1. Introduction
Pinot noir (
Vitis vinifera L.) is a challenging grape cultivar from both a viticultural and winemaking perspective. Viticulturally, Pinot noir grapevines produce compact clusters of thin-skinned berries, which increase susceptibility to fungal pathogens relative to other
V. vinifera cultivars. Pinot noir grapes (and their wines) are also inherently low in phenols [
1,
2]. Phenols are biomolecules originally present in the grapes (and subsequently extracted into wine). Phenols can be broadly classified as simple phenols having a C6–C1 or C6–C3 structure and a single aromatic ring containing one or more hydroxyl groups; and polyphenols, which contain multiple phenol rings and are defined by a C6–C3–C6 structure bearing hydroxyl and non-hydroxyl substitutions [
3]. In wines, polyphenols are responsible for color [
4,
5,
6], tactile sensations such as astringency [
7,
8], and taste sensations such as bitterness [
9,
10,
11]. In addition to sensory effects on astringency, taste, and aroma modulation [
12], flavonoids also play a critical role in the chemical stability of the wine during aging as these molecules intervene in metal-catalyzed oxidation reactions [
13,
14]. Because of the relatively lower phenolic content of Pinot noir, wines produced from it are lighter in color and astringency than wines made from other cultivars [
15]. Pinot noir is also notable for lacking acylated anthocyanins [
2], which are abundant in other cultivars such as Cabernet Sauvignon or Syrah and may in turn provide more stable color [
16,
17]. As color is one of the main drivers of perceived wine quality [
18], viticultural practices such as cluster thinning are often applied to Pinot noir grapes in an attempt to lower yields and influence fruit polyphenol composition by lowering vine crop load [
19,
20,
21,
22,
23,
24,
25].
Polyphenols such as anthocyanins and tannins, and their reaction products, known as polymeric pigments [
8], are positively associated with wine quality [
18]. In turn, and as mentioned above, vineyard crop load manipulation techniques such as cluster thinning are often applied to influence phenolic development [
19,
20,
21,
22,
23,
24,
25]. The traditional yield to fruit quality paradigm of a linear relationship with quality increasing as yield decreases [
24,
25,
26,
27], has been shown to be an oversimplification, and yields are more accurately described as a function of vine balance [
28,
29]. Vine balance, better described as the source/sink ratio, relates vine vegetative and reproductive growth, either through leaf area/yield (LA/Y) ratios [
30] or through the ratio between dormant vine pruning weights and yields, the latter known as the Ravaz Index [
31,
32]. For most cultivars, in warm climates, 0.8–1.2 m
2 of leaf area is needed to ripen 1 kg of fruit, which generally results in a yield/pruning weight ratio of 5 to 10 [
30].
Crop load metrics are dependent upon vine capacity, which is in turn influenced by regional and viticultural factors such as climate [
33], canopy training and trellising [
34], rootstock, and cultivar [
21,
35]. Grapes grown in cool climates require higher source/sink ratios than those grown in warmer regions because of lower daytime temperatures that restrict both leaf photosynthetic capacity and berry carbon assimilation [
29,
34,
36]. As a result of this restricted photosynthetic capacity, the ability of grapes grown in cool climates to ripen fruit to commercially viable total soluble solids (TSS) levels is limited and is significantly affected by seasonal variations in weather [
33,
34]. As such, in cool climates, seasonal variations in weather may have a greater effect than the source/sink ratio on Pinot noir ripening [
33].
Despite the prevalence of cluster thinning in Pinot noir, few studies have been conducted to investigate the effects of this viticultural technique on this cultivar. Indeed, most studies on cluster thinning have been conducted in warm, arid climates and on cultivars such as Cabernet Sauvignon [
37,
38] and Tempranillo [
22,
24,
39]. The bulk of this research suggests conflicting results, whereby it has not been conclusively shown that manipulating yields by cluster thinning will uniformly affect fruit composition. For example, some research has indicated that cluster thinning can positively impact fruit composition [
26,
27,
40], while other studies have found no effect on fruit composition [
21,
38,
41,
42] or that the effect of cluster thinning was dependent upon the climate conditions of any single growing season [
20,
21,
24]. In other instances, compositional effects as a result of cluster thinning have been found in grapes, but these have not translated into the finished wines [
27]. Pinot noir grown in cool climates may be more suited to benefit from cluster thinning because of the inherently low polyphenol content of the fruit and reduced carbon assimilation capacity of vines in cool climates [
29,
34,
36].
