Acetogen Communities in the Gut of Herbivores and Their Potential Role in Syngas Fermentation

Abstract
1. Introduction
2. Results
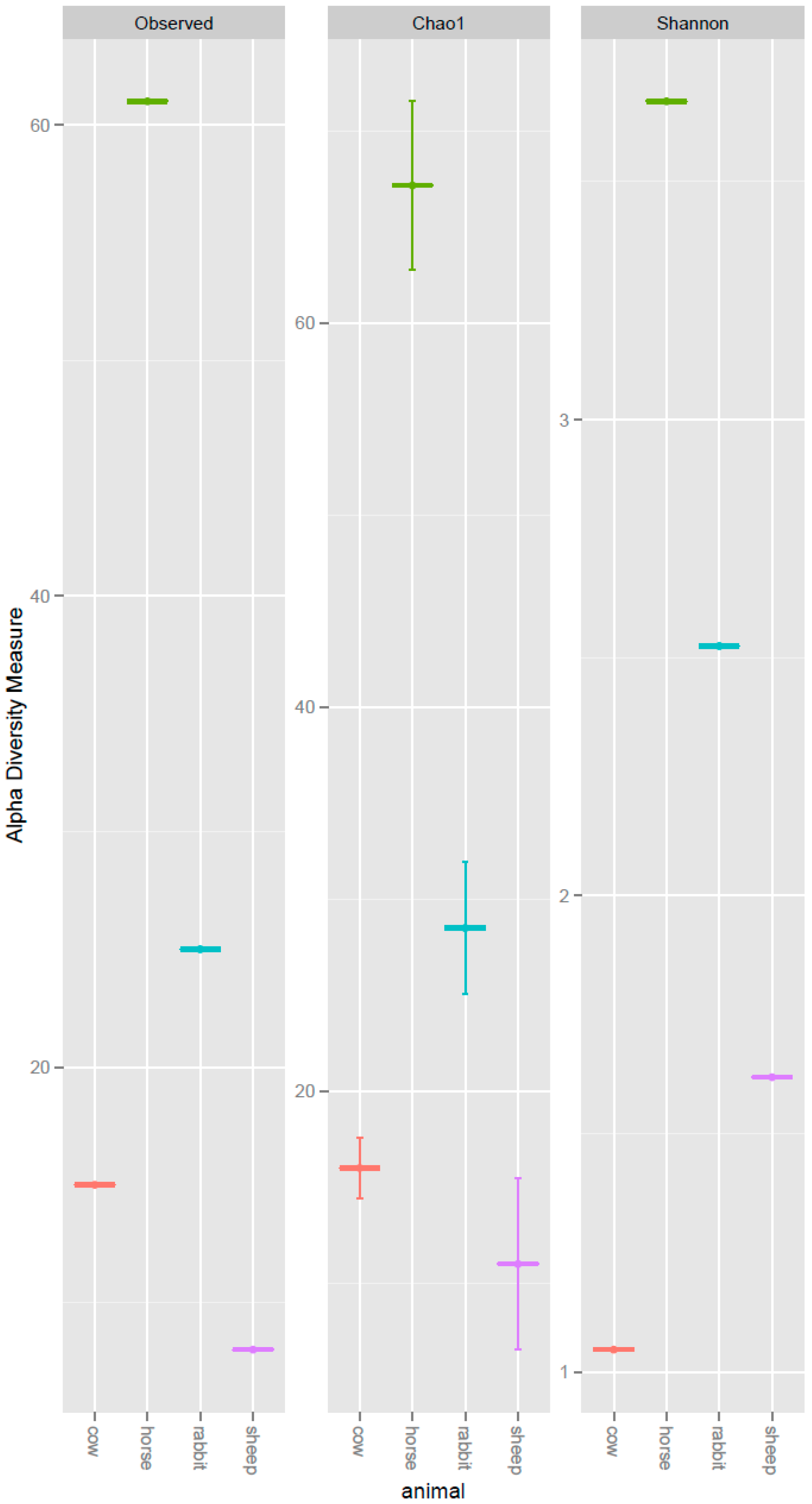
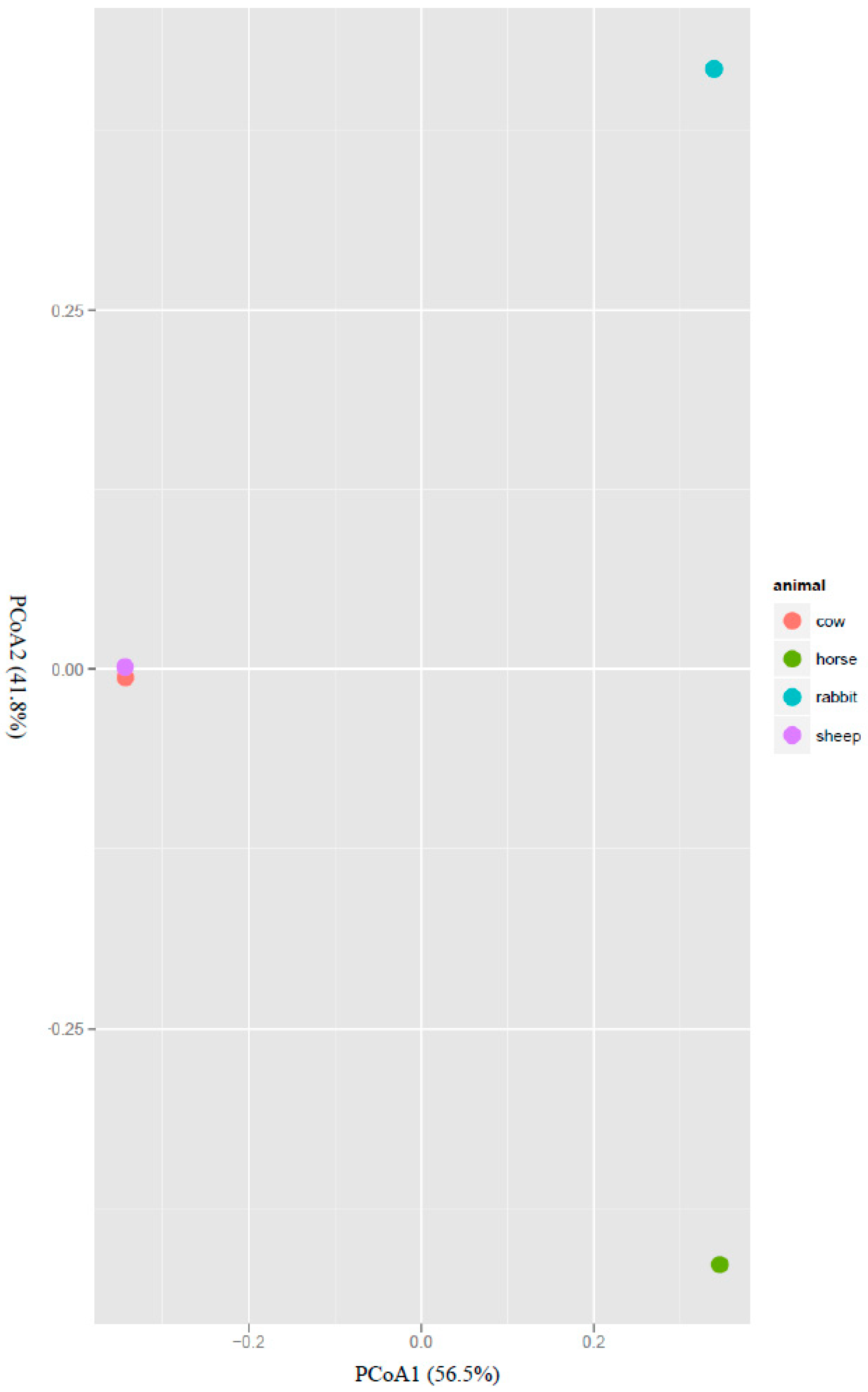
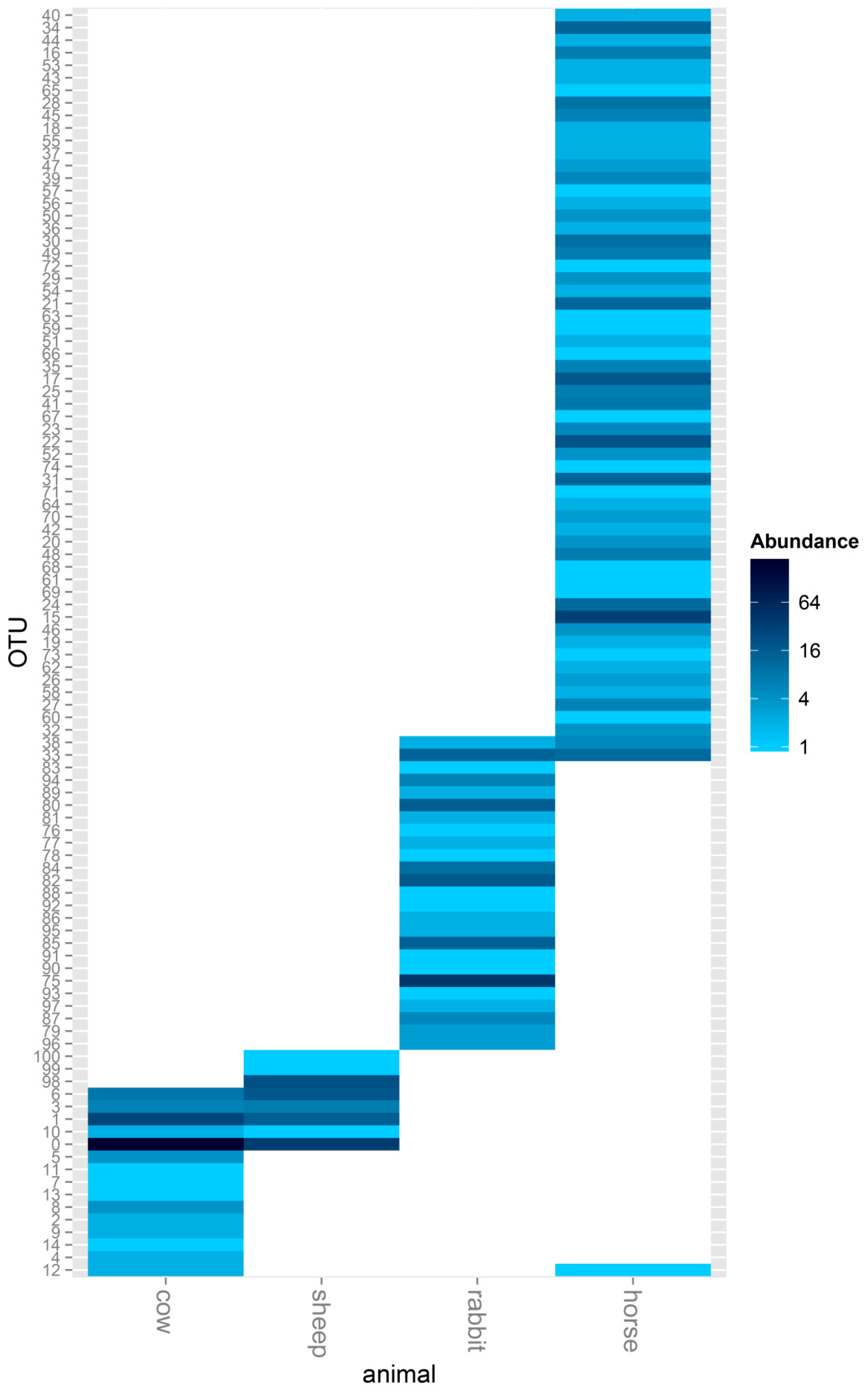
2.1. Comparison of the Acetogen Communities
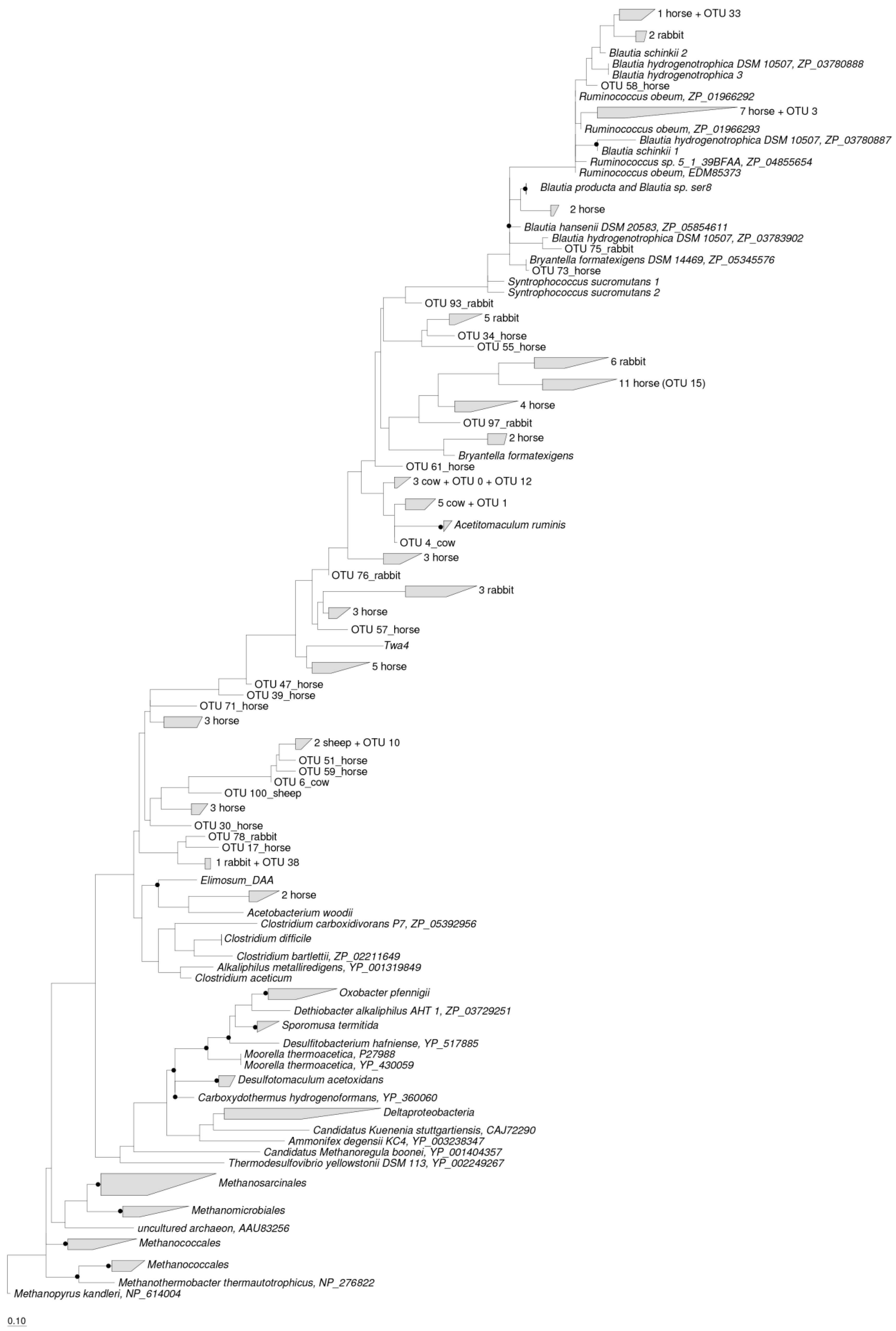
2.2. Phylogenetic Analysis of Acetogen Communities
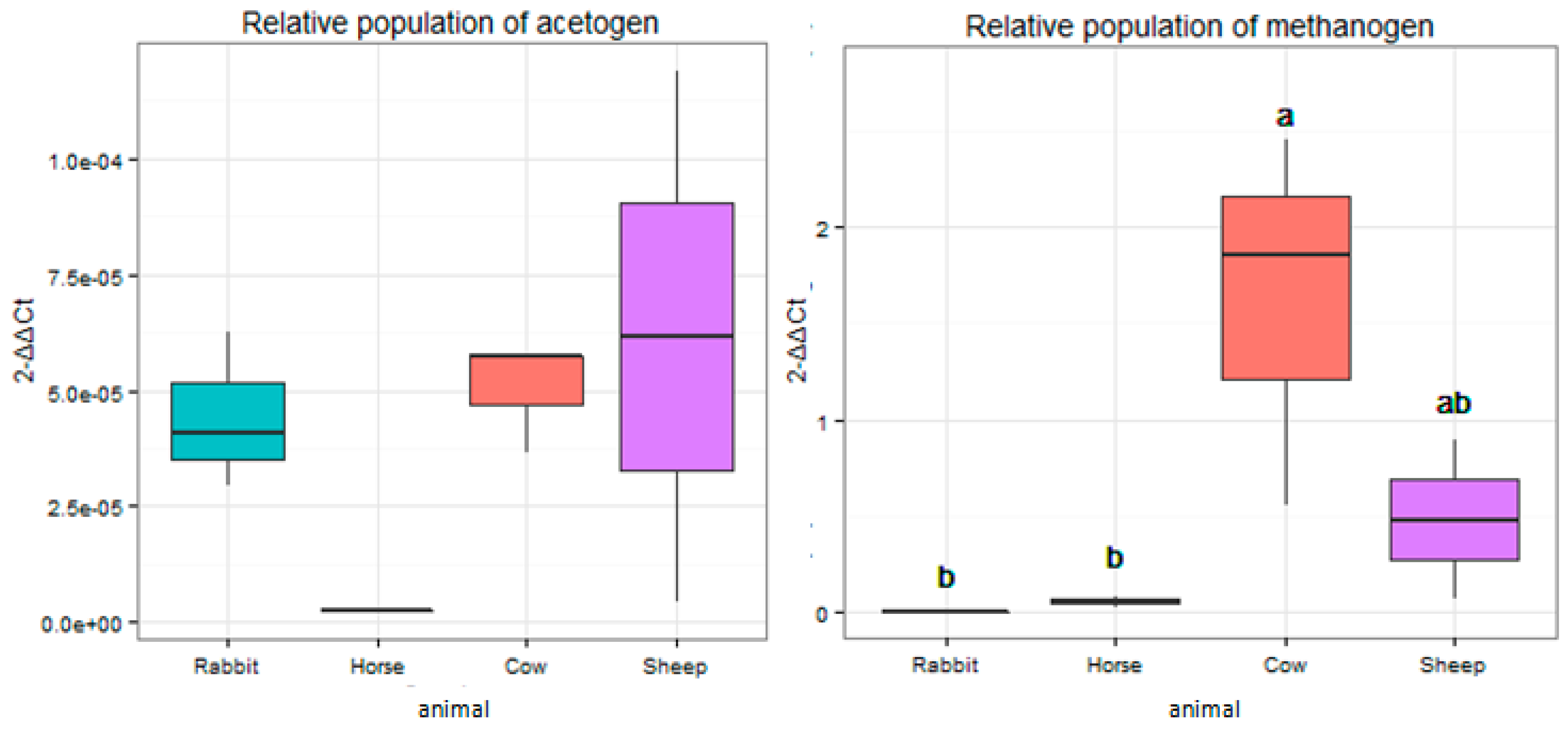
2.3. Relative Abundances of Acetogens and Methanogens
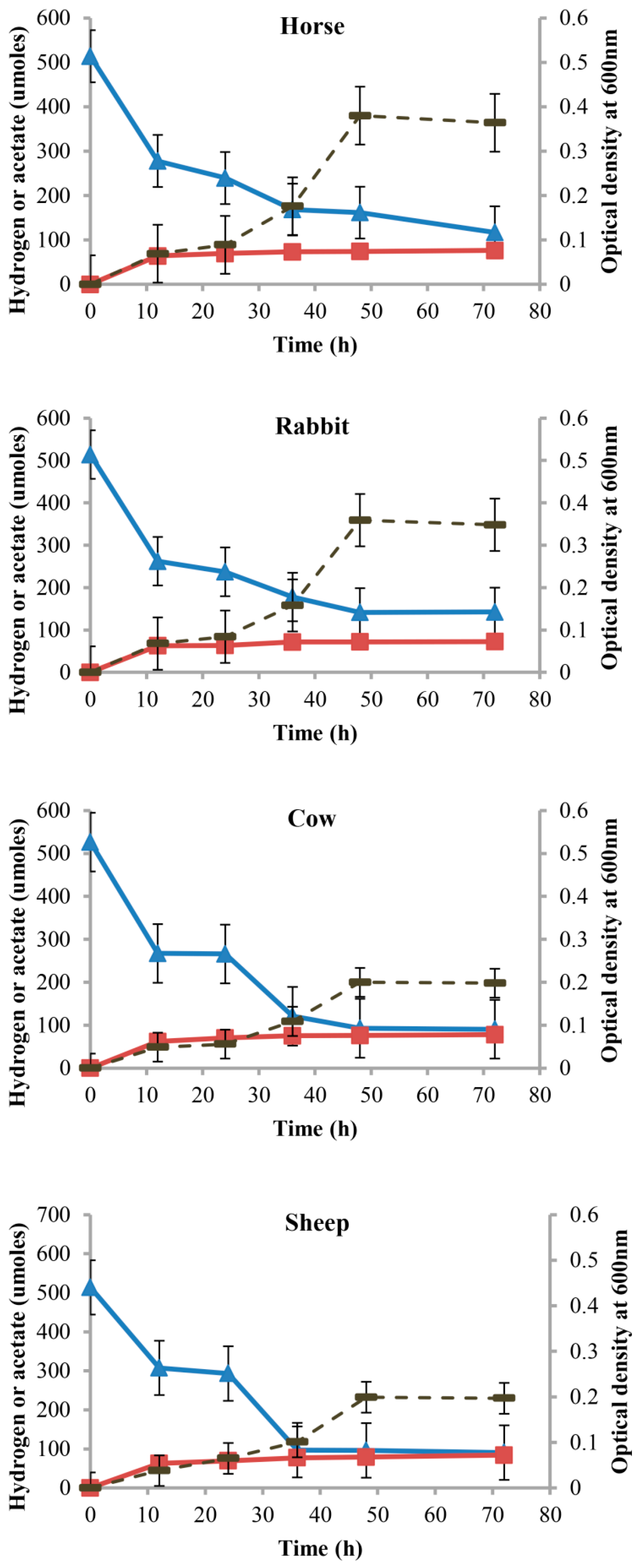
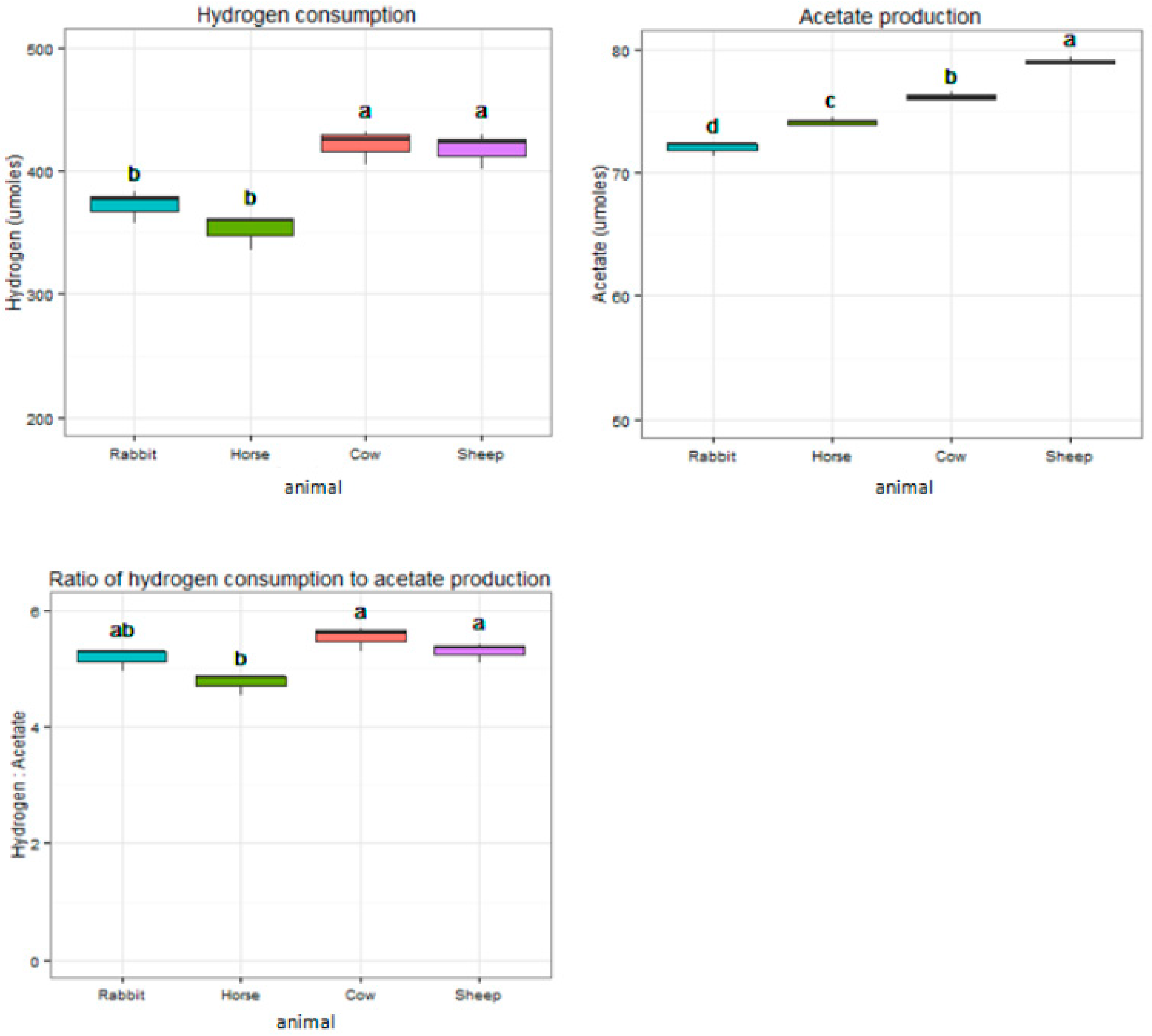
2.4. Hydrogenotrophic Characteristics of Gut Acetogen Enrichments
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Total DNA Extraction and Real-Time Quantitative PCR
4.3. Analysis of Acetogen Community
4.4. Enrichment Culture for Potential Acetogens
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Drake:, H.L. Acetogenesis, acetogenic bacteria, and the acetyl-CoA “Wood/Ljungdahl” pathway: Past and current perspectives. In Acetogenesis; Springer: Boston, MA, USA, 1994. [Google Scholar]
- Wood, H. Life with CO or CO2 and H2 as a source of carbon and energy. FASEB J. 1991, 5, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Drake, H.L.; Küsel, K.; Matthies, C. Acetogenic prokaryotes. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Eds.; Springer-Verlag: New York, NY, USA, 2006. [Google Scholar]
- Drake, H.L.; Gößner, A.S.; Daniel, S.L. Old acetogens, new light. Ann. N. Y. Acad. Sci. 2008, 1125, 100–128. [Google Scholar] [CrossRef] [PubMed]
- Bengelsdorf, F.R.; Straub, M.; Dürre, P. Bacterial synthesis gas (syngas) fermentation. Environ. Technol. 2013, 34, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Fast, A.G.; Papoutsakis, E.T. Stoichiometric and energetic analyses of non-photosynthetic CO2-fixation pathways to support synthetic biology strategies for production of fuels and chemicals. Curr. Opin. Chem. Eng. 2012, 1, 380–395. [Google Scholar] [CrossRef]
