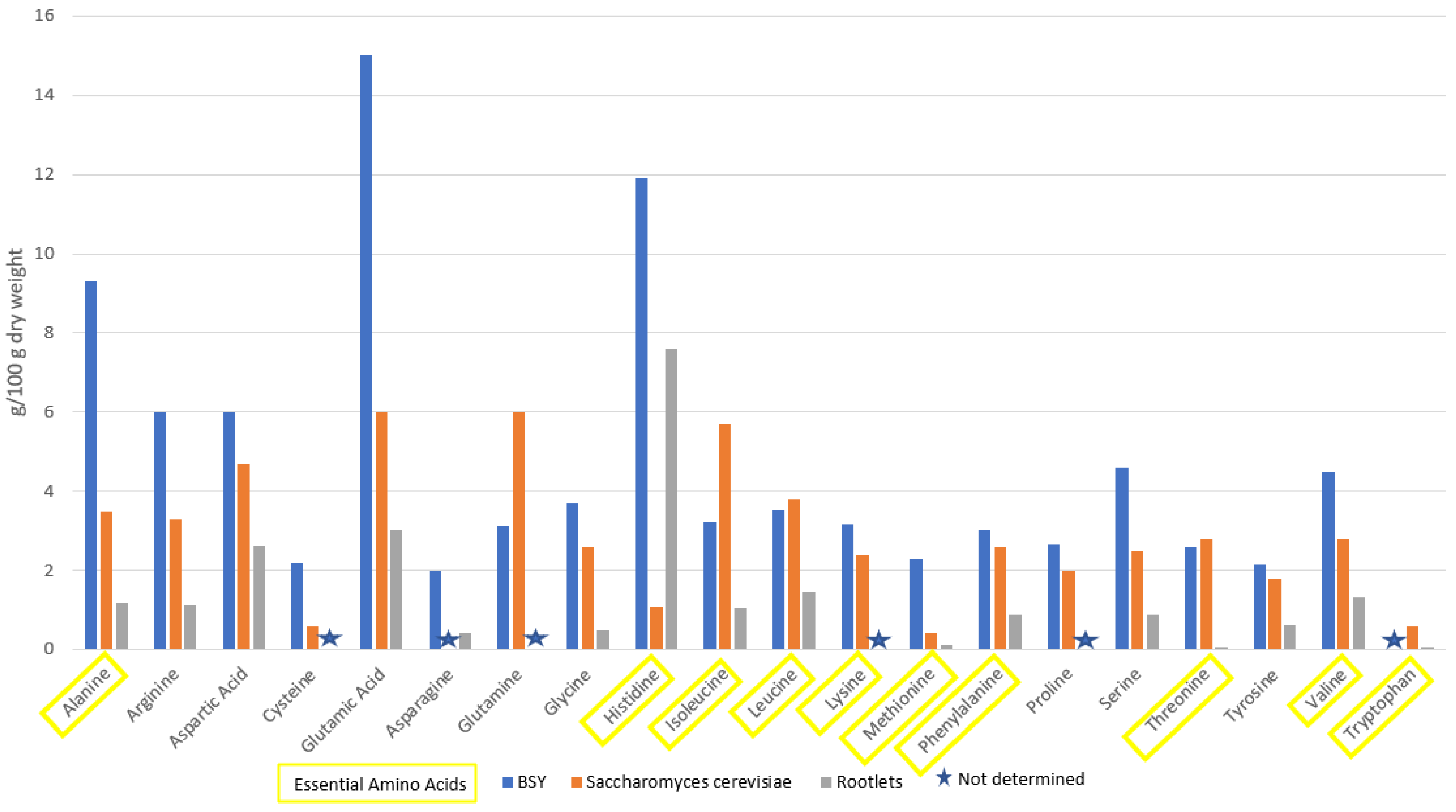
4.1. Protein
4.2. Carbohydrates
4.3. Mineral and Vitamin Content
4.4. Phenolic Compounds
4.5. Glutathione
5. BSY Applications
5.1. Animal Feed
5.2. Brewer’s Spent Yeast as an Enzyme Source
5.3. Functional Food Applications
5.3.1. β-Glucans and Mannoproteins as Functional Food Additives
5.3.2. Reducing Agents
5.3.3. Health and Nutritional Applications
Β-Glucans
Encapsulating Agent
Cryoprotectant for Probiotic Bacteria
Antioxidant Properties
5.3.4. Flavor-Enhancing Ingredient
5.3.5. The RNA Issue—Purines
5.3.6. Meat Substitute Applications
5.4. Fermentation Substrate
5.5. Biofuels—Methane Production
5.6. Biosorption Agent
5.7. Supplementation of Ethanol Fermentation
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kunze, W. Technology Brewing and Malting, 2nd ed.; VLB Berlin: Berlin, Germany, 1999. [Google Scholar]
- Podpora, B.; Świderski, F.; Sadowska, A.; Rakowska, R.; Wasiak-Zys, G. Spent brewer’s yeast extracts as a new component of functional food. Czech J. Food Sci. 2016, 34, 554–563. [Google Scholar] [CrossRef]
- Edozien, J.C.; Udo, U.U.; Young, V.R.; Scrimshaw, N.S. Effects of High Levels of Yeast Feeding on Uric Acid Metabolism of Young Men. Nat. Cell Biol. 1970, 228, 180. [Google Scholar] [CrossRef] [PubMed]
- Araújo, V.B.D.S.; De Melo, A.N.F.; Costa, A.G.; Castro-Gomez, R.H.; Madruga, M.S.; De Souza, E.L.; Magnani, M. Followed extraction of β-glucan and mannoprotein from spent brewer’s yeast (Saccharomyces uvarum) and application of the obtained mannoprotein as a stabilizer in mayonnaise. Innov. Food Sci. Emerg. Technol. 2014, 23, 164–170. [Google Scholar] [CrossRef]
- Pinto, M.; Coelho, E.; Nunes, A.; Brandão, T.; Coimbra, M.A. Valuation of brewers spent yeast polysaccharides: A structural characterization approach. Carbohydr. Polym. 2015, 116, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Pinho, O.; Ferreira, I.; Santos, L.H. Method optimization by solid-phase microextraction in combination with gas chromatography with mass spectrometry for analysis of beer volatile fraction. J. Chromatogr. A 2006, 1121, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing—A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Verachtert, H.; Delvaux, F.R. Yeast flocculation: What brewers should know. Appl. Microbiol. Biotechnol. 2003, 61, 197–205. [Google Scholar] [CrossRef]
- Ferreira, I.; Pinho, O.; Vieira, E.; Tavarela, J. Brewer’s Saccharomyces yeast biomass: Characteristics and potential applications. Trends Food Sci. Technol. 2010, 21, 77–84. [Google Scholar] [CrossRef]
- Bamforth, C. Beer: Tap into the Art and Science of Brewing, 1st ed.; Insight Books: New York, NY, USA, 1998. [Google Scholar]
- Lewis, M.J.J.; Young, T.W. Brewing, 1st ed.; Chapman & Hall: London, UK, 1995. [Google Scholar]
- Wainwright, T. Diacetyl-a review: Part i-analytical and biochemical considerations: Part ii-brewing experience. J. Inst. Brew. 1973, 79, 451–470. [Google Scholar] [CrossRef]
- Verbelen, P.J.; Dekoninck, T.M.L.; Saerens, S.M.G.; Van Mulders, S.E.; Thevelein, J.M.; Delvaux, F.R. Impact of pitching rate on yeast fermentation performance and beer flavour. Appl. Microbiol. Biotechnol. 2009, 82, 155–167. [Google Scholar] [CrossRef]
- Kucharczyk, K.; Tuszyński, T. The effect of pitching rate on fermentation, maturation and flavour compounds of beer produced on an industrial scale. J. Inst. Brew. 2015, 121, 349–355. [Google Scholar] [CrossRef]