The timing of cluster thinning may also have an impact on vine physiology and fruit composition. For example, it has been hypothesized that removing crop at bloom may lead to lower leaf transpiration rates, and therefore lower leaf photosynthesis rates [
43], which could negate the desired effect of enhanced berry ripeness [
21]. In addition, if photosynthesis rates remain unchanged but the source/sink ratio increases upon cluster thinning, the increased photo-assimilates may stimulate vegetative growth, counteracting or negating the benefits of the decreased crop load [
29,
32]. In a study spanning five seasons, early thinning at bloom increased berry weight in Cabernet Sauvignon, Riesling, and Chenin blanc, while late thinning performed at véraison was intermediate to early thinning and non-thinned vines [
21]. However, this effect was not found in all years of the previous study. Cluster thinning applied at bloom to Pinot noir vines resulted in increased berry size [
19], although there was no late thinning treatment included in the study. Other research has shown no effect of thinning on berry size in Tempranillo [
22], thereby suggesting that there is likely a cultivar-specific response of berry size as a result of cluster thinning. Berry size reduction has traditionally been considered desirable from a winemaking perspective based on the empirical assumption that comparatively smaller berries have higher berry surface area/volume than larger ones. However, the relationship between berry size and phenolic composition is not a simple linear relationship, as berry skin and seed mass grow along with berry size [
44], and therefore larger berries may not be necessarily undesirable as once thought. Multiple studies conducted with a variety of cultivars investigating the relationship between cluster thinning and berry size have found conflicting results dependent upon cultivar and growing season [
19,
21,
22], and as such, there is no current conclusive understanding of the relationship between cluster thinning and berry size, and the subsequent effect of the latter on wine composition.
In the central coast of California (USA), consisting of Santa Barbara, San Luis Obispo, and Monterey counties, there are over 7000 hectares of Pinot noir being grown [
45], representing a substantial contribution to the wine industry of the region. Indeed, in 2016, wine grapes were the most valuable crop produced in San Luis Obispo County [
46]. Despite the economic importance of Pinot noir on the central coast of California, no research has been undertaken to understand the relationship between Pinot noir crop load and fruit quality in the cool climate of San Luis Obispo county of the central coast. While research in cooler areas such as Oregon’s Willamette Valley (USA) have indicated that grape ripeness increased in a curvilinear fashion with increasing LA/Y ratios up to 1.25–1.75 m
2/kg [
34], which is higher than the LA/Y ratios observed in warm climates [
30], these source/sink ratios are intrinsically tied to the seasonal limitations of the region that result in inconsistent ripening and may not be translatable to more moderate cool climates such as California’s central coast.
In the present study, the effect of crop load reduction by cluster thinning was explored for the first time on Pinot noir grapes (clone 115) and wines grown in the cool climate conditions of the Edna Valley of the Central Coast (San Luis Obispo County) of California (USA). The objectives of this study were to evaluate the effect of the timing of cluster thinning crop reduction on vine capacity, berry composition, and wine composition over two consecutive seasons. An additional objective was to identify appropriate crop loads in cool climate Pinot noir grown on the central coast of California.
4. Discussion
A study was conducted over two growing seasons (2016 and 2017) to determine the agronomical effects (on grapes) and chemical effects (on the resulting wines) of cluster thinning timing in Pinot noir grown in the Edna Valley of California’s central coast. A secondary objective of the study was to identify appropriate crop loads for Pinot noir on the moderately cool climate of California’s central coast. Cluster thinning treatments were applied at 4, 8, and 12 weeks post-bloom, approximating the timing of the phenological growth events of fruit set and véraison and including a pre-harvest “red drop”. Thinning consisted of removing any second or third cluster from fruit bearing shoots, reducing cluster number by an average of 34.3% across all treatments (
Table 2), and reducing yield by an average of 44.4% across all treatments (
Table 2). Previous studies conducted on cluster thinning practices have applied variable yield reduction rates depending on cultivar. However, most research in Pinot noir has applied “half crop” treatments such as the one performed in the current study, removing all but the basal cluster on each fruiting shoot [
34] or reducing cluster number by 50% [
19].