- Henstra, A.M.; Sipma, J.; Rinzema, A.; Stams, A.J. Microbiology of synthesis gas fermentation for biofuel production. Curr. Opin. Biotechnol. 2007, 18, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Nevin, K.P. Electrobiocommodities: Powering microbial production of fuels and commodity chemicals from carbon dioxide with electricity. Curr. Opin. Biotechnol. 2013, 24, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Bruant, G.; Lévesque, M.J.; Peter, C.; Guiot, S.R.; Masson, L. Genomic analysis of carbon monoxide utilization and butanol production by Clostridium carboxidivorans strain P7. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Köpke, M.; Mihalcea, C.; Liew, F.; Tizard, J.H.; Ali, M.S.; Conolly, J.J.; Al-Sinawi, B.; Simpson, S.D. 2,3-Butanediol production by acetogenic bacteria, an alternative route to chemical synthesis, using industrial waste gas. Appl. Environ. Microbiol. 2011, 77, 5467–5475. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Najafpour, G.D.; Younesi, H.; Lahijani, P.; Uzir, M.H.; Mohamed, A.R. Bioconversion of synthesis gas to second generation biofuels: A review. Renew. Sustain. Energy Rev. 2011, 15, 4255–4273. [Google Scholar] [CrossRef]
- Ukpong, M.N.; Atiyeh, H.K.; De Lorme, M.J.; Liu, K.; Zhu, X.; Tanner, R.S.; Wilkins, M.R.; Stevenson, B.S. Physiological response of Clostridium carboxidivorans during conversion of synthesis gas to solvents in a gas-fed bioreactor. Biotechnol. Bioeng. 2012, 109, 2720–2728. [Google Scholar] [CrossRef] [PubMed]
- Demler, M.; Weuster-Botz, D. Reaction engineering analysis of hydrogenotrophic production of acetic acid by Acetobacterium woodii. Biotechnol. Bioeng. 2011, 108, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Leang, C.; Ueki, T.; Nevin, K.P.; Lovley, D.R. Lactoseinducible system for metabolic engineering of Clostridium ljungdahlii. Appl. Environ. Microbiol. 2014, 80, 2410–2416. [Google Scholar] [CrossRef] [PubMed]
- Köpke, M.; Held, C.; Hujer, S.; Liesegang, H.; Wiezer, A.; Wollherr, A.; Ehrenreich, A.; Liebl, W.; Gottschalk, G.; Dürre, P. Clostridium ljungdahlii represents a microbial production platform based on syngas. Proc. Natl. Acad. Sci. USA 2010, 107, 13087–13092. [Google Scholar] [CrossRef] [PubMed]