- Vieira, E.; Moura, C.; Almeida, T.; Meireles, S.; Brandão, T.; Pinho, O.; Ferreira, I. Influence of Serial Repitching on Beer Polypeptide Profiles. J. Am. Soc. Brew. Chem. 2012, 70, 275–279. [Google Scholar] [CrossRef]
- Bouix, M.; Leveau, J.-Y. Rapid Assessment of Yeast Viability and Yeast Vitality During Alcoholic Fermentation. J. Inst. Brew. 2001, 107, 217–225. [Google Scholar] [CrossRef]
- Painting, K.; Kirsop, B. A quick method for estimating the percentage of viable cells in a yeast population, using methylene blue staining. World J. Microbiol. Biotechnol. 1990, 6, 346–347. [Google Scholar] [CrossRef] [PubMed]
- Smart, K.A.; Chambers, K.M.; Lambert, I.; Jenkins, C.; Smart, C.A. Use of Methylene Violet Staining Procedures to Determine Yeast Viability and Vitality. J. Am. Soc. Brew. Chem. 1999, 57, 18–23. [Google Scholar] [CrossRef]
- Straver, M.H.; Aar, P.C.V.D.; Smit, G.; Kijne, J.W. Determinants of flocculence of brewer’s yeast during fermentation in wort. Yeast 1993, 9, 527–532. [Google Scholar] [CrossRef]
- Vieira, E.F.; Carvalho, J.; Pinto, E.; Cunha, S.; Almeida, A.A.; Ferreira, I. Nutritive value, antioxidant activity and phenolic compounds profile of brewer’s spent yeast extract. J. Food Compos. Anal. 2016, 52, 44–51. [Google Scholar] [CrossRef]
- Babayan, T.L.; Bezrukov, M.G. Autolysis in yeasts. Acta Biotechnol. 1985, 5, 129–136. [Google Scholar] [CrossRef]
- Podpora, B.; Swiderski, F.; Sadowska, A.; Piotrowska, A.; Rakowska, R. Spent Brewer’s Yeast Autolysates as a New and Valuable Component of Functional Food and Dietary Supplements. J. Food Process. Technol. 2015, 6, 6. [Google Scholar] [CrossRef]
- Tanguler, H.; Erten, H. Utilisation of spent brewer’s yeast for yeast extract production by autolysis: The effect of temperature. Food Bioprod. Process. 2008, 86, 317–321. [Google Scholar] [CrossRef]
- Bayarjargal, M.; Munkhbat, E.; Ariunsaikhan, T.; Odonchimeg, M.; Uurzaikh, T.; Gan-Erdene, T.; Regdel, D. Utilization of spent brewer’s yeast Saccharomyces cerevisiae for the production of yeast enzymatic hydrolysate. Mong. J. Chem. 2014, 12, 88–91. [Google Scholar] [CrossRef]
- Jacob, F.F.; Hutzler, M.; Methner, F.-J. Comparison of various industrially applicable disruption methods to produce yeast extract using spent yeast from top-fermenting beer production: Influence on amino acid and protein content. Eur. Food Res. Technol. 2019, 245, 95–109. [Google Scholar] [CrossRef]
- Amorim, M.; Pereira, J.O.; Gomes, D.; Pereira, C.D.; Pinheiro, H.; Pintado, M.M.E. Nutritional ingredients from spent brewer’s yeast obtained by hydrolysis and selective membrane filtration integrated in a pilot process. J. Food Eng. 2016, 185, 42–47. [Google Scholar] [CrossRef]
- Dwivedi, B.; Gibson, D. Processing of Spent Brewers’ Yeast for Food Use. Can. Inst. Food Technol. J. 1970, 3, 110–112. [Google Scholar] [CrossRef]
- Shotipruk, A.; Kittianong, P.; Suphantharika, M.; Muangnapoh, C. Application of rotary microfiltration in debittering process of spent brewer’s yeast. Bioresour. Technol. 2005, 96, 1851–1859. [Google Scholar] [CrossRef]
- Nand, K. Debittering of spent brewer’s yeast for food purposes. Food/Nahrung 1987, 31, 127–131. [Google Scholar] [CrossRef]