Growing degree day (GDD) accumulation varied between 2016 and 2017 enough to place each growing season in a separate Winkler Index region, with 2017 being warmer by 318.5 GDD than 2016 and qualifying as a region III (
Table 1). An increase in average temperature by 1 °C increases GDD by 214 over the course of a growing season. The GDD variation observed at this site between 2016 and 2017 corresponds to approximately 1.5 °C higher average temperatures in 2017. It is well documented that vine capacity and the ability to achieve ripeness by Brix accumulation in cool regions is dependent upon climatic conditions such as temperature [
48,
52,
53], with a base level of temperature required to adequately ripen fruit. However, Brix accumulation is not driven solely by temperature and, by extension, by GDD accumulation. In fact, differences in berry temperature may not directly impact Brix accumulation at all [
52]. Indeed, other factors such as soil moisture content [
54] and berry light exposure [
55] may also impact fruit ripening rate irrespective of atmospheric temperature and may even have a larger effect than GDD accumulation in cases where GDD accumulation is adequate for fruit ripening [
54]. Consequently, in the present study, there was no significant effect of the growing season on Brix at harvest to indicate an impact of the increased GDD accumulation in 2017 (
Table 3), despite the fruit being picked on the same date in both growing seasons.
In the present study, there was no impact of the thinning treatments on Brix accumulation. However, the climatic conditions of the growing season did have a clear impact on fruit pH and TA (
Table 3), resulting in lower TA and higher pH in 2017 fruit (
Table A2). While bloom + 12 fruit did have significantly lower pH than control fruit (
Table 3), after accounting for growing season in the model, the effect of cluster thinning treatment was not significant, indicating that the climatic conditions prevalent during the growing season had a greater effect on fruit pH and TA than cluster thinning timing. The decrease of malate concentration in grapes post-véraison through respiration increases with temperature and light exposure [
56,
57]; as such, the decrease in TA and the corresponding increase in pH observed in 2017, which was the warmer season (
Table A2), is likely a function of the increased temperature. There was a significant treatment × growing season interaction found in TA (
Table 3). In both growing seasons, bloom + 12 fruit had significantly different TA from the control treatment. However, the direction of the difference varied, with bloom + 12 having higher TA in 2016 and lower TA in 2017 relative to the control treatment. In both growing seasons, bloom + 4 was indistinguishable from the control treatment. Bloom + 8 was indistinguishable from the control or any treatment in 2016 and significantly lower than the control in 2017. Overall, there was no consistent effect of cluster thinning on fruit TA across both growing seasons (
Table 3). Previous studies on Pinot noir, in which half of the crop was thinned, have found increased pH [
19,
53] in both fruit and juice, but mixed effects on TA, showing either no impact [
19] or a reduction in TA with cluster thinning [
53]. While no significant treatment effect of pH was found in the present study after considering the growing season, the inconsistent results observed in bloom + 12 fruit is similar to what has been found in previous research, suggesting the influence of some external factor on malate degradation in late-thinned fruit. It is possible that late thinning resulted in more convective heat exchange between clusters and air within the canopy, which, given the substantially warmer air temperatures in August of 2017 (
Table A7), resulted in an increase in berry temperature and therefore malate degradation in the 2017 bloom + 12 fruit that was not seen in 2016. Indeed, cooler air temperatures in 2016 during the same period (
Table A7) may have resulted in decreased berry temperatures and the observed increase in fruit TA relative to the non-thinned control.
Wine pH is tied intrinsically to a wine’s microbial and oxidative stability, with lower pH values inhibiting (synergistically with ethanol) microbial growth [
58] and increasing the ability of phenolics to protect the wine from premature oxidation [
59]. There was a significant effect of thinning timing on wine pH, with bloom + 12 having lower wine pH relative to the control in accordance with the observed lowered fruit pH. Conversely, the bloom + 4 wines showed higher wine pH relative to the control wines despite indistinguishable fruit pH (
Table 3 and
Table 4). The growing season also significantly affected wine pH (
Table 4) with the warmer 2017 growing season resulting in higher wine pH than 2016, much like the effect seen on fruit pH (
Table A2). However, the interaction for treatment and growing season was not significant, suggesting that while seasonal variances in environmental factors do influence wine pH, environmental variance did not affect the response of wine pH to thinning timing. Interestingly, differences in fruit pH did not correspond linearly with differences in wine pH. Both growing season and treatment affected wine lactic acid content; fruit malic acid was measured in 2017 and no significant difference was found between treatments (
p = 0.38, df = 4.7; data not shown), indicating that a difference in fruit malic acid content was not responsible for the differences observed in lactic acid content in 2017. Different strains of malolactic bacteria were used in 2016 and 2017; VP-41 (
Oenococcus oeni) in 2016 and ML-Prime (
Lactobacillus Plantarum) in 2017. Unrelated to malic acid content,
Lactobacillus and
Oenococcus fermentation activity in wine is affected by wine temperature, ethanol level, pH, and acetic acid levels [
60], each of which exhibited some degree of variation within the wines that could be responsible for the differences observed between treatment groups in wine lactic acid and pH levels, irrespective of differences in fruit composition. Lower average and maximum fermentation temperatures in 2017 wines corresponded with higher wine pH and higher acetic acid levels (
Table A3 and
Table A4), indicating that fermentation temperature was a likely contributor to differences in wine pH and acetic acid levels. Indeed, a two-way ANOVA utilizing treatment, average temperature, maximum temperature, and treatment × average temperature and treatment × maximum temperature interactions, found average temperature to be a significant predictor of wine pH (
p = 0.0136) and acetic acid (
p = 0.0136).