- Ueki, T.; Nevin, K.P.; Woodard, T.L.; Lovley, D.R. Converting carbon dioxide to butyrate with an engineered strain of Clostridium ljungdahlii. mBio 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Kita, A.; Iwasaki, Y.; Sakai, S.; Okuto, S.; Takaoka, K.; Suzuki, T.; Yano, S.; Sawayama, S.; Tajima, T.; Kato, J.; et al. Development of genetic transformation and heterologous expression system in carboxydotrophic thermophilic acetogen Moorella thermoacetica. J. Biosci. Bioeng. 2013, 115, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Schiel-Bengelsdorf, B.; Dürre, P. Pathway engineering and synthetic biology using acetogens. FEBS Lett. 2012, 586, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Kamra, D.; Pawar, M.; Singh, B. Effect of plant secondary metabolites on rumen methanogens and methane emissions by ruminants. In Dietary Phytochemicals and Microbes; Patra, A.K., Ed.; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Wolin, M.; Miller, T.; Stewart, C. Microbe-microbe interactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; An Imprint of Chapman & Hall: London, UK, 1997. [Google Scholar]
- Cord-Ruwisch, R.; Seitz, H.-J.; Conrad, R. The capacity of hydrogenotrophic anaerobic bacteria to compete for traces of hydrogen depends on the redox potential of the terminal electron acceptor. Arch. Microbiol. 1988, 149, 350–357. [Google Scholar] [CrossRef]
- Joblin, K. Ruminal acetogens and their potential to lower ruminant methane emissions. Crop. Pasture Sci. 1999, 50, 1307–1314. [Google Scholar] [CrossRef]
- Fievez, V.; Mbanzamihigo, L.; Piattoni, F.; Demeyer, D. Evidence for reductive acetogenesis and its nutritional significance in ostrich hindgut as estimated from in vitro incubations. J. Anim. Physiol. Anim. Nutr. 2001, 85, 271–280. [Google Scholar] [CrossRef]
- Gagen, E.J.; Denman, S.E.; Padmanabha, J.; Zadbuke, S.; Al Jassim, R.; Morrison, M.; McSweeney, C.S. Functional gene analysis suggests different acetogen populations in the bovine rumen and tammar wallaby forestomach. Appl. Environ. Microb. 2010, 7, 7785–7795. [Google Scholar] [CrossRef] [PubMed]
