Dual Extraction of Crustacean and Fungal Chitosan from a Single Mucor circinelloides Fermentation


Abstract
1. Introduction
2. Materials and Methods
2.1. Fermentation of Prawn Shell Waste using Mucor circinelloides Van Tieghem ATCC 24905
2.2. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
2.3. Elemental Analysis (EA)
2.4. Statistical Analysis
3. Results and Discussion
3.1. Dry Weights of Crustacean Chitin and Fungal Chitosan Extracted
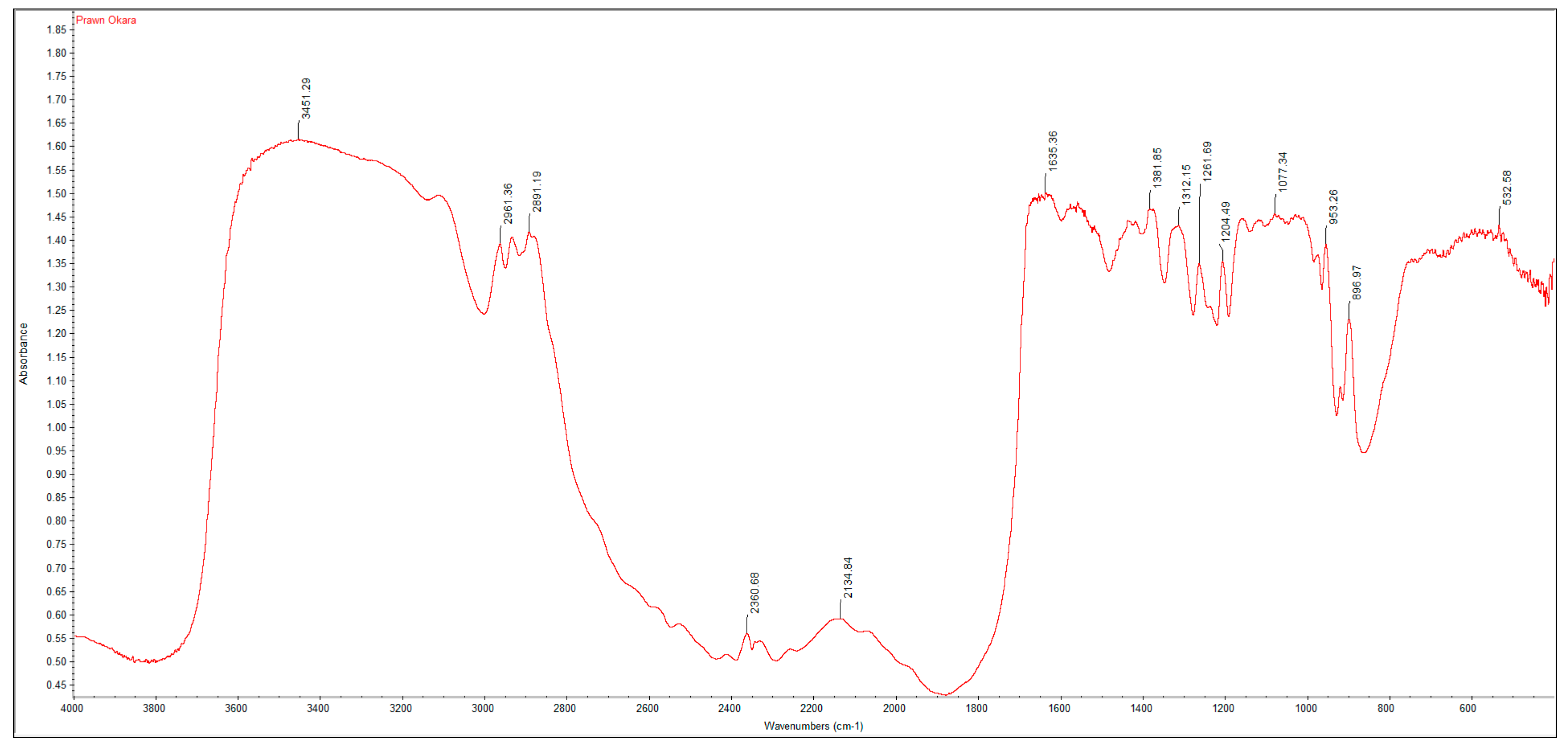
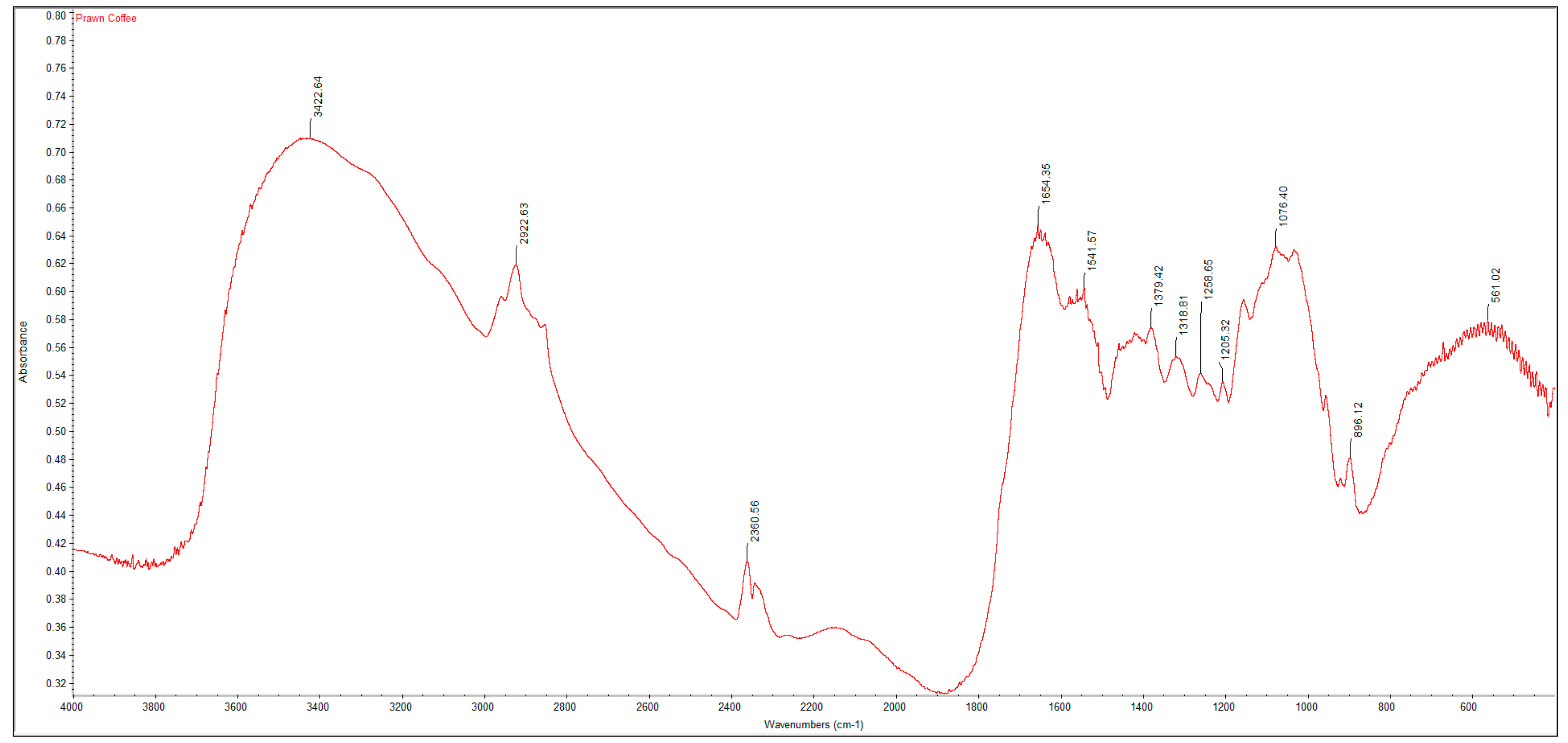
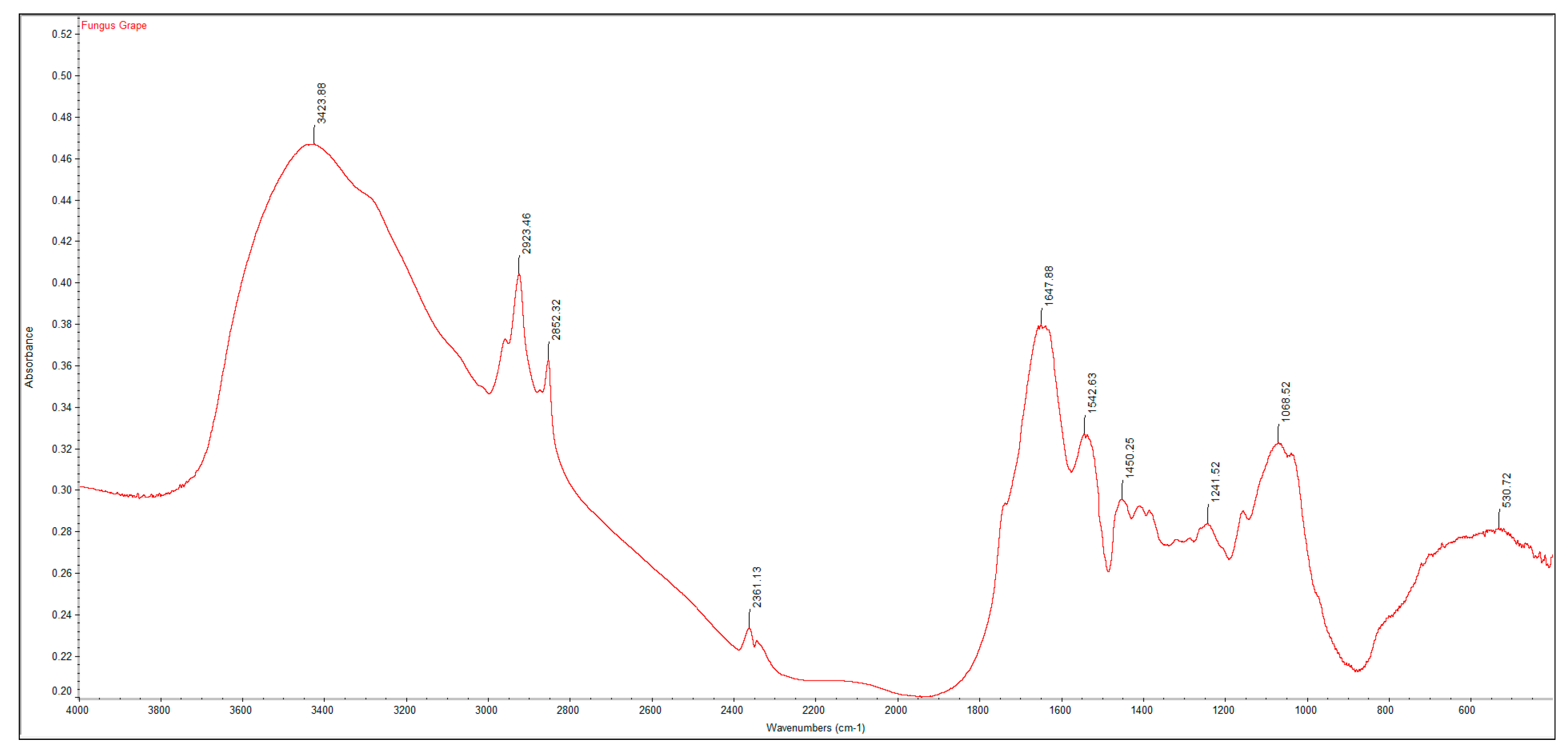
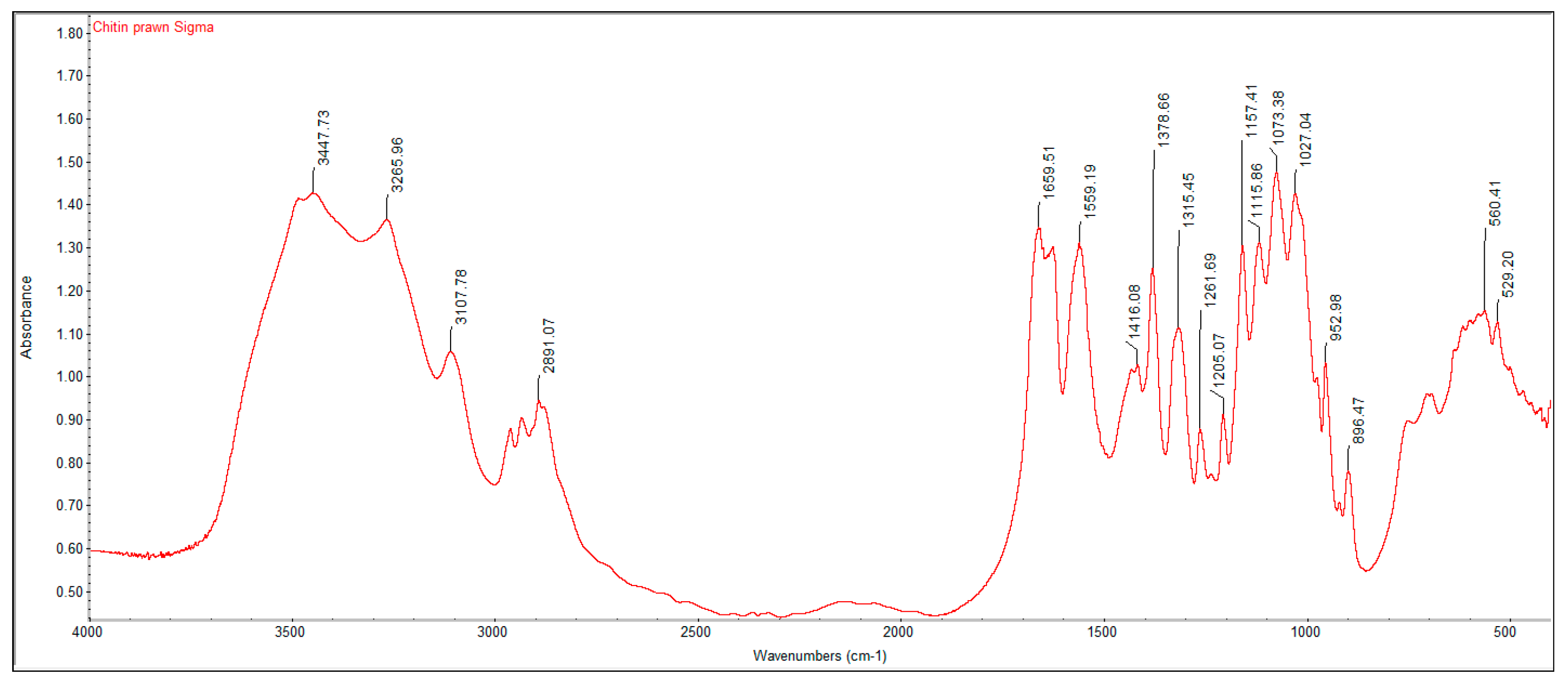
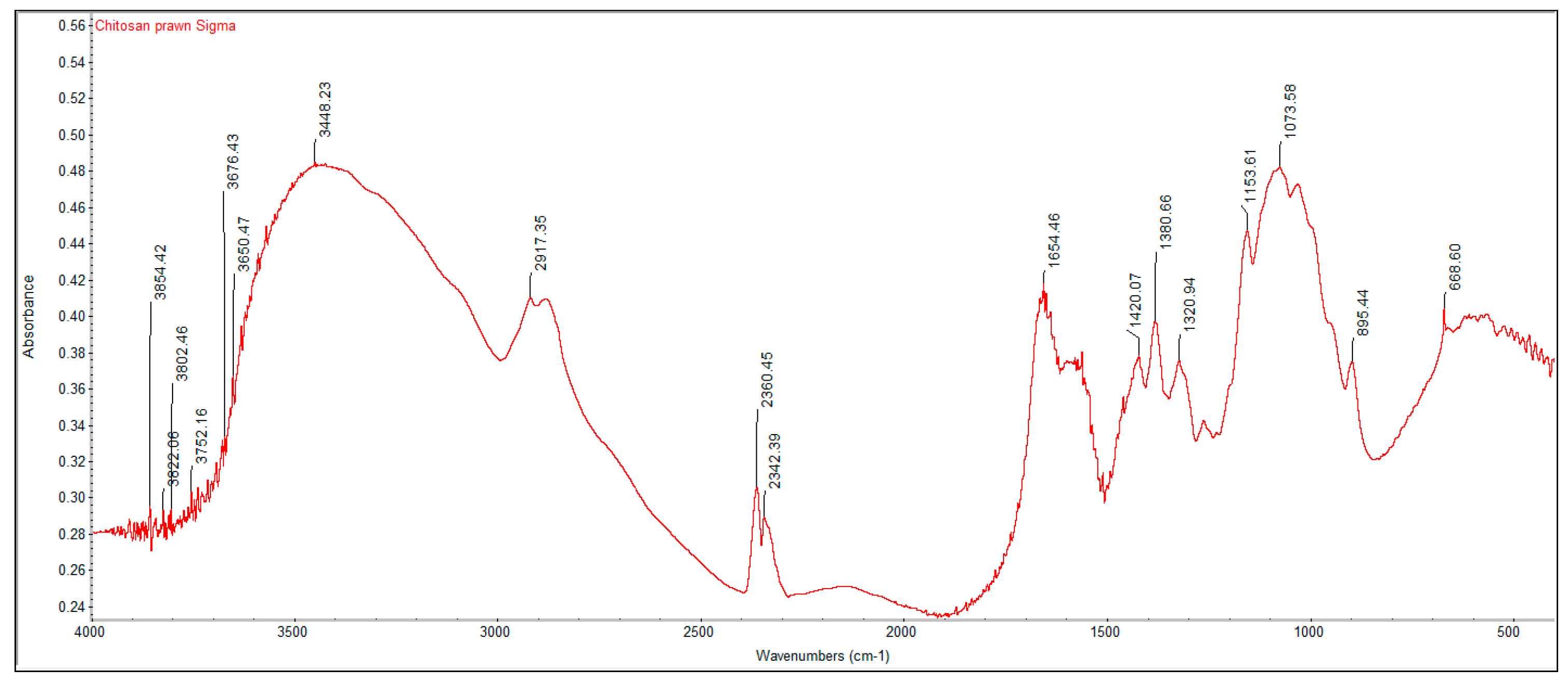
3.2. FTIR Spectrums and Degree of Deacetylation of Extracted Crustacean Chitin and Fungal Chitosan
3.3. Elemental Analysis of Extracted Crustacean Chitin and Fungal Chitosan
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sitanggang, A.B.; Sophia, L.; Wu, H.S. Aspects of glucosamine production using microorganisms. Int. Food Res. J. 2012, 19, 393–404. [Google Scholar]
- Nwe, N.; Furuike, T.; Tamura, H. Chitosan from Aquatic and Terrestrial Organisms and Microorganisms: Production, Properties and Applications. In Biodegradable Materials, 1st ed.; Johnson, B.M., Berkel, Z.E., Eds.; Nova Science Pub., Inc.: Hauppauge, NY, USA, 2011; pp. 29–50. [Google Scholar]
- Elieh-Ali-Komi, D.; Hamblin, M.R. Chitin and Chitosan: Production and Application of Versatile Biomedical Nanomaterials. Int. J. Adv. Res. 2016, 4, 411–427. [Google Scholar]
- Zukiewicz-Sobczak, W.; Sobczak, P.; Zawislak, K.; Zagorski, J.; Wojtyla-Buciora, P.; Wojtyla, A. Physical and chemical properties comparison of fungal and crustaceous chitosan. J. Health Inequal. 2015, 1, 7–14. [Google Scholar] [CrossRef]
