

Use of Trichoderma, Aspergillus, and Rhizopus Fungi for the Biological Production of Hydrolytic Enzymes and Uronic Acids from Sargassum Biomass

 ,
, 
 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Substrate Sampling and Conditioning
2.2. Fungal Inoculum Obtention
2.3. Solid-State Fermentation (SSF)
2.4. Enzyme Extraction
2.5. Sugar Characterization
2.6. Enzymatic Activity Determination
2.7. Sample Characterization
2.8. Phenolic Extraction and Determination
2.9. Statistical Analysis
3. Results
3.1. Characterization of Sargassum spp.
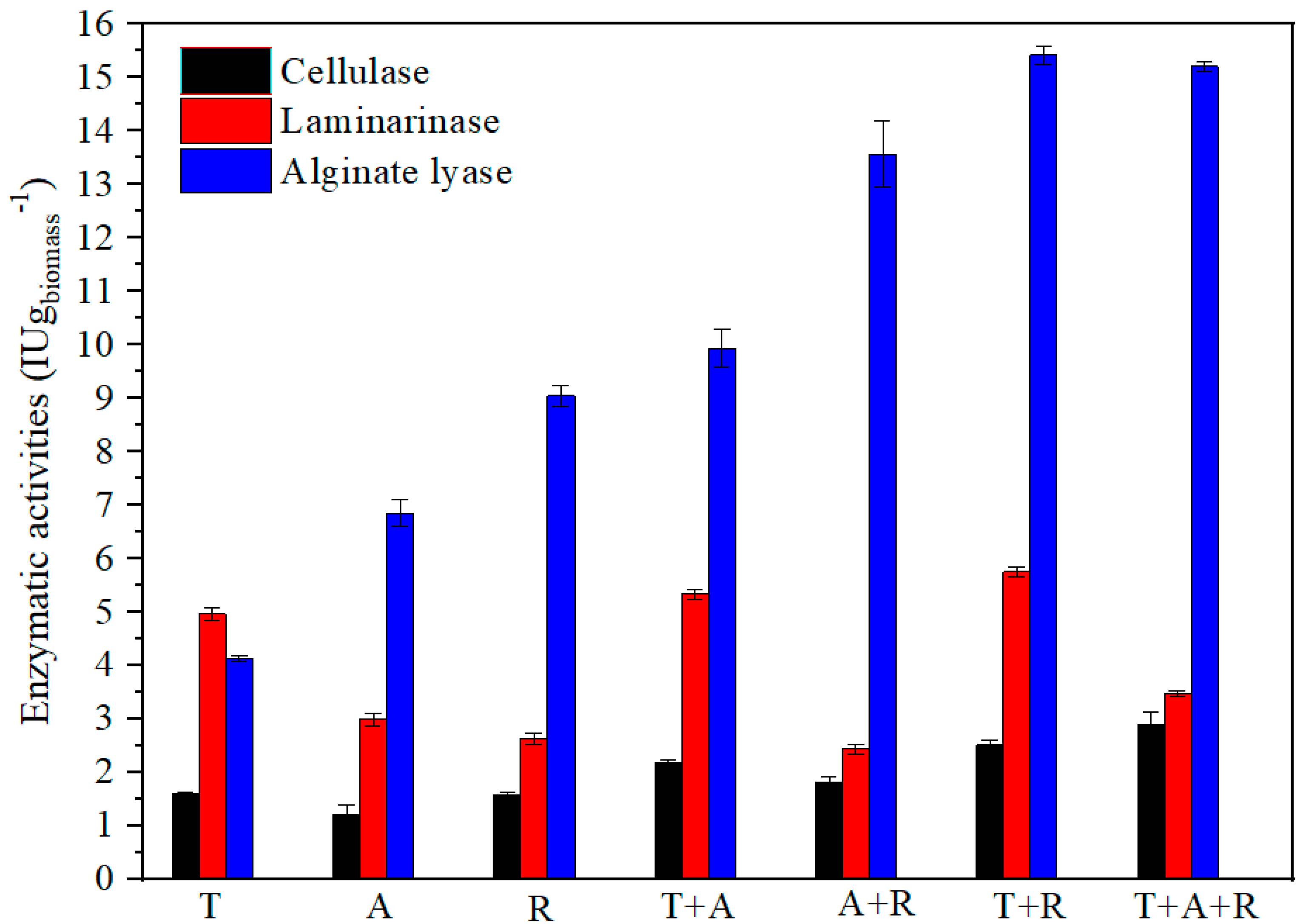
3.2. Enzymatic Production
3.3. Total Phenolic Compounds
3.4. Sugar Release
4. Discussion
4.1. Sargassum spp. As Sugary Biomass for Bio-Processes
4.2. Effect of Co-Culture in Enzymatic Production
4.3. Effect of Total Phenolic Compounds in Enzymatic Production
4.4. Fungal Co-Cultures Improve the Release of Sugars
4.5. SSF as a Feasible Alternative for Uronic Acid Production
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| A | Aspergillus oryzae |
| COD | Chemical Oxygen Demand |
| FPU | Filter paper unit |
| IU | International unit |
| R | Rhizopus oryzae |
| SSF | Solid-state fermentation |
| T | Trichoderma asperellum |
| TKN | Total Kjeldahl nitrogen |
| TPC | Total phenolic compounds |
| TRS | Total reducing sugars |
| TS | Total Solids |
| TVS | Volatile Solids |
References
- EUMOFA. Blue Bioeconomy Report; Luxembourg: European Commission. 2023. Available online: https://eumofa.eu/documents/20178/84590/blue+bioeconomy+report+2022+final.pdf/eb889d94-74a6-2c15-e136-4d2204118c6a?t=1673441855108 (accessed on 31 July 2024).
- Farobie, O.; Amrullah, A.; Bayu, A.; Syaftika, N.; Anis, L.A.; Hartulistiyoso, E. In-depth study of bio-oil and biochar production from macroalgae Sargassum sp. via slow pyrolysis. RSC Adv. 2022, 12, 9567–9578. [Google Scholar] [CrossRef] [PubMed]
- Marx, U.C.; Roles, J.; Hankamer, B. Sargassum blooms in the Atlantic Ocean—From a burden to an asset. Algal Res. 2021, 54, 102188. [Google Scholar] [CrossRef]
- Kostas, E.T.; Adams, J.M.M.; Ruiz, H.A.; Durán-Jiménez, G.; Lye, G.J. Macroalgal biorefinery concepts for the circular bioeconomy: A review on biotechnological developments and future perspective. Renew. Sustain. Energy Rev. 2021, 151, 111553. [Google Scholar] [CrossRef]
- González-Gloria, K.D.; Rodríguez-Jasso, R.M.; Aparicio, S.E.; Chávez González, M.L.; Kostas, E.T.; Ruiz, H.A. Macroalgal biomass in terms of third-generation biorefinery concept: Current status and techno-economic analysis. A review. Bioresour. Technol. Rep. 2016, 16, 100863. [Google Scholar] [CrossRef]
- Pereira, L. Non-indigenous seaweeds in the Iberian Peninsula, Macaronesia Islands (Madeira, Azores, Canary Islands) and Balearic Islands: Biodiversity, ecological impact, invasion dynamics, and potential industrial applications. Algal Res. 2024, 78, 103407. [Google Scholar] [CrossRef]