- Gagen, E.J.; Mosoni, P.; Denman, S.E.; Al Jassim, R.; McSweeney, C.S.; Forano, E. Methanogen colonisation does not significantly alter acetogen diversity in lambs isolated 17 h after birth and raised aseptically. Microb. Ecol. 2012, 64, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Dansen, O.; Pellikaan, W.; Hendriks, W.; Dijkstra, J.; Jacobs, M.; Everts, H.; van Doorn, D. Daily methane production pattern of Welsh ponies fed a roughage diet with or without a cereal mixture. J. Anim. Sci. 2015, 93, 1916–1922. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.B. Methanogenesis in monogastric animals. Environ. Monit. Assess. 1996, 42, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Abecia, L.; Fondevila, M.; Rodríguez-Romero, N.; Martínez, G.; Yáñez-Ruiz, D. Comparative study of fermentation and methanogen community structure in the digestive tract of goats and rabbits. J. Anim. Physiol. Anim. Nutr. 2013, 97, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Fievez, V.; Piattoni, F.; Mbanzamihigo, L.; Demeyer, D. Reductive acetogenesis in the hindgut and attempts to its induction in the rumen-a review. J. Appl. Anim. Res. 1999, 16, 1–22. [Google Scholar] [CrossRef]
- Stevens, C.E.; Hume, I.D. Comparative Physiology of the Vertebrate Digestive System; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Leedle, J.; Greening, R. Postprandial changes in methanogenic and acidogenic bacteria in the rumens of steers fed high-or low-forage diets once daily. Appl. Environ. Microb. 1988, 54, 502–506. [Google Scholar]
- Demeyer, D.I.; Locquet, N.; De Graeve, K. Effect van Aminozuren op Hooifermentatie door Pens-en Caecuminhoud van Runderen, 18de Studiedag der Nederlandstalige Voedingsonderzoekers, Gent, 16 April 1993; Ghent University Library: Gent, Belgien, 1993. [Google Scholar]
- Sweeten, J.M. Concentrated Animal Feeding Operations in Texas: An Extension, Research and Educational Action Plan for Environmental Quality Management; Texas A&M University: College Station, TX, USA, 1994. [Google Scholar]