- Ghormade, V.; Pathan, E.; Desphande, M. Can fungi compete with marine sources for chitosan production? Int. J. Biol. Macromol. 2017, 104, 1415–1421. [Google Scholar] [CrossRef]
- Islam, S.; Rahman Buiyan, M.A.; Islam, M.N. Chitin and Chitosan: Structure, Properties and Applications in Biomedical Engineering. J. Polym. Environ. 2017, 25, 854–866. [Google Scholar] [CrossRef]
- Ferreira, J.A.; Lennartsson, P.R.; Edebo, L.; Taherzadeh, M.J. Zygomycetes-based biorefinery: Present status and future prospects. Bioresour. Technol. 2013, 135, 523–532. [Google Scholar] [CrossRef]
- Younes, I.; Hajji, S.; Frachet, V.; Rinaudo, M.; Jellouli, K.; Nasri, M. Chitin extraction from shrimp shell using enzymatic treatment. Antitumor, antioxidant and antimicrobial activities of chitosan. Int. J. Biol. Macromol. 2014, 69, 489–498. [Google Scholar] [CrossRef]
- Abo Elsoud, M.M.; El Kady, E.M. Current trends in fungal biosynthesis of chitin and chitosan. Bull. Natl. Res. Cent. 2019, 43, 1–12. [Google Scholar] [CrossRef]
- Kaur, S.; Dhillon, G.S. The versatile biopolymer chitosan: Potential sources, evaluation of extraction methods and applications. Crit. Rev. Microbiol. 2014, 40, 155–175. [Google Scholar] [CrossRef]
- Philibert, T.; Lee, B.H.; Fabien, N. Current Status and New Perspectives on Chitin and Chitosan as Functional Biopolymers. Appl. Biochem. Biotech. 2017, 181, 1314–1337. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Green synthesis approach: Extraction of chitosan from fungus mycelia. Crit. Rev. Biotech. 2013, 33, 379–403. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Gao, X. Categories and biomanufacturing methods of glucosamine. Appl. Microbiol. Biot. 2019, 103, 7883–7889. [Google Scholar] [CrossRef] [PubMed]
- Aranday-Garcia, R.; Guerrero, A.R.; Ifuku, S.; Shirai, K. Successive inoculation of Lactobacillus brevis and Rhizopus oligosporus on shrimp wastes for recovery of chitin and added-value products. Process Chem. 2017, 58, 17–24. [Google Scholar] [CrossRef]