- González-Fernández, L.A.; Castillo Ramos, V.; Sánchez Polo, M.; Medellín Castillo, N.A. Fundamentals in applications of algae biomass: A review. J. Environ. Manag. 2023, 338, 117830. [Google Scholar] [CrossRef]
- Morales-Contreras, B.E.; Flórez-Fernández, N.; Torres, M.D.; Domínguez, H.; Rodríguez-Jasso, R.M.; Ruiz, H.A. Hydrothermal systems to obtain high value-added compounds from macroalgae for bioeconomy and biorefineries. Bioresour. Technol. 2022, 343, 126017. [Google Scholar] [CrossRef]
- Baghel, R.S. Developments in seaweed biorefinery research: A comprehensive review. Chem. Eng. J. 2023, 454, 140177. [Google Scholar] [CrossRef]
- Kwon, O.M.; Kim, D.H.; Kim, S.K.; Jeong, G.T. Production of sugars from macro-algae Gracilaria verrucosa using combined process of citric acid-catalyzed pretreatment and enzymatic hydrolysis. Algal Res. 2016, 13, 293–297. [Google Scholar] [CrossRef]
- García-Vaquero, M.; Rajauria, G.; O’Doherty, J.V.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef]
- Jumadi, O.; Annisi, A.D.; Djawad, Y.A.; Bourgougnon, N.; Amaliah, N.A.; Asmawati, A.; Baso Manguntungi, A.; Inubushi, K. Brown algae (Sargassum sp.) extract prepared by indigenous microbe fermentation enhanced tomato germination parameters. Biocatal. Agric. Biotechnol. 2023, 47, 102601. [Google Scholar] [CrossRef]
- Yuhendra, A.P.; Farghali, I.; Mohamed, M.A.; Iwasaki, M.; Tangtaweewipat, S.; Ihara, I.; Sakai, R.; Umetsu, K. Potential of biogas production from the anaerobic digestion of Sargassum fulvellum macroalgae: Influences of mechanical, chemical, and biological pretreatments. Biochem. Eng. J. 2021, 175, 108140. [Google Scholar] [CrossRef]
- Ramírez-Partida, A.E.; García-Cayuela, T.; Amador-Castro, L.F.; Alper, H.S.; Carrillo-Nieves, D. Towards a biorefinery processing Sargassum seaweed: Techno-economic assessment of alginate and fucoidan production through SuperPro Designer® process simulation. Environ. Technol. Innov. 2024, 34, 103587. [Google Scholar] [CrossRef]
- Gordillo-Sierra, A.R.G.; Amador-Castro, L.F.; Ramírez-Partida, A.E.; García-Cayuela, T.; Carrillo-Nieves, D.; Alper, H.S. Valorization of Caribbean Sargassum biomass as a source of alginate and sugars for de novo biodiesel production. J. Environ. Manag. 2022, 324, 116364. [Google Scholar] [CrossRef] [PubMed]
- Grand View Research Inc. Alginate Market Size, Share & Trends Analysis Report by Type (High M, High G), by Product (Sodium, Propylene Glycol), By Application (Pharmaceutical, Industrial), by Region, and Segment Forecasts, 2021–2028 (No. GVR-2-68038-244-0). 2021. Available online: https://www.grandviewresearch.com/industry-analysis/alginate-market. (accessed on 30 September 2024).
- Sari-Chmayssem, N.; Taha, S.; Mawlawi, H.; Guégan, J.P.; Jeftić, J.; Benvegnu, T. Extracted and depolymerized alginates from brown algae Sargassum vulgare of Lebanese origin: Chemical, rheological, and antioxidant properties. J. Appl. Phycol. 2016, 28, 1915–1929. [Google Scholar] [CrossRef]
- Mohammed, A.; Rivers, A.; Stuckey, D.C.; Ward, K. Alginate extraction from Sargassum seaweed in the Caribbean region: Optimization using response surface methodology. Carbohydr. Polym. 2020, 245, 116419. [Google Scholar] [CrossRef]