- Greening, R.; Leedle, J. Enrichment and isolation of Acetitomaculum ruminis, gen. nov., sp. nov.: Acetogenic bacteria from the bovine rumen. Arch. Microbiol. 1989, 151, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Bernalier, A.; Willems, A.; Leclerc, M.; Rochet, V.; Collins, M.D. Ruminococcus hydrogenotrophicus sp. nov., a new H2/CO2-utilizing acetogenic bacterium isolated from human feces. Arch. Microbiol. 1996, 166, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Rieu-Lesme, F.; Morvan, B.; Collins, M.; Fonty, G.; Willems, A. A new H2/CO2-using acetogenic bacterium from the rumen: Description of Ruminococcus schinkii sp. nov. FEMS Microbiol. Lett. 1996, 140, 281–286. [Google Scholar] [PubMed]
- Lorowitz, W.H.; Bryant, M.P. Peptostreptococcus productus strain that grows rapidly with CO as the energy source. Appl. Environ. Microbiol. 1984, 47, 961–964. [Google Scholar] [PubMed]
- Geerligs, G.; Aldrich, H.; Harder, W.; Diekert, G. Isolation and characterization of a carbon monoxide utilizing strain of the acetogen Peptostreptococcus productus. Arch. Microbiol. 1987, 148, 305–313. [Google Scholar] [CrossRef]
- Godoy-Vitorino, F.; Goldfarb, K.C.; Karaoz, U.; Leal, S.; Garcia-Amado, M.A.; Hugenholtz, P.; Tringe, S.G.; Brodie, E.L.; Dominguez-Bello, M.G. Comparative analyses of foregut and hindgut bacterial communities in hoatzins and cows. ISME J. 2012, 6, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Oh, P.L.; Benson, A.K.; Peterson, D.A.; Patil, P.B.; Moriyama, E.N.; Roos, S.; Walter, J. Diversification of the gut symbiont Lactobacillus reuteri as a result of host-driven evolution. ISME J. 2010, 4, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Schubert, A.M.; Zackular, J.P.; Iverson, K.D.; Young, V.B.; Petrosino, J.F. Stabilization of the murine gut microbiome following weaning. Gut Microbes 2012, 3, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Godwin, S.; Kang, A.; Gulino, L.-M.; Manefield, M.; Gutierrez-Zamora, M.-L.; Kienzle, M.; Ouwerkerk, D.; Dawson, K.; Klieve, A.V. Investigation of the microbial metabolism of carbon dioxide and hydrogen in the kangaroo foregut by stable isotope probing. ISME J. 2014, 8, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Breznak, J.A.; Blum, J.S. Mixotrophy in the termite gut acetogen, Sporomusa termitida. Arch. Microbiol. 