- Razak, M.A.; Pinjari, A.B.; Begum, P.S.; Viswanath, B. Biotechnological Production of Fungal Biopolymers Chitin and Chitosan: Their Potential Biomedical and Industrial Applications. Curr. Biotech. 2018, 7, 214–230. [Google Scholar] [CrossRef]
- Chan, L.G.; Cohen, J.L.; Nobrega de Moura Bell, J.M.L. Conversion of Agricultural Streams and Food-Processing By-Products to Value-Added Compounds Using Filamentous Fungi. Annu. Rev. Food Sci. Technol. 2018, 9, 503–523. [Google Scholar] [CrossRef]
- Rodrigues Reis, C.E.; Bento, H.B.S.; Carvalho, A.K.F.; Rajendran, A.; Hu, B.; De Castro, H.F. Critical applications of Mucor circinelloides within a biorefinery context. Crit. Rev. Biotech. 2019, 39, 555–570. [Google Scholar] [CrossRef]
- Stamford, T.; Freitas Silva, M.; Berger, L.; Anjos, F.; Alcantara, S.; Stamford, N.; Campos-Takaki, G. Chitin and chitosan produced by Mucoralean fungi using a new economic medium corn steep. In Microorganisms in Industry and Environment, 1st ed.; Mendez-Vilas, A., Ed.; World Scientific: Singapore, 2010; pp. 579–583. [Google Scholar]
- Sharifyazd, S.; Karimi, K. Effects of fermentation conditions on valuable products of ethanolic fungus Mucor indicus. Electron. J. Biotechn. 2017, 30, 77–82. [Google Scholar] [CrossRef]
- Teng, W.L.; Khor, E.; Tan, T.K.; Lim, Y.L.; Tan, S.C. Concurrent production of chitin from shrimp shells and fungi. Carbohyd. Res. 2001, 332, 305–316. [Google Scholar] [CrossRef]
- Anandhi, K. Biological Extraction of Chitin and Chitosan from Marine Fungi, its Characterization, Anti-Microbial Activity, Anti-Textile Activity against MDR Pathogens and Anti-Cancer Activity. World J. Pharm. Res. 2017, 6, 844–863. [Google Scholar]
