
Unlocking the Potential of Lacticaseibacillus rhamnosus 73 as a Ripening Agent in Semi-Hard Cheese After Freeze-Drying and a Six-Month Storage Period


Abstract
1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Biomass Production, Freeze-Drying, and Storage
2.3. Analysis of the Freeze-Dried Samples
2.4. Cultured Milk
2.5. Cheesemaking
2.6. Cheese Composition, pH Determination, and Microbiological Counts
2.7. Carbohydrates and Organic Acids of Cultured Milk and Cheese
2.8. Peptide Profiles of the Soluble Fraction of Cheese
2.9. Statistical Analyses
3. Results and Discussion
3.1. Microbiological Counts of Freeze-Dried Cultures
3.2. Growth and Activity in Cultured Milk
3.3. Cheese Composition, pH Values, and Microbiological Counts
3.4. Carbohydrates and Organic Acids in Cheese
3.5. Peptide Profiling of Cheese
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gänzle, M.G.; Monnin, L.; Zheng, J.; Zhang, L.; Coton, M.; Sicard, D.; Walter, J. Starter culture development and innovation for novel fermented foods. Annu. Rev. Food Sci. Technol. 2024, 15, 211–239. [Google Scholar] [CrossRef] [PubMed]
- Wouters, J.T.M.; Ayad, E.H.E.; Hugenholtz, J.; Smit, G. Microbes from raw milk for fermented dairy products. Int. Dairy J. 2002, 12, 91–109. [Google Scholar] [CrossRef]
- Bintsis, T.; Papademas, P. The application of protective cultures in cheese: A review. Fermentation 2024, 10, 117. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Ciocia, F.; Mcsweeney, P.L.H.; Piraino, P.; Parente, E. Use of dairy and non-dairy Lactobacillus plantarum, Lactobacillus paraplantarum and Lactobacillus pentosus strains as adjuncts in cheddar cheese. Dairy Sci. Technol. 2015, 93, 623–640. [Google Scholar] [CrossRef]
- Peighambardoust, S.H. Application of spray drying for preservation of lactic acid starter cultures: A review. Trends Food Sci. Technol. 2011, 22, 215–224. [Google Scholar] [CrossRef]
- Aschenbrenner, M.; Foerst, P.; Kulozik, U. Freeze-drying of probiotics. In Advances in Probiotic Technology; Foerst, P., Santivarangkna, C., Eds.; CRC Press: Boca Ratón, FL, USA, 2015; pp. 213–241. [Google Scholar] [CrossRef]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 2004, 14, 835–847. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Q.; Chen, W.; Du, G.; Chen, J. Short communication: Protection of lyophilized milk starter Lactobacillus casei Zhang by glutathione. J. Dairy Sci. 2016, 99, 1846–1852. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Binetti, A.; Burns, P.; Cuatrin, A.; Reinheimer, J.; Salminen, S.; Vinderola, G. Occurrence of bacteria with technological and probiotic potential in Argentinian human breast-milk. Benef. Microbes 2020, 11, 685–702. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X. Survival of freeze-dried Lactobacillus plantarum and Lactobacillus rhamnosus during storage in the presence of protectants. Biotechnol. Lett. 2002, 24, 1587–1591. [Google Scholar] [CrossRef]
- Oluwatosin, S.O.; Tai, S.L.; Fagan-endres, M.A. Sucrose, maltodextrin and inulin efficacy as cryoprotectant, preservative and prebiotic—Towards a freeze dried Lactobacillus plantarum topical probiotic. Biotechnol. Rep. 2022, 33, e00696. [Google Scholar] [CrossRef] [PubMed]
- Tyutkov, N.; Zhernyakova, A.; Birchenko, A.; Eminova, E.; Nadtochii, L.; Baranenko, D. Probiotics viability in frozen food products. Food Biosci. 2022, 50, 101996. [Google Scholar] [CrossRef]
- Ugarte, M.B.; Guglielmotti, D.; Giraffa, G.; Reinheimer, J.; Hynes, E. Nonstarter lactobacilli isolated from soft and semi-hard argentinean cheeses: Genetic characterization and resistance to biological barriers. J. Food Prot. 2006, 69, 2983–2991. [Google Scholar] [CrossRef] [PubMed]
- Batistela, M.E.; Bergamini, C.V.; Ale, E.C.; Renzo, S.; Peralta, G.H. Culture medium based on whey permeate for biomass production of lactobacilli with technological aptitudes. Food Biosci. 2024, 62, 105445. [Google Scholar] [CrossRef]
- Peralta, G.H.; Bergamini, C.V.; Hynes, E.R. Aminotransferase and glutamate dehydrogenase activities in lactobacilli and streptococci. Braz. J. Microbiol. 2016, 47, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Peralta, G.H.; Bergamini, C.V. Mini-cheese models. In Dairy Foods Processing; Gomes da Cruz, A., Colombo Pimentel, T., Esmerino, E.A., Verruck, S., Eds.; Methods and Protocols in Food Science; Humana: New York, NY, USA, 2025; pp. 151–162. [Google Scholar] [CrossRef]
- ISO 5534:2004; Cheese and Processed Cheese: Determination of the Total Solids Content (5534/IDF 4, Reference method). International Organisation for Standardisation: Geneva, Switzerland, 2004.