1991, 156, 105–110. [Google Scholar] [CrossRef]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Breznak, J.A.; Kane, M.D. Microbial H2/CO2 acetogenesis in animal guts: Nature and nutritional significance. FEMS Microbiol. Rev. 1990, 7, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Liu, H.; Du, G.; Chen, J. Real-time PCR assays targeting formyltetrahydrofolate synthetase gene to enumerate acetogens in natural and engineered environments. Anaerobe 2009, 15, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Leaphart, A.B.; Lovell, C.R. Recovery and analysis of formyltetrahydrofolate synthetase gene sequences from natural populations of acetogenic bacteria. Appl. Environ. Microb. 2001, 67, 1392–1395. [Google Scholar] [CrossRef] [PubMed]
- Morvan, B.; Bonnemoy, F.; Fonty, G.; Gouet, P. Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and methanogenic archaea from digestive tract of different mammals. Curr. Microbiol. 1996, 32, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Hallenbeck, P.C.; Benemann, J.R. Biological hydrogen production; fundamentals and limiting processes. Int. J. Hydrog. Energy 2002, 27, 1185–1193. [Google Scholar] [CrossRef]
- Suzuki, T.; Matsuo, T.; Ohtaguchi, K.; Koide, K. Continuous production of acetic acid from CO2 in repeated-batch cultures using flocculated cells of Acetobacterium woodii. J. Chem. Eng. Jpn. 1993, 26, 459–462. [Google Scholar] [CrossRef]
- Balch, W.E.; Schoberth, S.; Tanner, R.S.; Wolfe, R. Acetobacterium, a new genus of hydrogen-oxidizing, carbon dioxide-reducing, anaerobic bacteria. Int. J. Syst. Evol. Microbiol. 1977, 27, 355–361. [Google Scholar] [CrossRef]
- Baba, S.; Tashiro, Y.; Shinto, H.; Sonomoto, K. Development of high-speed and highly efficient butanol production systems from butyric acid with high density of living cells of Clostridium saccharoperbutylacetonicum. J. Biotechnol. 2012, 157, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.W.; Yamazaki, H. Natural flavors production-a biotechnological approach. Trends Biotechnol. 1986, 4, 264–268. [Google Scholar] [CrossRef]
- Sharpell, F.H.J. Microbial flavors and fragrances. In Comprehensive Biotechnology; Blanch, H.W., Drew, S., Wang, D.I.C., Eds.; Pergamon Press: Oxford, UK, 1985. [Google Scholar]