- FAO/WHO. Report of a joint FAO/WHO/UNU Expert Consultation technical report FAO/WHO and United Nations University. In Energy and Protein Requirements; World Health Organization: Geneva, Switzerland, 1985. [Google Scholar]
- Jacob, F.F.; Striegel, L.; Rychlik, M.; Hutzler, M.; Methner, F.-J. Yeast extract production using spent yeast from beer manufacture: Influence of industrially applicable disruption methods on selected substance groups with biotechnological relevance. Eur. Food Res. Technol. 2019, 245, 1169–1182. [Google Scholar] [CrossRef]
- Fsai Commission Directive. 2008/100/ECof 28 October 2008 amending Council Directive 90/496/EEC on nutrition labelling for foodstuffs as regards recommended daily allowances, energy conversion factors and definitions. Off. J. Eur. Union 2008, 51, 1–4. [Google Scholar]
- Waters, D.M.; Kingston, W.; Jacob, F.; Titze, J.; Arendt, E.K.; Zannini, E. Wheat bread biofortification with rootlets, a malting by-product. J. Sci. Food Agric. 2013, 93, 2372–2383. [Google Scholar] [CrossRef]
- Martini, A.E.V.; Miller, M.W.; Martini, A. Amino acid composition of whole cells of different yeasts. J. Agric. Food Chem. 1979, 27, 982–984. [Google Scholar] [CrossRef]
- Lipke, P.N.; Ovalle, R. Cell Wall Architecture in Yeast: New Structure and New Challenges. J. Bacteriol. 1998, 180, 3735–3740. [Google Scholar] [CrossRef] [PubMed]
- Manners, D.J.; Masson, A.J.; Patterson, J.C. The structure of a β-(1 to 3)-d-glucan from yeast cell walls. Biochem. J. 1973, 135, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A critical review on the impacts of β-glucans on gut microbiota and human health. J. Nutr. Biochem. 2018, 61, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Worrasinchai, S.; Suphantharika, M.; Pinjai, S.; Jamnong, P. β-Glucan prepared from spent brewer’s yeast as a fat replacer in mayonnaise. Food Hydrocoll. 2006, 20, 68–78. [Google Scholar] [CrossRef]
- Thammakiti, S.; Suphantharika, M.; Phaesuwan, T.; Verduyn, C. Preparation of spent brewer’s yeast beta-glucans for potential applications in the food industry. Int. J. Food Sci. Technol. 2003, 39, 21–29. [Google Scholar] [CrossRef]
- Arvindekar, A.U.; Patil, N.B. Glycogen—A covalently linked component of the cell wall in Saccharomyces cerevisiae. Yeast 2002, 19, 131–139. [Google Scholar] [CrossRef]
- Cameron, D.R.; Cooper, D.G.; Neufeld, R.J. The mannoprotein of Saccharomyces cerevisiae is an effective bioemulsifier. Appl. Environ. Microbiol. 1988, 54, 1420–1425. [Google Scholar] [CrossRef]
- Pinto, L.; Lopes, M.; Carvalho Filho, C.; Alves, L.; Benevides, C. Determinação do valor nutritivo de derivados de levedura de cervejaria (Saccharomyces spp.). Rev. Bras. Prod. Agroind. 2013, 15, 7–17. [Google Scholar] [CrossRef]
- León-González, M.E.; Gómez-Mejía, E.; Rosales-Conrado, N.; Madrid-Albarrán, Y. Residual brewing yeast as a source of polyphenols: Extraction, identification and quantification by chromatographic and chemometric tools. Food Chem. 2018, 267, 246–254. [Google Scholar] [CrossRef]
- Alanazi, A.M.; Mostafa, G.A.E.; Al-Badr, A.A. Profiles of Drug Substances, Excipients and Related Methodology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 40, pp. 43–158. [Google Scholar]
- Shimizu, H.; Araki, K.; Shioya, S.; Suga, K.-I. Optimal production of glutathione by controlling the specific growth rate of yeast in fed-batch culture. Biotechnol. Bioeng. 1991, 38, 196–205. [Google Scholar] [CrossRef]