- ISO 3433:2008; Cheese: Determination of Fat Content: Van Gulik Method. (3433/IDF 222). International Organisation for Standardisation: Geneva, Switzerland, 2018.
- ISO 8968-1:2014; Milk and Milk Products: Determination of Nitrogen Content: Part 1: Kjeldahl Principle and Crude Protein Calculation. (8968/IDF 20). International Organisation for Standardisation: Geneva, Switzerland, 2014.
- Ardö, Y.; Polychroniadou, A. Laboratory Manual for Chemical Analysis Of Cheese: Improvement of the Quality of the Production of Raw Milk Cheeses; Publications Office: Luxembourg, 1999. [Google Scholar]
- Giménez, P.; Peralta, G.H.; Wolf, I.V.; Hynes, E.R.; Bergamini, C.V. Adjunct cultures of autochthonous Lactobacillus strains to diversify and improve cheese maturation. Int. J. Dairy Technol. 2014, 78, 13111. [Google Scholar] [CrossRef]
- Giménez, P.; Peralta, G.H.; Batistela, M.E.; George, G.A.; Ale, E.C.; Quintero, J.P.; Hynes, E.R.; Bergamini, C.V. Impact of the use of skim milk powder and adjunct cultures on the composition, yield, proteolysis, texture and melting properties of Cremoso cheese. Int. Dairy J. 2023, 140, 105595. [Google Scholar] [CrossRef]
- Milesi, M.; Vinderola, G.; Sabbag, N.; Meinardi, C.; Hynes, E. Influence on cheese proteolysis and sensory characteristics of non-starter lactobacilli strains with probiotic potential. Food Res. Int. 2009, 42, 1186–1196. [Google Scholar] [CrossRef]
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Spigelman, A.F.; MacDonald, P.E.; Wishart, D.S.; Li, S.; et al. MetaboAnalyst 6.0: Towards a unified platform for metabolomics data processing, analysis and interpretation. Nucleic Acids Res. 2024, 52, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Champagne, C.P.; Gardner, N.; Brochu, E.; Beaulieu, Y. The freeze-drying of lactic acid bacteria. a review. Can. Inst. Food Sci. Technol. J. 1991, 24, 118–128. [Google Scholar] [CrossRef]
- Morgan, C.A.; Herman, N.; White, P.A.; Vesey, G. Preservation of microorganisms by drying; a review. J. Microbiol. Methods 2006, 66, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.; Girardeau, A.; Passot, S. Freeze-drying of lactic acid bacteria: A stepwise approach for developing a freeze-drying protocol based on physical properties. In Cryopreservation and Freeze-Drying Protocols; Wolkers , W.F., Oldenhof, H., Eds.; Springer Nature: New York, NY, USA, 2021; pp. 703–720. [Google Scholar]
- Senovieski, M.L.; Loyeau, P.A.; Puntillo, M.A.; Binetti, A.G.; Vinderola, C.G. Cheese whey for production of breast milk-derived bifidobacteria: Influence of fermentation conditions on the survival to spray drying and storage. Int. J. Dairy Technol. 2024, 77, 853–861. [Google Scholar] [CrossRef]
- Lee, S.; Kim, D.; Park, H. Effects of protectant and rehydration conditions on the survival rate and malolactic fermentation efficiency of freeze-dried Lactobacillus plantarum JH287. Appl. Microbiol. Biotechnol. 2016, 100, 7853–7863. [Google Scholar] [CrossRef] [PubMed]
- Bai, S. Enhancing the Stability of Probiotics: Freeze-Drying and Encapsulation. Doctoral Thesis, Lund University, Lund, Sweden, 2024. [Google Scholar]