- Stamford, T.C.M.; Stamford, T.L.M.; Stamford, N.P.; Neto, B.D.B.; Campos-Takaki, G.M.D. Growth of Cunninghamella elegans UCP 542 and production of chitin and chitosan using yam bean medium. Electron. J. Biotechnol. 2007, 10, 61–68. [Google Scholar]
- Franca, E.F.; Lins, R.D.; Freitas, L.C.G.; Straatsma, T.P. Characterization of Chitin and Chitosan Molecular Structure in Aqueous Solution. J. Chem. Theory Comput. 2008, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.R.R.; Cavalcante, H.M.D.M.; Stamford, T.C.M.; da Silva, M.C.D.F.; de Oliveira, C.E.V.; Sarmento, B.F.C.C.; de Campos-Takaki, G.M. Chitin and chitosan produced by Cunninghamella elegans using alternative medium–coconut water. In Microbes in Applied Research-Current Advances and Challenges, 1st ed.; Mendez-Vilas, A., Ed.; World Scientific: Malaga, Spain, 2012; Volume 1, pp. 377–381. [Google Scholar]
- Elizabeth, A.C.F.; Stamford, T.C.; Stamford-Arnaud, T.M.; D´Amorim Santa-Cruz, P.; Freitas da Silva, M.C.; Campos-Takaki, G.M.; Stamford, T.L. Physico-Chemical Characteristics and Functional Properties of Chitin and Chitosan Produced by Mucor circinelloides using Yam Bean as Substrate. Molecules 2011, 16, 10. [Google Scholar]
- Chatterjee, S.; Adhya, M.; Guha, A.; Chatterjee, B. Chitosan from Mucor circinelloides: Production and physico-chemical characterization. Process Chem. 2005, 40, 395–400. [Google Scholar]
- Patria, A. Production and characterization of Chitosan from shrimp shells waste. Aquac. Aquar. Conserv. Legis. Int. J. Bioflux Soc. 2013, 6, 339–344. [Google Scholar]
- Chatterjee, S.; Guha, A. A study on biochemical changes during cultivation of Rhizopus oryzae in deproteinized whey medium in relation to chitosan production. Lett. Appl. Microbiol. 