- Liu, C.-H.; Hwang, C.-F.; Liao, C.-C. Medium optimization for glutathione production by Saccharomyces cerevisiae. Process. Biochem. 1999, 34, 17–23. [Google Scholar] [CrossRef]
- Wohlt, J.; Corcione, T.; Zajac, P. Effect of Yeast on Feed Intake and Performance of Cows Fed Diets Based on Corn Silage during Early Lactation. J. Dairy Sci. 1998, 81, 1345–1352. [Google Scholar] [CrossRef]
- Vohra, A.; Syal, P.; Madan, A. Probiotic yeasts in livestock sector. Anim. Feed Sci. Technol. 2016, 219, 31–47. [Google Scholar] [CrossRef]
- Harlow, B.; Bryant, R.; Cohen, S.; O’Connell, S.; Flythe, M.D. Degradation of spent craft brewer’s yeast by caprine rumen hyper ammonia-producing bacteria. Lett. Appl. Microbiol. 2016, 63, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.L.; Oliveira, R.J.F.; Bezerra, L.; Nascimento, T.; de Pellegrini, C.B.; de Freitas Neto, M.D.; do Nascimento, N.G., Jr.; de Souza, W.F. Substitution of corn meal with dry brewer´s yeast in the diet of sheep. Rev. Colomb. Cienc. Pecu. 2016, 29, 99–107. [Google Scholar] [CrossRef]
- Line, J.E.; Bailey, J.S.; Cox, N.A.; Stern, N.J.; Tompkins, T. Effect of yeast-supplemented feed on Salmonella and Campylobacter populations in broilers. Poult. Sci. 1998, 77, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Milewski, S.; Wójcik, R.; Zaleska, B.; Małaczewska, J.; Tański, Z.; Siwicki, A.K. Effect of Saccharomyces cerevisiae dried yeast on the meat performance traits and selected indicators of humoral immunity in lambs. Acta Vet. Brno 2013, 82, 147–151. [Google Scholar] [CrossRef][Green Version]
- Martin, D.S.; Orive, M.; Iñarra, B.; Castelo, J.; Estévez, A.; Nazzaro, J.; Iloro, I.; Elortza, F.; Zufía, J. Brewers’ Spent Yeast and Grain Protein Hydrolysates as Second-Generation Feedstuff for Aquaculture Feed. Waste Biomass Valoriz. 2020, 11, 5307–5320. [Google Scholar] [CrossRef]
- Nguyen, N.H.Y.; Trinh, L.T.; Chau, D.T.; Baruah, K.; Lundh, T.; Kiessling, A. Spent brewer’s yeast as a replacement for fishmeal in diets for giant freshwater prawn (Macrobrachium rosenbergii), reared in either clear water or a biofloc environment. Aquac. Nutr. 2019, 25, 970–979. [Google Scholar] [CrossRef]
- Castro, C.; Pérez-Jiménez, A.; Coutinho, F.; Pousão-Ferreira, P.; Brandão, T.M.; Oliva-Teles, A.; Peres, H. Digestive enzymes of meagre (Argyrosomus regius) and white seabream (Diplodus sargus). Effects of dietary brewer’s spent yeast supplementation. Aquaculture 2013, 416–417, 322–327. [Google Scholar] [CrossRef]
- Vieira, E.F.; Ferreira, I.M. Antioxidant and antihypertensive hydrolysates obtained from by-products of cannery sardine and brewing industries. Int. J. Food Prop. 2016, 20, 662–673. [Google Scholar] [CrossRef]
- Vieira, E.F.; Pinho, O.; Ferreira, I.M. Bio-functional properties of sardine protein hydrolysates obtained by brewer’s spent yeast and commercial proteases. J. Sci. Food Agric. 2017, 97, 5414–5422. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.F.; Van Camp, J.; Ferreira, I.; Grootaert, C. Protein hydrolysate from canned sardine and brewing by-products improves TNF-α-induced inflammation in an intestinal–endothelial co-culture cell model. Eur. J. Nutr. 2017, 57, 2275–2286. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.F.; Da Silva, D.D.; Carmo, H.; Ferreira, I. Protective ability against oxidative stress of brewers’ spent grain protein hydrolysates. Food Chem. 2017, 228, 602–609. [Google Scholar] [CrossRef]