- Playne, M.J. Propionic and butyric acids. In Comprehensive Biotechnology; Moo-Young, M., Ed.; Pergamon Press: Oxford, UK, 1985. [Google Scholar]
- Posey-Dowty, J.D.; Seo, K.S.; Walker, K.R.; Wilson, A.K. Carboxymethylcellulose acetate butyrate in water-based automotive paints. Surf. Coat. Int. B Coat. Trans. 2002, 85, 203–208. [Google Scholar] [CrossRef]
- Zidwick, M.J.; Chen, J.S.; Rogers, P. Organic acid and solvent production: Propionic and butyric acids and ethanol. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Álvarez-Chávez, C.R.; Edwards, S.; Moure-Eraso, R.; Geiser, K. Sustainability of bio-based plastics: General comparative analysis and recommendations for improvement. J. Clean. Prod. 2012, 23, 47–56. [Google Scholar] [CrossRef]
- Gonzalez-Garcia, R.A.; McCubbin, T.; Navone, L.; Stowers, C.; Nielsen, L.K.; Marcellin, E. Microbial propionic acid production. Fermentation 2017, 3, 21. [Google Scholar] [CrossRef]
- Worden, R.M.; Grethlein, A.J.; Zeikus, J.G.; Datta, D.R. Butyrate production from carbon monoxide by Butyribacterium methylotrophicum. Appl. Biochem. Biotechnol. 1989, 20–21, 687–698. [Google Scholar] [CrossRef]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Guan, L.L.; Liu, J.X.; Wang, J.K. Rumen fermentation and acetogen population changes in response to an exogenous acetogen TWA4 strain and Saccharomyces cerevisiae fermentation product. J. Zhejiang Univ. Sci. B 2015, 16, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucl. Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 22. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer Science & Business Media: Berlin, Deutschland, 2009. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Mächler, M.; Magnusson, A.; Möller, S. gplots: Various R Programming Tools for Plotting Data. 2009. Available online: https://www.researchgate.net/publication/303186599_gplots_Various_R_programming_tools_for_plottingdata (accessed on 6 June 2018).