2014, 59, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Devikrishna, S.; Remya, R. Effect of Degree of Deacetylation and Molecular Weight of Chitosan Extracted From Various Marine Sources on Its Applications. Int. J. Innov. Res. Sci. Eng. Technol. 2015, 4, 6078–6083. [Google Scholar]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H.; Chan, W.Y. Chitosan: An Update on Potential Biomedical and Pharmaceutical Applications. Mar. Drugs 2015, 13, 30. [Google Scholar] [CrossRef]
- Pochanavanich, P.; Suntornsuk, W. Fungal chitosan production and its characterization. Lett. Appl. Microbiol. 2002, 35, 4. [Google Scholar] [CrossRef]
- Corbin, K.R.; Hsieh, Y.S.; Betts, N.S.; Byrt, C.S.; Henderson, M.; Stork, J.; DeBolt, S.; Fincher, G.B.; Burton, R.A. Grape marc as a source of carbohydrates for bioethanol: Chemical composition, pre-treatment and saccharification. Bioresour. Technol. 2015, 193, 8. [Google Scholar] [CrossRef]
- Mulyati, A.; Widiastuti, D.; Oktaviani, L. Characterization of Durian Seed Flour (Durio zibhetinuss l.) and Estimation of its Self Life with Accelerated Self Life Testing (ASLT) Moisture Critical Method. J. Phy. Conf. Ser. 2018, 1095, 7. [Google Scholar] [CrossRef]
- Ejiofor, N.C.; Ezeagu, I.E.; Ayoola, M.B.; Umera, E.A. Determination of the Chemical Composition of Avocado (Persea Americana) Seed. Adv. Food Technol. Nutr. Sci. 2018, 2, 5. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Teixeira, J.A.; Mussatto, S.I. Chemical, Functional, and Structural Properties of Spent Coffee Grounds and Coffee Silverskin. Food Bioprocess Technol. 2014, 7, 11. [Google Scholar] [CrossRef]
- Chatterjee, S.; Chatterjee, B.; Guha, A.K. Kinetics of Mucor circinelloides Fermentation in Relation to Chitosan Production. Res. J. Microbiol. 2006, 1, 5. [Google Scholar]
- Stoykov, Y.M.; Pavlov, A.I.; Krastanov, A.I. Chitinase Biotechnology: Production, Purification and Application. Eng. Life Sci. 2015, 15, 9. [Google Scholar] [CrossRef]