- Mathias, T.R.D.S.; De Aguiar, P.F.; Silva, J.B.D.A.E.; De Mello, P.P.M.; Sérvulo, E.F.C. Brewery Waste Reuse for Protease Production by Lactic Acid Fermentation. Food Technol. Biotechnol. 2017, 55, 218–224. [Google Scholar] [CrossRef]
- Djokoto, D.; Dzogbefia, V.; Oldham, J. Rapid Extraction of Pawpaw Juice with the Application of Locally Produced Pectic Enzymes from Saccharomyces Cerevisiae ATCC 51712. Food Biotechnol. 2006, 20, 31–41. [Google Scholar] [CrossRef]
- Dzogbefia, V.P.; Amoke, E.; Oldham, J.H.; Ellis, W.O. Production and Use of Yeast Pectolytic Enzymes to Aid Pineapple Juice Extraction. Food Biotechnol. 2001, 15, 25–34. [Google Scholar] [CrossRef]
- Martins, Z.E.; Pinho, O.; Ferreira, I. Impact of new ingredients obtained from brewer’s spent yeast on bread characteristics. J. Food Sci. Technol. 2018, 55, 1–6. [Google Scholar] [CrossRef]
- Pancrazio, G.; Cunha, S.C.; De Pinho, P.G.; Loureiro, M.; Meireles, S.; Ferreira, I.; Pinho, O. Spent brewer’s yeast extract as an ingredient in cooked hams. Meat Sci. 2016, 121, 382–389. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Yano, H. Comparison of Oxidized and Reduced Glutathione in the Bread-Making Qualities of Rice Batter. J. Food Sci. 2012, 77, C182–C188. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, C.; Albrecht, A.; Herrmann, J.; Strobl, M.; Jekle, M.; Becker, T. The contribution of glutathione to the destabilizing effect of yeast on wheat dough. Food Chem. 2015, 173, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Henrion, M.; Francey, C.; Lê, K.-A.; Lamothe, L. Cereal B-Glucans: The Impact of Processing and How It Affects Physiological Responses. Nutrients 2019, 11, 1729. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, M.S.; Bellini, M.F.; Angeli, J.P.F.; Oliveira, R.J.; Silva, A.F.; Ribeiro, L.R. β-Glucans in promoting health: Prevention against mutation and cancer. Mutat. Res. Rev. Mutat. Res. 2008, 658, 154–161. [Google Scholar] [CrossRef]
- Waszkiewicz-Robak, B.; Bartnikowska, E. Effects of spent brewer’s yeast and biological β-glucans on selected parameters of lipid metabolism in blood and liver in rats. J. Anim. Feed Sci. 2009, 18, 699–708. [Google Scholar] [CrossRef]
- Bell, S.; Goldman, V.M.; Bistrian, B.R.; Arnold, A.H.; Ostroff, G.; Forse, R.A. Effect of β-Glucan from Oats and Yeast on Serum Lipids. Crit. Rev. Food Sci. Nutr. 1999, 39, 189–202. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. A critical review on production and industrial applications of beta-glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Salgado, M.; Santos, F.; Rodríguez-Rojo, S.; Reis, R.L.; Duarte, A.R.C.; Cocero, M.J. Development of barley and yeast β-glucan aerogels for drug delivery by supercritical fluids. J. CO2 Util. 2017, 22, 262–269. [Google Scholar] [CrossRef]
- Marson, G.V.; Saturno, R.P.; Comunian, T.A.; Consoli, L.; Machado, M.T.D.C.; Hubinger, M.D. Maillard conjugates from spent brewer’s yeast by-product as an innovative encapsulating material. Food Res. Int. 2020, 136, 109365. [Google Scholar] [CrossRef]