- Nogueira, M.T.; Chica, L.R.; Yamashita, C.; Nunes, N.S.S.; Moraes, I.C.F.; Branco, C.C.Z.; Branco, I.G. Optimal conditions for alkaline treatment of alginate extraction from the brown seaweed Sargassum cymosum C. Agardh by response surface methodology. Appl. Food Res. 2022, 2, 100141. [Google Scholar] [CrossRef]
- Youssouf, L.; Lallemand, L.; Giraud, P.; Soulé, F.; Bhaw-Luximon, A.; Meilhac, O.; Couprie, J. Ultrasound-assisted extraction and structural characterization by NMR of alginates and carrageenans from seaweeds. Carbohydr. Polym. 2017, 166, 55–63. [Google Scholar] [CrossRef]
- Yuan, Y.; Macquarrie, D.J. Microwave assisted step-by-step process for the production of fucoidan, alginate sodium, sugars and biochar from Ascophyllum nodosum through a biorefinery concept. Bioresour. Technol. 2015, 198, 819–827. [Google Scholar] [CrossRef]
- Gravdahl, M.; Aarstad, O.; Petersen, A.; Karlsen, S.; Donati, I.; Czjzek, M.; Alexander, O.; Åstrand, H.; Rye, P.D.; Tøndervik, A.; et al. A Chemo-Enzymatic Method for Preparation of Saturated Oligosaccharides from Alginate and Other Uronic Acid-Containing Polysaccharides. Carbohydr. Polym. 2024, 343, 122487. [Google Scholar] [CrossRef]
- Chen, C.; Li, X.; Lu, C.; Zhou, X.; Chen, L.; Qiu, C.; Long, J.; Qiu, C.; Jin, Z.; Long, J. Advances in alginate lyases and the potential application of enzymatic prepared alginate oligosaccharides: A mini review. Int. J. Biol. Macromol. 2024, 260 Pt 1, 129506. [Google Scholar] [CrossRef] [PubMed]
- Agabo-García, C.; Romero-García, L.I.; Álvarez-Gallego, C.J.; Blandino, A. Valorisation of the invasive alga Rugulopteryx okamurae through the production of monomeric sugars. Appl. Microbiol. Biotechnol. 2023, 107, 1971–1982. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Liu, C.; Zhang, C.; He, X.; Wang, H.; Peng, W.; Zheng, C. Trichoderma species from plant and soil: An excellent resource for biosynthesis of terpenoids with versatile bioactivities. J. Adv. Res. 2023, 49, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Salwan, R.; Sharma, V. Extracellular proteins of Trichoderma and their role in plant health. S. Afr. J. Bot. 2022, 147, 359–369. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, L.; Yu, D.; Lin, H.; Shen, Q.; Zhao, Y. Excellent waste biomass-degrading performance of Trichoderma asperellum T-1 during submerged fermentation. Sci. Total Environ. 2017, 31, 1329–1339. [Google Scholar] [CrossRef]
- Zheng, F.; Han, T.; Basit, A.; Liu, J.; Miao, T.; Jiang, W. Whole-genome sequence and comparative analysis of Trichoderma asperellum ND-1 reveal its unique enzymatic system for efficient biomass degradation. Catalysts 2022, 12, 437. [Google Scholar] [CrossRef]
- Vargas-Hoyos, H.A.; Gilchrist-Ramelli, E. Producción de enzimas hidrolíticas y actividad antagónica de Trichoderma asperellum sobre dos cepas de Fusarium aisladas de cultivos de tomate (Solanum lycopersicum). Sci. Fungorum 2015, 42, 9–16. [Google Scholar]