- Zamani, A.; Edebo, L.; Sjostrom, B.; Taherzadeh, M.J. Extraction and Precipitation of Chitosan from Cell Wall of Zygomycetes Fungi by Dilute Sulfuric Acid. Biomacromolecules 2007, 8, 3786–3790. [Google Scholar] [CrossRef] [PubMed]
- White, S.A.; Farina, P.R.; Fulton, I. Production and Isolation of Chitosan from Mucor circinelloides. Appl. Environ. Microbiol. 1979, 38, 323–328. [Google Scholar] [CrossRef]
- Berger, L.R.R.; Stamford, T.C.M.; Stamford-Arnaud, T.M.; de Oliveira Franco, L.; do Nascimento, A.E.; Cavalcante, H.M.D.M.; Macedo, R.O.; de Campos-Takaki, G.M. Effect of Corn Steep Liquor (CSL) and Cassava Wastewater (CW) on Chitin and Chitosan Production by Cunninghamella elegans and Their Physicochemical Characteristics and Cytotoxicity. Molecules 2014, 19, 2771–2792. [Google Scholar] [CrossRef]
- Abasian, L.; Alavijeh, R.S.; Satari, B.; Karimi, K. Sustainable and Effective Chitosan Production by Dimorphic Fungus Mucor circinelloides via Replacing Yeast Extract with Fungal Extract. Appl. Biochem. Biotechnol. 2019, in press. [Google Scholar] [CrossRef]
- Sousa Andrade, V.; de Barros Neto, B.; Fukushima, K.; de Campos-Takaki, G.M. Effect of medium components and time of cultivation on chitin production by Mucor circinelloides (Mucor javanicus IFO 4570) -- a factorial study. Revista Iberoamericana de Micología 2003, 20, 149–153. [Google Scholar]
- Andrade, V.S.; Sarubbo, L.A.; Fukushima, K.; Miyaji, M.; Nishimura, K.; de Campos-Takaki, G.M. Production of Extracellular Proteases by Mucor circinelloides using D-Glucose as Carbon Source Substrate. Braz. J. Microbiol. 2002, 33, 106–110. [Google Scholar] [CrossRef]
- Karimi, K.; Zamani, A. Mucor indicus: Biology and industrial application perspectives: A review. Biotechnol. Adv. 2013, 31, 466–481. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Waste Substrate | Dry Weight of Crustacean Chitin (g) | Dry Weight of Fungal Chitosan (g) |