Despite no differences being found in fruit Brix (
Table 3), wine ethanol was significantly higher in 2016 than 2017 (
Table A4). As yeast and fermentation practices were constant between growing seasons, observed differences in ethanol content are most likely the result of variations of the alcohol conversion ratio of the yeast. Average must temperature during the 10-day maceration period in 2016 was 21.3 °C and 24.9 °C in 2017, with peak fermentation temperatures of 26.7 °C and 29.9 °C, respectively (
Table A3). Similar to
Lactobacillus and
Oenococcus fermentation activity, temperature is one of the most influential factors on
Saccharomyces cerevisiae fermentation activity and ethanol biosynthesis [
61,
62], with ethanol formation decreasing as fermentation temperature increases [
61]. In addition to decreasing ethanol biosynthesis in wine yeast, increased fermentation temperatures also increase the rate of ethanol volatilization, further lowering already diminished wine ethanol levels [
61].
The growing season also affected cluster and berry weight (
Table 2), with 2017 resulting in fruit having 34% higher cluster weight and 9% lower berry weight relative to 2016 (
Table A1). Several factors influence berry size, including berry temperature during various phenological growth stages, light incidence, water and nutrient supply, and seed number per berry [
63,
64,
65,
66]. Berry size can be reduced by increased heat and resultant increased berry temperature prior to the lag phase [
64]. Ambient temperature was on average 1.25 °C warmer in May 2017 than May 2016 (
Table A7), which may explain current results. Cluster weight is a function of berry size and berry number, so as berry weight decreased while cluster weight increased, berry number must have increased in 2017. Typical berry set in wine grapes ranges from 20% to 50% [
67], and can be reduced by temperatures during bloom below 15 °C [
64]. From 15 April to 1 June, there were 29 days with an average air temperature below 15 °C in 2016 and 14 days in 2017 (
Table A7). It is likely that warmer temperatures during bloom in 2017 resulted in a higher fruit set and therefore higher cluster weight than 2016.
Pruning weights were collected in January 2018 and Ravaz Index was calculated for the 2017 growing season. The non-thinned control vines had a Ravaz Index of 3.23, and Ravaz Index was not significantly different between treatments (
Table A1). Within the control vines, one replication had a substantially lower cluster number than other replications, which inflated the deviation of cluster number, vine yield, and Ravaz Index of the sampled population (
Table A1). This was confirmed by conducting outlier analysis of control treatment repetition 1, which, for the Ravaz Index model, had a Cook’s Distance value of 16 (data not shown), indicating high influence on the model. It is possible that the low cluster number on the vines within this repetition is due to natural site variation (e.g., block to block variations in soil composition or water holding capacity), affecting vine capacity. While the abnormally low cluster number affected vine yield, cluster number, and Ravaz Index, little influence of this repetition was found in models of fruit or wine chemistry, with no Cook’s distance value above 0.5 (data not shown). As a result of the potential impact of eliminating one of the three replications of the control treatment from the dataset on experimental balance and statistical analysis, the outlier was retained within the dataset.