- Agabo-García, C.; Sarafidou, M.; Psaki, O.; Argeiti, C.; Blandino, A.; Álvarez-Gallego, C.; Romero-García, L.I.; Koutinas, A. Bioprocess development for the production of poly (3-hydroxybutyrate) using the invasive macroalgae Rugulopteryx okamurae. Environ. Technol. Innov. 2025, 40, 104342. [Google Scholar] [CrossRef]
- Romero-Vargas, A.; Cala, K.; Blandino, A.; Díaz, A.B. Bioconversion of the invasive seaweed Rugulopteryx okamurae into enzymes and polyhydroxyalkanoates. Algal Res. 2024, 81, 103587. [Google Scholar] [CrossRef]
- Sun, Z.; Wu, Y.; Long, S.; Feng, S.; Jia, X.; Hu, Y.; Maomao, M.; Jingxin, L.; Zeng, B. Aspergillus oryzae as a cell factory: Research and applications in industrial production. J. Fungi 2024, 10, 248. [Google Scholar] [CrossRef]
- Londoño-Hernández, L.; Ramírez-Toro, C.; Ruiz, H.A.; Ascacio-Valdés, J.A.; Aguilar-Gonzalez, M.A.; Rodríguez-Herrera, R.; Aguilar, C.N. Rhizopus oryzae–Ancient microbial resource with importance in modern food industry. Int. J. Food Microbiol. 2017, 257, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, E.; Benoit, I.; van den Brink, J.; Wiebenga, A.; Coutinho, P.M.; Henrissat, B.; de Vries, R.P. Carbohydrate-active enzymes from the zygomycete fungus Rhizopus oryzae: A highly specialized approach to carbohydrate degradation depicted at genome level. BMC Genom. 2011, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Vellozo-Echevarría, T.; Barrett, K.; Vuillemin, M.; Meyer, A.S. Mini-Review: The distinct carbohydrate active enzyme secretome of Rhizopus spp. represents fitness for mycelium remodeling and solid-state plant food fermentation. ACS Omega 2024, 9, 34185–34195. [Google Scholar] [CrossRef] [PubMed]
- Villasante, J.; Espinosa-Ramírez, J.; Pérez-Carrillo, E.; Heredia-Olea, E.; Almajano, M. Extrusion and solid-state fermentation with Aspergillus oryzae on the phenolic compounds and radical scavenging activity of pecan nut (Carya illinoinensis) shell. Br. Food J. 2021, 123, 4367–4382. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Koteshwara, A.; Philip, N.V.; Aranjani, J.M.; Hariharapura, R.C.; Mallikarjuna, S.V. A set of simple methods for detection and extraction of laminarinase. Sci. Rep. 2021, 11, 2489. [Google Scholar] [CrossRef]
- APHA-AWWA-WPCF. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Wu, Y.; Gao, H.; Wang, Y.; Peng, Z.; Guo, Z.; Ma, Y.; Zhang, R.; Zhang, M.; Wu, Q.; Xiao, J.; et al. Effects of different extraction methods on contents, profiles, and antioxidant abilities of free and bound phenolics of Sargassum polycystum from the South China Sea. J. Food Sci. 2022, 87, 968–981. [Google Scholar] [CrossRef]
- Putra, V.G.P.; Mutiarahma, S.; Chaniago, W.; Rahmadi, P.; Kurnianto, D.; Hidayat, C.; Carrera, C.; Palma, M.; Setyaningsih, W. An ultrasound-based technique for the analytical extraction of phenolic compounds in red algae. Arab. J. Chem. 2022, 15, 103597. [Google Scholar] [CrossRef]