- Guedes, J.D.S.; Pimentel, T.C.; Diniz-Silva, H.T.; Almeida, E.T.D.C.; Tavares, J.F.; De Souza, E.L.; Garcia, E.F.; Magnani, M. Protective effects of β-glucan extracted from spent brewer yeast during freeze-drying, storage and exposure to simulated gastrointestinal conditions of probiotic lactobacilli. LWT 2019, 116, 108496. [Google Scholar] [CrossRef]
- Vieira, E.F.; Das Neves, J.; Vitorino, R.; Dias-Da-Silva, D.; Carmo, H.; Ferreira, I. Impact of in Vitro Gastrointestinal Digestion and Transepithelial Transport on Antioxidant and ACE-Inhibitory Activities of Brewer’s Spent Yeast Autolysate. J. Agric. Food Chem. 2016, 64, 7335–7341. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.F.; Melo, A.; Ferreira, I.M. Autolysis of intracellular content of Brewer’s spent yeast to maximize ACE-inhibitory and antioxidant activities. LWT 2017, 82, 255–259. [Google Scholar] [CrossRef]
- Marson, G.V.; Machado, M.T.D.C.; De Castro, R.J.S.; Hubinger, M.D. Sequential hydrolysis of spent brewer’s yeast improved its physico-chemical characteristics and antioxidant properties: A strategy to transform waste into added-value biomolecules. Process. Biochem. 2019, 84, 91–102. [Google Scholar] [CrossRef]
- Komorowska, A.; Sieliwanowicz, B.; Mrówka, E.; Stecka, K.; Ha, A. Studies on yeast extracts enriched in 5′ nucleotides flavour enhancers obtained from spent brewery yeast. Electron. J. Polish Agric. Univ. 2003, 6, 3. [Google Scholar]
- Vieira, E.; Brandão, T.; Ferreira, I. Evaluation of Brewer’s Spent Yeast to Produce Flavor Enhancer Nucleotides: Influence of Serial Repitching. J. Agric. Food Chem. 2013, 61, 8724–8729. [Google Scholar] [CrossRef]
- Rakowska, R.; Sadowska, A.; Dybkowska, E.; Świderski, F. Spent yeast as natural source of functional food additives. Rocz. Państwowego Zakładu Hig. 2017, 68, 115–121. [Google Scholar]
- Varavinit, S.; Shobsngob, S.; Bhidyachakorawat, M.; Suphantharika, M. Production of Meat-Like Flavor. Sci. Asia 2000, 26, 219–224. [Google Scholar] [CrossRef]
- Alim, A.; Yang, C.; Song, H.; Liu, Y.; Zou, T.; Zhang, Y.; Zhang, S. The behavior of umami components in thermally treated yeast extract. Food Res. Int. 2019, 120, 534–543. [Google Scholar] [CrossRef]
- Cameron, J.S.; Moro, F.; Simmonds, H.A. Gout, uric acid and purine metabolism in paediatric nephrology. Pediatr. Nephrol. 1993, 7, 105–118. [Google Scholar] [CrossRef]
- Trevelyan, W.E. Chemical methods for the reduction of the purine content of baker’s yeast, a form of single-cell protein. J. Sci. Food Agric. 1976, 27, 225–230. [Google Scholar] [CrossRef]
- Canepa, A.; Pieber, M.; Romero, C.; Tohá, J.C. A method for large reduction of the nucleic acid content of yeast. Biotechnol. Bioeng. 1972, 14, 173–177. [Google Scholar] [CrossRef]
- Wiebe, M.G. Myco-protein from Fusarium venenatum: A well-established product for human consumption. Appl. Microbiol. Biotechnol. 2002, 58, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Trevelyan, W.E. Autolytic methods for the reduction of the purine content of baker’s yeast, a form of single-cell protein. J. Sci. Food Agric. 1976, 27, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.; Dwivedi, B. Production of Meat Substitutes from Spent Brewers’ Yeast and Soy Protein. Can. Inst. Food Technol. J. 1970, 3, 113–115. [Google Scholar] [CrossRef]
- Dwivedi, B.; Gibson, D.; Bell, J. Nutritive Value of Meat Substitute Prepared from Brewers’ Yeast and Isolated Soy Protein. Can. Inst. Food Sci. Technol. J. 1972, 5, 155–158. [Google Scholar] [CrossRef]