|---|---|---|
| 10 g of okara | 0.74 ± 0.01 | - |
| 10 g of coffee waste | 0.58 ± 0.01 | 1.0 ± 0.01 |
| 10 g of durian seed | 0.56 ± 0.01 | 0.77 ± 0.01 |
| 10 g of red grape pomace | 0.6 ± 0.01 | 0.76 ± 0.01 |
| 10 g of avocado seed | 1.13 ± 0.01 | 0.46 ± 0.01 |
| Functional Group | Commercial Chitin | Extracted Crustacean Chitin from Coffee Waste Fermentation | Commercial Chitosan | Extracted Fungal Chitosan from Red Grape Pomace Fermentation |
|---|---|---|---|---|
| O–H stretch | 3447 | 3423 | 3448 | 3424 |
| C–H stretch | 2891 | 2922 | 2917 | 2925 |
| C=O stretch of N-acetyl group (Amide I) | 1660 | 1654 | 1655 | 1648 |
| N–H bend, C–N stretch (Amide II) | 1559 | 1542 | 1540 | 1543 |
| CH3 in NHCOCH3 group | 1379 | 1379 | 1381 | 1450 |
| CH2 wagging (Amide III) | 1315 | 1319 | 1321 | 1242 |
| C–O–C stretch | 1073 | 1076 | 1074 | 1069 |
| Waste Substrate | Degree of Deacetylation (%) of Crustacean Chitin | Degree of Deacetylation (%) of Fungal Chitosan |
|---|---|---|
| Commercial Chitin | 70.5 ± 0.1 | - |
| Commercial Chitosan | - | 64.0 ± 0.1 |
| 10 g of okara | 69.9 ± 0.1 | - |
| 10 g of coffee waste | 68.8 ± 0.1 | 57.5 ± 0.1 |
| 10 g of durian seed | 63.1 ± 0.1 | 52.9 ± 0.1 |
| 10 g of red grape pomace | 62.7 ± 0.1 | 61.1 ± 0.1 |
| 10 g of avocado seed | 61.3 ± 0.1 | 42.1 ± 0.1 |
| Waste Substrate | %C | %H | %N | C/N Ratio of Crustacean Chitin |
|---|---|---|---|---|
| Commercial Chitin | 46.1 ± 0.1 | 7.3 ± 0.1 | 6.7 ± 0.1 | 6.9 ± 0.1 |
| 10 g of okara | 33.9 ± 0.1 | 10.5 ± 0.1 | 5.4 ± 0.1 | 6.3 ± 0.1 |
| 10 g of coffee waste | 43.1 ± 0.1 | 15.1 ± 0.1 | 5.2 ± 0.1 | 8.3 ± 0.1 |
| 10 g of durian seed | 38.8 ± 0.1 | 6.6 ± 0.1 | 5.7 ± 0.1 | 6.9 ± 0.1 |