- Espada-Bellido, E.; Ferreiro-González, M.; Carrera, C.; Palma, M.; Barroso, G.G.; Barbero, C.F. Optimization of the ultrasound-assisted extraction of anthocyanins and total phenolic compounds in mulberry (Morus nigra) pulp. Food Chem. 2017, 15, 23–32. [Google Scholar] [CrossRef]
- Davis, D.; Simister, R.; Campbell, S.; Marston, M.; Bose, S.; McQueen-Mason, S.J.; Gomez, L.D.; Gallimore, W.A.; Tonon, T. Biomass composition of the golden tide pelagic seaweeds Sargassum fluitans and S. natans (morphotypes I and VIII) to inform valorisation pathways. Sci. Total Environ. 2021, 762, 143134. [Google Scholar] [CrossRef]
- Tonon, T.; Machado, C.B.; Webber, M.; Webber, D.; Smith, J.; Pilsbury, A.; Allen, M.J. Biochemical and elemental composition of pelagic Sargassum biomass harvested across the Caribbean. Phycology 2022, 2, 204–215. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, W.; Zeng, W.; Chen, R.; Lin, D.; Li, G.; Liang, H. Bacterial-algae biofilm enhance MABR adapting a wider COD/N ratios wastewater: Performance and mechanism. Sci. Total Environ. 2021, 781, 146663. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, M.R.; Xia, A.; Murphy, J.D. The effect of seasonal variation on biomethane production from seaweed and on application as a gaseous transport biofuel. Bioresour. Technol. 2016, 209, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Li, Q.; Han, X.; Du, Y.; Jiang, Y.; Yan, X.; Cui, Y.; Kang, W.; Meng, L.; Cao, Z. Unleashing the potential of short-chain fatty acids: Alginate-degrading consortium boosts anaerobic sludge fermentation. J. Environ. Chem. Eng. 2024, 12, 112178. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Narsico, J.; Kobayashi, T.; Inoue, A.; Ojima, T. Production of poly (3-hydroyxybutylate) by a novel alginolytic bacterium Hydrogenophaga sp. strain UMI-18, using alginate as a sole carbon source. J. Biosci. Bioeng. 2019, 128, 203–208. [Google Scholar] [CrossRef]
- Fagundo-Mollineda, A.; Robledo, D.; Vásquez-Elizondo, R.M.; Freile-Pelegrín, Y. Antioxidant activities in holopelagic Sargassum species from the Mexican Caribbean: Temporal changes and intra-thallus variation. Algal Res. 2023, 76, 103289. [Google Scholar] [CrossRef]
- Fernández-Medina, P.; Álvarez-Gallego, C.J.; Caro, I. Yield evaluation of enzyme hydrolysis and dark fermentation of the brown seaweed Rugulopteryx okamurae hydrothermally pretreated by microwave irradiation. J. Environ. Chem. Eng. 2022, 10, 108817. [Google Scholar] [CrossRef]
- Sharma, P.; Melkania, U. Impact of furan derivatives and phenolic compounds on hydrogen production from the organic fraction of municipal solid waste using co-culture of Enterobacter aerogenes and E. coli. Bioresour. Technol. 2017, 239, 49–56. [Google Scholar] [CrossRef]
- Fernandes-Qualhato, T.; Alvares Cardoso Lopes, F.; Stecca Steindorff, A.; Silva Brandão, R.; Santos, R.; Jesuino, A.; Ulhoa, C.J. Mycoparasitism studies of Trichoderma species against three phytopathogenic fungi: Evaluation of antagonism and hydrolytic enzyme production. Biotechnol. Lett. 2013, 35, 1461–1468. [Google Scholar] [CrossRef]