- Mathias, T.R.D.S.; Alexandre, V.M.F.; Cammarota, M.C.; De Mello, P.P.M.; Sérvulo, E.F.C. Characterization and determination of brewer’s solid wastes composition. J. Inst. Brew. 2015, 121, 400–404. [Google Scholar] [CrossRef]
- Radosavljević, M.; Pejin, J.; Pribić, M.; Kocić-Tanackov, S.; Mladenović, D.; Djukić-Vuković, A.; Mojović, L. Brewing and malting technology by-products as raw materials in L-(+)-lactic acid fermentation. J. Chem. Technol. Biotechnol. 2020, 95, 339–347. [Google Scholar] [CrossRef]
- Sawisit, A.; Seesan, S.; Chan, S.; Kanchanatawee, S.; Jantama, S.S.; Jantama, K. Validation of Fermentative Parameters for Efficient Succinate Production in Batch Operation by Actinobacillus succinogenes 130ZT. Adv. Mater. Res. 2012, 550, 1448–1454. [Google Scholar] [CrossRef]
- Neira, K.; Jeison, D. Anaerobic co-digestion of surplus yeast and wastewater to increase energy recovery in breweries. Water Sci. Technol. 2010, 61, 1129–1135. [Google Scholar] [CrossRef]
- Sosa-Hernández, O.; Parameswaran, P.; Alemán-Nava, G.S.; Torres, C.I.; Parra-Saldívar, R. Evaluating biochemical methane production from brewer’s spent yeast. J. Ind. Microbiol. Biotechnol. 2016, 43, 1195–1204. [Google Scholar] [CrossRef]
- Bocher, B.T.; Agler, M.; Garcia, M.L.; Beers, A.R.; Angenent, L.T. Anaerobic digestion of secondary residuals from an anaerobic bioreactor at a brewery to enhance bioenergy generation. J. Ind. Microbiol. Biotechnol. 2008, 35, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Zupančič, G.D.; Škrjanec, I.; Logar, R.M. Anaerobic co-digestion of excess brewery yeast in a granular biomass reactor to enhance the production of biomethane. Bioresour. Technol. 2012, 124, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Šafaříková, M.; Ptáčková, L.; Kibriková, I.; Šafařík, I. Biosorption of water-soluble dyes on magnetically modified Saccharomyces cerevisiae subsp. uvarum cells. Chemosphere 2005, 59, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, A.; Tavares, T.; Teixeira, J.A. Cr (III) removal and recovery from. Chem. Eng. J. 2004, 105, 11–20. [Google Scholar] [CrossRef]
- Riordan, C.; Bustard, M.; Putt, R.; McHale, A.P. Removal of uranium from solution using residual brewery yeast: Combined biosorption and precipitation. Biotechnol. Lett. 1997, 19, 385–388. [Google Scholar] [CrossRef]
- Machado, M.D.; Santos, M.S.F.; Gouveia, C.; Soares, H.M.V.M.; Soares, E.V. Removal of heavy metals using a brewer’s yeast strain of Saccharomyces cerevisiae: The flocculation as a separation process. Bioresour. Technol. 2008, 99, 2107–2115. [Google Scholar] [CrossRef]
- Laopaiboon, L.; Sridee, W.; Jaisil, P.; Laopaiboon, P. The use of dried spent yeast as a low-cost nitrogen supplement in ethanol fermentation from sweet sorghum juice under very high gravity conditions. Electron. J. Biotechnol. 2011, 14. [Google Scholar] [CrossRef]
- Kawa-Rygielska, J.; Pietrzak, W. Ethanol fermentation of very high gravity (VHG) maize mashes by Saccharomyces cerevisiae with spent brewer’s yeast supplementation. Biomass Bioenergy 2014, 60, 50–57. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Chemical Composition | Vieira et al. (2016) (g/100 g dw) | Cabellero-Cordoba and Sgarbieri (2000) (%) | Saksinchai et al. (2001) (%w/w) | Marson et al. (2020) (% dw) | Mathias et al. (2015) (% dw) | Jacob et al. (2019) (g/100 g) |