| 10 g of red grape pomace | 43.8 ± 0.1 | 7.5 ± 0.1 | 6.9 ± 0.1 | 6.4 ± 0.1 |
| 10 g of avocado seed | 44.0 ± 0.1 | 5.8 ± 0.1 | 5.4 ± 0.1 | 8.1 ± 0.1 |
| Waste Substrate | %C | %H | %N | C/N Ratio of Fungal Chitosan |
|---|---|---|---|---|
| Commercial Chitosan | 43.5 ± 0.1 | 8.7 ± 0.1 | 7.8 ± 0.1 | 5.6 ± 0.1 |
| 10 g of coffee waste | 34.1 ± 0.1 | 6.6 ± 0.1 | 6.7 ± 0.1 | 5.1 ± 0.1 |
| 10 g of durian seed | 46.2 ± 0.1 | 6.8 ± 0.1 | 5.0 ± 0.1 | 9.2 ± 0.1 |
| 10 g of red grape pomace | 49.5 ± 0.1 | 7.0 ± 0.1 | 6.9 ± 0.1 | 7.2 ± 0.1 |
| 10 g of avocado seed | 43.4 ± 0.1 | 6.7 ± 0.1 | 8.5 ± 0.1 | 5.1 ± 0.1 |
| Waste Substrate | Extraction Efficiency of Crustacean Chitin | Substrate Conversion of Fungal Chitosan |
|---|---|---|
| 10 g of okara | 14.8% | - |
| 10 g of coffee waste | 11.6% | 10.0% |
| 10 g of durian seed | 11.2% | 7.7% |
| 10 g of red grape pomace | 12.0% | 7.6% |
| 10 g of avocado seed | 22.6% | 4.6% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, Y.N.; Lee, P.P.; Chen, W.N. Dual Extraction of Crustacean and Fungal Chitosan from a Single Mucor circinelloides Fermentation. Fermentation 2020, 6, 40. https://doi.org/10.3390/fermentation6020040
Tan YN, Lee PP, Chen WN. Dual Extraction of Crustacean and Fungal Chitosan from a Single Mucor circinelloides Fermentation. Fermentation. 2020; 6(2):40. https://doi.org/10.3390/fermentation6020040
Chicago/Turabian StyleTan, Yun Nian, Pei Pei Lee, and Wei Ning Chen. 2020. "Dual Extraction of Crustacean and Fungal Chitosan from a Single Mucor circinelloides Fermentation" Fermentation 6, no. 2: 40. https://doi.org/10.3390/fermentation6020040
APA StyleTan, Y. N., Lee, P. P., & Chen, W. N. (2020). Dual Extraction of Crustacean and Fungal Chitosan from a Single Mucor circinelloides Fermentation. Fermentation, 6(2), 40. https://doi.org/10.3390/fermentation6020040