- Erpel, F.; Mateos, R.; Pérez-Jiménez, J.; Pérez-Correa, J.R. Phlorotannins: From isolation and structural characterization, to the evaluation of their antidiabetic and anticancer potential. Food Res. Int. 2020, 137, 109589. [Google Scholar] [CrossRef]
- Borisova, A.S.; Pihlajaniemi, V.; Kont, R.; Niemelä, K.; Koitto, T.; Mikkelson, A.; Väljamäe, P.; Kristiina Kruus, K.; Marjamaa, K. The effect of soluble phenolic compounds from hydrothermally pretreated wheat straw on Trichoderma reesei cellulases and commercial enzyme cocktails. Biomass Convers. Biorefinery 2024, 14, 971–984. [Google Scholar] [CrossRef]
- Ximenes, E.; Kim, Y.; Mosier, N.; Dien, B.; Ladisch, M. Deactivation of cellulases by phenols. Enzyme Microb. Technol. 2011, 48, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Waghmode, A.V.; Khilare, C.J. RP-HPLC profile of major phenolics from brown marine macro algae. J. Appl. Pharm. 2018, 10, 1–5. [Google Scholar] [CrossRef]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid glucosides are hydrolyzed and thus activated in the oral cavity in humans. J. Nutr. 2005, 135, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Kashegari, A.T.; Shalabi, A.A.; Darwish, K.M.; El-Halawany, A.M.; Algandaby, M.M.; Ibrahim, S.R.M.; Abdel-Naim, A.B.; Koshak, A.E.; Mohamed, G.; et al. Phenolics from Chrozophora oblongifolia aerial parts as inhibitors of α-glucosidases and advanced glycation end products: In-vitro assessment, molecular docking and dynamics studies. Biology 2022, 11, 762. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol 2020, 10, 604923. [Google Scholar] [CrossRef]
- Vaghela, P.; Trivedi, K.; Anand, K.V.; Brahmbhatt, H.; Nayak, J.; Khandhediya, K.; Ghosh, A. Scientific basis for the use of minimally processed homogenates of Kappaphycus alvarezii (red) and Sargassum wightii (brown) seaweeds as crop biostimulants. Algal Res. 2023, 70, 102969. [Google Scholar] [CrossRef]
- Darko, C.N.S.; Premarathna, A.D.; Humayun, S.; Agyei-Tuffour, B.; Goosen, N.J.; Tuvikene, R. Physico- and biochemical properties of alginates extracted from Ecklonia maxima and Sargassum fluitans using a simple cascade process. J. Appl. Phycol. 2024, 36, 661–674. [Google Scholar] [CrossRef]
- Peteiro, C. Alginate Production from Marine Macroalgae, with Emphasis on Kelp Farming. In Alginates and Their Biomedical Applications; Series in Biomaterials Science and Engineering; Rehm, B., Moradali, M., Eds.; Springer: Singapore, 2018; Chapter 2; pp. 27–66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Enzymatic Activities | Cellulase (FPU gbiomass−1) | Laminarinase (IU gbiomass−1) | Alginate Lyase (IU gbiomass−1) | TRS (g L−1) |
|---|---|---|---|---|
| t | e/t | T | e/t | |
| T + A | 1.40 | 1.55 | 2.97 | 1.46 |
| A + R | 1.40 | 1.30 | 1.81 | 0.79 |
| T + R | 1.59 | 1.58 | 2.80 | 1.70 |
| T + A + R | 1.45 | 1.99 | 2.50 | 0.99 |
| Samples | TPC mg L−1 (Liquid Extract) | TPC mg L−1 (Solid After Extraction) | TPC Liquid/Solid Ratio | %TPCresidual (Dry Solid Biomass) |