|---|---|---|---|---|---|---|
| Protein | 64.1+/−0.2 | 47.19 | n.d. | 40.8 | 45.6 | 74.3+/−0.5 |
| α-amino nitrogen | 12.9+/−0.1 | n.d. | 4.5+/−0.2 | n.d | 4.09+/−0.04 | 5.81+/−0.10 |
| Ash | 14.0+/−0.2 | 8.55 | 13.3+/−0.7 | 7.0+/−0.1 | 5.9+/−0.05 | 13.5+/−2.3 |
| Fat | 1.32+/−0.04 | 3.53 | n.d. | 0.21 | n.d. | 0.67+/−0.01 |
| Moisture | 7.70+/−0.12 | n.d. | n.d. | n.d | n.d. | 6.8 |
| Carbohydrate | 12.9+/−0.1 | 21.52 | 26.8+/−0.3 | n.d | n.d. | 14.7 |
| RNA | 4.00+/−0.16 | 7.04 | 21.3+/−0.4 (by orcinol) | 1.9+/−0.1 | n.d. | 5.518+/−0.10 |
| 23.2+/−0.6 (by UV260nm) | ||||||
| Soluble Fiber | n.d. | 9.65 | n.d. | 6.6+/−0.1 | n.d. | n.d. |
| Insoluble Fiber | n.d. | 2.57 | n.d. | n.d. | n.d. |
| Macrominerals | Vieira et al. (2016) mg/100 g dw | Jacob et al. (2019) mg/100 g dw | RDA (FSAI) |
|---|---|---|---|
| Sodium (Na) | 1228+/−22 | 88.1+/−0.001 | 1600 mg |
| Potassium (K) | 9148+/−69 | 6248.7+/−0.001 | 2000 mg |
| Calcium (Ca) | 27.1+/−0.40 | 16.4+/−0.001 | 800 mg |
| Magnesium (Mg) | 273+/−2.31 | 210.2+/−0.001 | 375 mg |
| Trace Elements | |||
| Chromium | 0.019+/−0.00 | n.a. | 40 μg |
| Iron | 1.76+/−0.03 | 3.672+/−0.01 | 14 mg |
| Manganese | 0.564+/−0.013 | 0.15+/−0.01 | 2 mg |
| Cobalt | 0.030+/−0.001 | 0.252+/−0.01 | n.a. |
| Molybdenum | 0.003+/−0.00 | n.a. | 50 μg |
| Zinc | 11.9+/−0.29 | 9.963+/−0.01 | 10 mg |
| Copper | 0.364+/−0.001 | 0.221+/−0.011 | 1 mg |
| Selenium | 0.030+/−0.00 | n.a. | 55 μg |
| Vitamins | Vieira et al. (2016) (mg/100 g) | Jacob et al. (2019) mg/100 g dw | Pinto et al. (2013) (mg/100 g) | RDA (FSAI) |
|---|---|---|---|---|
| Thiamine | NA | 7.46+/−0.5 | NA | 1.1 mg |
| Nicotinic acid (B3) | 77.2+/−1.1 | 78.6+/−2.0 | 0.79+/−0.06 | 16 mg |
| Pyridoxine (B6) | 55.1+/−2.5 | 5.90+/−0.5 | 9.99+/−0.06 | 1.4 mg |
| Folic Acid (B9) | 3.01+/−0.02 | 5.29+/−0.1 | 0.25+/−0.03 | 200 μg |
| Riboflavin (B2) | NQ (0.329) | 10.6+/−0.5 | 2.34+/−0.03 | 1.4 mg |
| Cyanocobalamin (B12) | NQ (0.256) | 0.16+/−0.1 | NQ | 2.5 μg |
| Sample | Suspension Medium | Total Solids Recovery (%) | Guanine (µmol/g) | Adenine (μmol/g) | Total Purines as RNA (%) |
|---|---|---|---|---|---|
| Pressed yeast cells after autoclaving | Water | 73.8 | 29 | 30 | 5.14 |
| 1.0 M NaCl | 69 | 5.9 | 6.6 | 1.14 | |
| Water | 69.4 | 24.4 | 25.4 | 4.62 | |
| 0.4 M NaCl | 66.3 | 12.1 | 12.6 | 2.39 | |
| 0.8 M NaCl | 66.1 | 4.5 | 4.8 | 0.91 | |
| 1.2 M NaCl | 66.8 | 3.2 | 4.2 | 0.71 | |
| 1.6 M NaCl | 67.2 | 2.6 | 3.3 | 0.57 | |
| 2.0 M NaCl | 69.1 | 3.9 | 3.4 | 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaeger, A.; Arendt, E.K.; Zannini, E.; Sahin, A.W. Brewer’s Spent Yeast (BSY), an Underutilized Brewing By-Product. Fermentation 2020, 6, 123. https://doi.org/10.3390/fermentation6040123
Jaeger A, Arendt EK, Zannini E, Sahin AW. Brewer’s Spent Yeast (BSY), an Underutilized Brewing By-Product. Fermentation. 2020; 6(4):123. https://doi.org/10.3390/fermentation6040123
Chicago/Turabian StyleJaeger, Alice, Elke K. Arendt, Emanuele Zannini, and Aylin W. Sahin. 2020. "Brewer’s Spent Yeast (BSY), an Underutilized Brewing By-Product" Fermentation 6, no. 4: 123. https://doi.org/10.3390/fermentation6040123
APA StyleJaeger, A., Arendt, E. K., Zannini, E., & Sahin, A. W. (2020). Brewer’s Spent Yeast (BSY), an Underutilized Brewing By-Product. Fermentation, 6(4), 123. https://doi.org/10.3390/fermentation6040123