- Lodato, P.; Segovia de Huergo, M.; Buera, M.P. Viability and thermal stability of a strain of Saccharomyces cerevisiae freeze-dried in different sugar and polymer matrices. Appl. Microbiol. Biotechnol. 1999, 52, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Santivarangkna, C.Ã.; Higl, B.; Foerst, P. Protection mechanisms of sugars during different stages of preparation process of dried lactic acid starter cultures. Food Microbiol. 2008, 25, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Li, Y.; Kang, W.; Han, Y.; Yin, B.; Yang, R.; Tang, R.; Pan, L.; Wang, J.; Li, W.; et al. Synergistic combination of cryoprotectants improved freeze-dried survival rate and viable counts of Lactiplantibacillus plantarum. Int. J. Dairy Technol. 2024, 77, 348–357. [Google Scholar] [CrossRef]
- Silva, J.; Silva, J.; Sofia, A.; Helena, C.; Teixeira, P.; Gibbs, P.A. Induction of stress tolerance in Lactobacillus delbrueckii ssp. bulgaricus by the addition of sucrose to the growth medium. J. Dairy Res. 2004, 71, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Gaudreau, H.; Champagne, C.P.; Jelen, P. The use of crude cellular extracts of Lactobacillus delbrueckii ssp. bulgaricus 11842 to stimulate growth of a probiotic Lactobacillus rhamnosus culture in milk. Enzym. Microb. Technol. 2005, 36, 83–90. [Google Scholar] [CrossRef]
- Shah, N. Probiotic bacteria: Selective enumeration and survival in dairy foods. J. Dairy Sci. 2000, 83, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, H.; Qiao, Y.; Liu, G.; Leng, C.; Zhang, Y.; Lv, X.; Feng, Z. The nutrient requirements of Lactobacillus rhamnosus GG and their application to fermented milk. J. Dairy Sci. 2019, 102, 5971–5978. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Christine, K.W.; Shah, N.P. Towards galactose accumulation in dairy foods fermented by conventional starter cultures: Challenges and strategies. Trends Food Sci. Technol. 2015, 41, 24–36. [Google Scholar] [CrossRef]
- Tormo, M.; Izco, J.M. Alternative reversed-phase high-performance liquid chromatography method to analyse organic acids in dairy products. J. Chromatogr. A 2004, 1033, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.; Winter, C.K. Diacetyl in foods: A review of safety and sensory characteristics. Compr. Rev. Food Sci. Food Saf. 2015, 14, 634–643. [Google Scholar] [CrossRef]
- Calasso, M.; Gobbetti, M. Lactic acid bacteria|Lactobacillus spp.: Other species. In Encyclopedia of Dairy Sciences; Elsevier: Amsterdam, The Netherlands, 2011; pp. 125–131. [Google Scholar] [CrossRef]
- Hatti-kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef] [PubMed]
- Prada, S. Uso de Homogeneización de Alta Presión y Lisozima Para Mejorar la Actividad Metabólica de Lactiplantibacillus plantarum 29 en Queso Semiduro sin sal Añadida. Bachelor’s Thesis, Universidad Nacional del Litoral, Santa Fe, Argentina, 2025. [Google Scholar]
- Johnson, M.E. Mesophilic and thermophilic cultures used in traditional cheesemaking. Microbiol. Spectr. 2013, 1, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Kindstedt, P.S. The Basics of cheesemaking. Microbiol. Spectr. 2013, 1, 17–38. [Google Scholar] [CrossRef] [PubMed]