|---|---|---|---|---|
| T | 50.4 ± 0.10 | 62.0 ± 2.18 | 0.81 | 16.9 ± 4.23 |
| A | 37.8 ± 4.00 | 44.3 ± 0.10 | 0.85 | 12.1 ± 3.63 |
| R | 47.9 ± 1.00 | 54.9 ± 0.10 | 0.87 | 15.0 ± 3.63 |
| T + A | 45.3 ± 0.73 | 60.8 ± 0.36 | 0.75 | 16.6 ± 3.65 |
| A + R | 41.4 ± 1.31 | 79.2 ± 0.10 | 0.52 | 21.6 ± 3.63 |
| T + R | 52.4 ± 2.40 | 67.6 ± 0.54 | 0.77 | 18.5 ± 3.67 |
| T + A + R | 46.6 ± 2.18 | 84.1 ± 0.09 | 0.55 | 22.9 ± 3.62 |
| Samples | TPC/FPU (mgTPC FPU−1) | TPC/IULaminarinase (mgTPC IU−1) | TPC/IUAlginate lyase (mgTPC IU−1) |
|---|---|---|---|
| T | 0.63 | 0.25 | 0.47 |
| A | 0.62 | 0.38 | 0.16 |
| R | 0.61 | 0.59 | 0.14 |
| T + A | 0.42 | 0.21 | 0.11 |
| A + R | 0.46 | 0.58 | 0.07 |
| T + R | 0.42 | 0.22 | 0.08 |
| T + A + R | 0.32 | 0.38 | 0.07 |
| Uronic Acids (mg gbiomass−1) | Method Extraction | Reference |
|---|---|---|
| 6.19 ± 0.55 | Commercial enzyme (215 μL of Cellic2® CTec2) + 30% S-L (algae–acetate buffer). Incubation: 50 °C; 24 h | [43] |
| 4.67 ± 0.40 | Commercial enzyme (150 μL of amyloglucosidase) + 30% S-L (algae–potassium phosphate buffer). Incubation: 25 °C; 24 h | [43] |
| 11.7 ± 1.1 | Commercial enzyme (100 μL of pronase) + 30% S-L (algae–potassium phosphate buffer) + Incubation: 40 °C; 24 h | [43] |
| 8.67 ± 0.73 | Commercial enzyme (50 μL laminarinase) + 30% S-L (algae–HEPES buffer). Incubation: 90 °C; 24 h | [43] |
| 7.88 ± 0.40 | Commercial enzyme (50 μL alginate lyase) + 30% S-L (algae–HEPES buffer). Incubation: 90 °C; 24 h | [43] |
| 14.4 ± 0.0 | Enzymes from SSF (A + R) + 5% S-L (Algae-Tween 80). Incubation: 4 °C, 30 min, 150 rpm | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agabo-García, C.; Cahyanto, M.N.; Setyaningsih, W.; Romero-García, L.I.; Álvarez-Gallego, C.J.; Blandino, A. Use of Trichoderma, Aspergillus, and Rhizopus Fungi for the Biological Production of Hydrolytic Enzymes and Uronic Acids from Sargassum Biomass. Fermentation 2025, 11, 430. https://doi.org/10.3390/fermentation11080430
Agabo-García C, Cahyanto MN, Setyaningsih W, Romero-García LI, Álvarez-Gallego CJ, Blandino A. Use of Trichoderma, Aspergillus, and Rhizopus Fungi for the Biological Production of Hydrolytic Enzymes and Uronic Acids from Sargassum Biomass. Fermentation. 2025; 11(8):430. https://doi.org/10.3390/fermentation11080430
Chicago/Turabian StyleAgabo-García, Cristina, Muhammad Nur Cahyanto, Widiastuti Setyaningsih, Luis I. Romero-García, Carlos J. Álvarez-Gallego, and Ana Blandino. 2025. "Use of Trichoderma, Aspergillus, and Rhizopus Fungi for the Biological Production of Hydrolytic Enzymes and Uronic Acids from Sargassum Biomass" Fermentation 11, no. 8: 430. https://doi.org/10.3390/fermentation11080430
APA StyleAgabo-García, C., Cahyanto, M. N., Setyaningsih, W., Romero-García, L. I., Álvarez-Gallego, C. J., & Blandino, A. (2025). Use of Trichoderma, Aspergillus, and Rhizopus Fungi for the Biological Production of Hydrolytic Enzymes and Uronic Acids from Sargassum Biomass. Fermentation, 11(8), 430. https://doi.org/10.3390/fermentation11080430