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; Förster, W.; et al. ARB: A software environment for sequence data. Nucl. Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- McSweeney, C.S.; Denman, S.E.; Mackie, R.I. Rumen Bacteria. In Methods in Gut Microbial Ecology for Ruminants; Makkar, H.P.S., McSweeney, C.S., Eds.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Hungate, R.E. A roll tube method for cultivation of strict anaerobes. In Methods in Microbiology; Norris, J.R., Ribbons, D.W., Eds.; Academic Press: London, UK, 1969. [Google Scholar]
- Gagen, E.J.; Wang, J.; Padmanabha, J.; Liu, J.; de Carvalho, I.P.; Liu, J.; Webb, R.I.; Al Jassim, R.; Morrison, M.; Denman, S.E. Investigation of a new acetogen isolated from an enrichment of the tammar wallaby forestomach. BMC Microbiol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Propionate | Butyrate | Isovalerate |
|---|---|---|---|
| Rabbit | 6.86 b | 6.30 c | 0 d |
| Horse | 7.12 b | 7.02 a | 4.00 b |
| Cow | 8.38 a | 6.85 b | 3.49 c |
| Sheep | 8.59 a | 6.90 b | 4.17 a |
| SEM | 0.075 | 0.016 | 0.016 |
| Target | Forward/Reverse | Primer Sequences | Size (bp) |
|---|---|---|---|
| 16S rRNA of TB | F | CGGCAACGAGCGCAACCC | 130 |
| R | CCATTGTAGCACGTGTGTAGCC | ||
| fhs | F | GTWTGGGCWAARGGYGGMGAAGG | 250 |
| R | GTATTGDGTYTTRGCCATACA | ||
| mcrA | F | TTCGGTGGATCDCARAGRGC | 140 |
| R | GBARGTCGWAWCCGTAGAATCC | ||
| acsB | F | CTBTGYGGDGCIGTIWSMTGG | 216 |
| R | AARCAWCCRCADGADGTCATIGG |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C. Acetogen Communities in the Gut of Herbivores and Their Potential Role in Syngas Fermentation. Fermentation 2018, 4, 40. https://doi.org/10.3390/fermentation4020040
Yang C. Acetogen Communities in the Gut of Herbivores and Their Potential Role in Syngas Fermentation. Fermentation. 2018; 4(2):40. https://doi.org/10.3390/fermentation4020040
Chicago/Turabian StyleYang, Chunlei. 2018. "Acetogen Communities in the Gut of Herbivores and Their Potential Role in Syngas Fermentation" Fermentation 4, no. 2: 40. https://doi.org/10.3390/fermentation4020040
APA StyleYang, C. (2018). Acetogen Communities in the Gut of Herbivores and Their Potential Role in Syngas Fermentation. Fermentation, 4(2), 40. https://doi.org/10.3390/fermentation4020040