Phenolic composition (
Table 5) and wine color (
Table 6) were not affected consistently by any of the thinning treatments, but all chromatic parameters as well as wine polymeric pigment, tannin, and non-tannin phenolic content were significantly affected by growing season. Polymeric pigments, formed by the covalent polymerization of anthocyanins with monomeric flavan-3-ols or tannins [
8] provide protection for anthocyanins against oxidation [
68,
69], which can be beneficial in wines made from cultivars lacking in phenolics, such as Pinot noir. However, as their formation may also lower wine saturation on accounts of their lower molar extinction coefficient relative to that of intact anthocyanins [
70], increasing polymeric pigments may result in comparative decreases in wine color saturation. Notwithstanding, polymeric pigments generally provide desirable mouthfeel properties as they are less astringent than tannins of the same molecular weight [
8]. Polymeric pigments and tannins were lower in 2017, while non-tannin phenolics were higher (
Table A6). While some parameters (total phenolics, non-tannin phenolics, L*, a*, chroma) were affected by thinning treatment in 2017 (
Table A6), no consistent effect of treatment or treatment × growing season interaction was found in any wine phenolic or chromatic parameter. Polymeric pigments were likely higher in 2016 because of the increased level of tannins observed (
Table A6), despite no difference in anthocyanin levels between growing seasons.
Wine CIE L*a*b* color parameters L*, b*, and hue angle were higher in 2017 than 2016, while a* and chroma were lower, indicating wine color was darker and bluer in the wines from the warmer growing season, but less saturated and red than wines from the cooler growing season. The color shift observed in 2017 wines is likely due to differences in wine pH and polymeric pigment content. The effect of pH on wine color and anthocyanin chromatic parameters is well established. As pH decreases, the equilibrium of anthocyanin forms shifts to favor the flavilium cation, which increases red hue, and as pH increases, the equilibrium shifts to favor the quinonoidal hydrobase, which increased blue hue [
71]. Additionally, as pH increases, there is a linear decrease in chroma value observed [
71]. As wine pH was generally higher in the 2017 wines (
Table A4), it would follow that the color in 2017 wines, while not having statistically distinguishable anthocyanin concentration relative to 2016 wines, would have increased blue hue and lower chroma. Polymeric pigment formation may lower saturation (as indicated by chroma). Saturation may be lowered through the transformation (and subsequent reduction) of anthocyanins, or through the modulation of the chromatic properties of the anthocyanin subunit following a reduction of the molar extinction coefficient relative to the native anthocyanin, although there is only indirect experimental evidence of this molar extinction coefficient reduction [
72,
73].
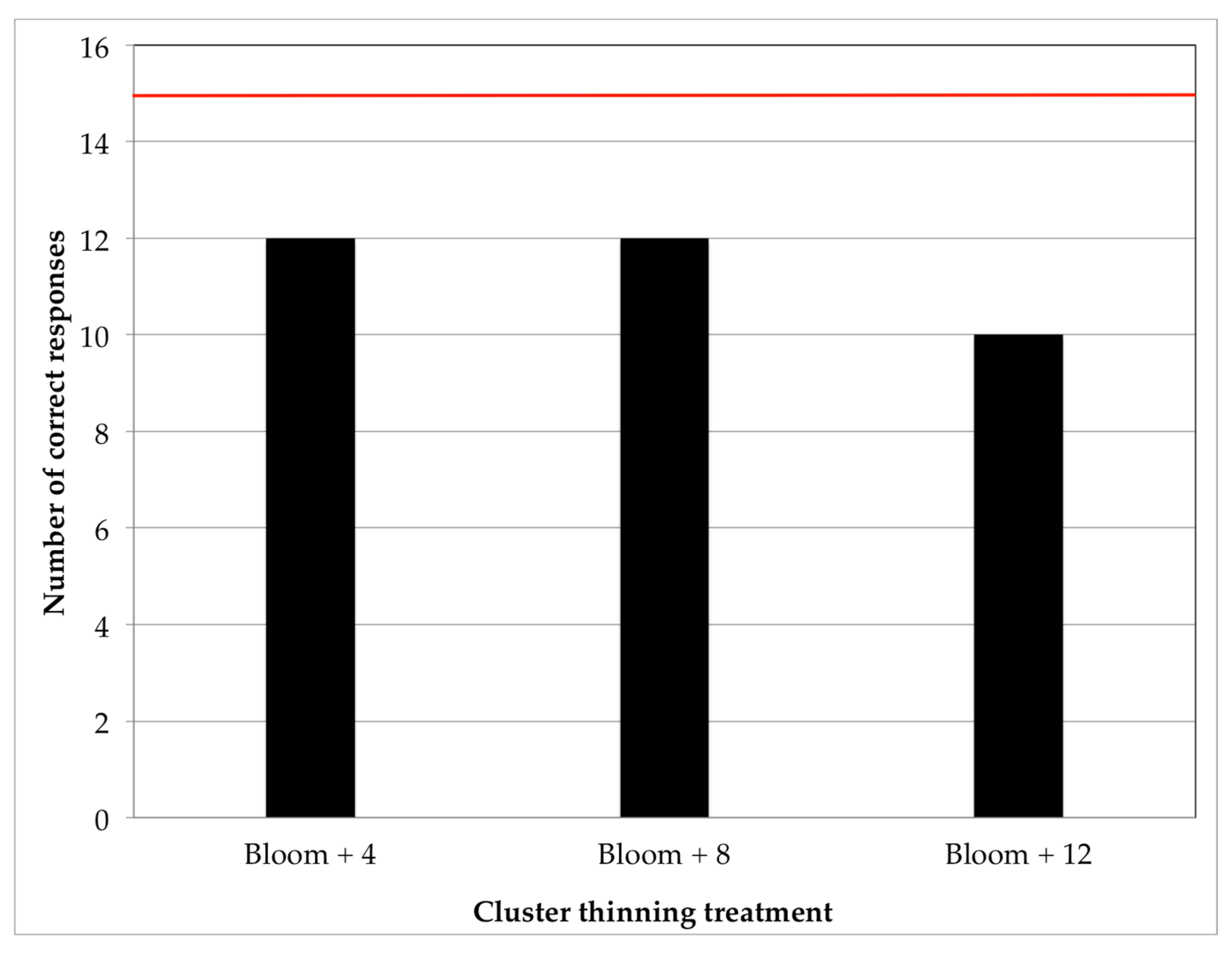
The results of the overall sensory test performed in the 2016 wines generally mirrored previously uncovered trends in the basic, phenolic, and chromatic composition of the resulting wines. That is, none of the cluster thinning treatments produced wines that were distinguishable, from a sensory standpoint, from the control wines. Similar to what has been found in the present study, cluster thinning performed in Chardonnay Musqué grapes, while producing chemical differences in fruit, resulted in little sensory differences in the resulting wines [
27]. In another study, wine produced from cluster thinned Cabernet Sauvignon vines exhibited a small increase in perceived wine quality relative to wine produced from non-thinned vines [
40], although location was found to have a greater impact on sensory perception than the cluster thinning treatment, and the effect was not consistent across growing seasons. While wine chemical composition and perceived sensory attributes rarely follow linear correlations, without corresponding differences in chemical composition, it is unlikely that cluster thinning will have an impact on wine sensory perception. Therefore, the lack of sensory differences observed in the wines of the present study is unsurprising, and we hypothesize that any chemical differences in volatiles that may have occurred within the wines were below sensory thresholds, and therefore practically irrelevant. Unless cluster thinning is necessitated by the vine balance (i.e., the vine is overcropped) in cool climate Pinot noir, it is unlikely that there will be any sensory benefit to cluster thinning that would justify the negative economic impact associated with cluster thinning.
In much of the previous research performed on cluster thinning, external factors independent of (although at times in combination with) vine crop load impacted berry and wine chemical composition to a greater degree than crop load. Factors such as climatic variation in growing season [
21] and viticultural practices such as floor management [
34], deficit irrigation [
21,
22], and leaf thinning [
74,
75] have all been found to have a greater effect on fruit composition than cluster thinning. Indeed, in the present study, no consistent effect of cluster thinning or cluster thinning timing was observed across two growing seasons, a cooler growing season and a warmer growing season. Conversely, the growing season had a greater effect on variation in fruit and wine composition than thinning treatment for most parameters. A Ravaz Index range of 3 to 6 has been previously proposed as an optimum crop load for Pinot noir grown in cool-climates [
30]. Based on the lack of differences observed in fruit Brix accumulation, wine composition, and wine sensory perception, Pinot noir vineyards on the central coast of California can, barring climatic conditions severely increasing crop set or severely limiting ripening potential, likely support higher crop levels than those of the vineyard utilized in this this study, which had Ravaz Index values of 3.23 across the non-thinned blocks. Considering previously proposed ranges and the results of the current study, a Ravaz Index value of 6 could be appropriate for Pinot noir on the central coast of California, and should be examined and evaluated accordingly in future work.






{kind=link}
{kind=link}