- Galat, A.; Dufresne, J.; Combrisson, J.; Thépaut, J.; Boumghar-Bourtchai, L.; Boyer, M.; Fourmestraux, C. Novel method based on chromogenic media for discrimination and selective enumeration of lactic acid bacteria in fermented milk products. Food Microbiol. 2016, 55, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Ur-Rehman, S.; Fox, P.F.; McSweeney, P.L.H.; Madkor, S.A.; Farkye, N.Y. Alternatives to pilot plant experiments in cheese-ripening studies. Int. J. Dairy Technol. 2001, 54, 121–126. [Google Scholar] [CrossRef]
- Hutkins, R.; Halambeck, S.M.; Morris, H.A. Use of galactose-fermenting Streptococcus thermophilus in the manufacture of Swiss, mozzarella, and short-method cheddar cheese. J. Dairy Sci. 1986, 69, 1–8. [Google Scholar] [CrossRef]
- Green, I.R.; Oberg, C.J.; McMahon, D.J. Galactose-positive adjunct cultures prevent gas formation by Paucilactobacillus wasatchensis WDC04 in a model gas production test. J. Dairy Sci. 2021, 104, 10540–10549. [Google Scholar] [CrossRef] [PubMed]
- Ortakci, F.; Broadbent, J.R.; Oberg, C.J.; McMahon, D.J. Growth and gas production of a novel obligatory heterofermentative Cheddar cheese nonstarter lactobacilli species on ribose and galactose. J. Dairy Sci. 2015, 98, 3645–3654. [Google Scholar] [CrossRef] [PubMed]
- Ortakci, F.; Broadbent, J.R.; Oberg, C.J.; McMahon, D.J. Late blowing of Cheddar cheese induced by accelerated ripening and ribose and galactose supplementation in presence of a novel obligatory heterofermentative nonstarter Lactobacillus wasatchensis. J. Dairy Sci. 2015, 98, 7460–7472. [Google Scholar] [CrossRef] [PubMed]
- Neviani, E.; Gatti, M.; Gardini, F.; Levante, A. Microbiota of cheese ecosystems: A perspective on cheesemaking. Foods 2025, 14, 830. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, V.K.; McSweeney, P.L.H.; Magboul, A.A.A.; Fox, P.F. Proteolysis in cheese during ripening. In Cheese: Chemistry, Physics and Microbiology; Elsevier: Amsterdam, The Netherlands, 2004; pp. 391–433. [Google Scholar] [CrossRef]
- Fox, P.F.; Singh, T.K.; McSweeney, P.L.H. Proteolysis in cheese during ripening. In Biochemistry of Milk Products; Andrews, A.T., Varley, J., Eds.; Woodhead Publishing: Sawston, UK, 2005; pp. 1–31. [Google Scholar] [CrossRef]
- Ardö, Y.; McSweeney, P.L.H.; Magboul, A.A.A.; Upadhyay, V.K.; Fox, P.F. Biochemistry of cheese ripening: Proteolysis. In Cheese Chemistry, Physics and Microbiology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 445–482. [Google Scholar] [CrossRef]
- Fallico, V.; McSweeney, P.L.H.; Horne, J.; Pediliggieri, C.; Hannon, J.A.; Carpino, S.; Licitra, G. Evaluation of bitterness in Ragusano cheese. J. Dairy Sci. 2005, 88, 1288–1300. [Google Scholar] [CrossRef] [PubMed]
- Ardö, Y. Flavour formation by amino acid catabolism. Biotechnol. Adv. 2006, 24, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.A.O.; Lucey, J.A.; McSweeney, P.L.H. Chymosin-mediated proteolysis, calcium solubilization, and texture development during the ripening of Cheddar cheese. J. Dairy Sci. 2005, 88, 3101–3114. [Google Scholar] [CrossRef] [PubMed]

 ), galactose (
), galactose ( ), citric acid (
), citric acid ( ), orotic acid (
), orotic acid ( ), lactic acid (
), lactic acid ( ), acetic acid (
), acetic acid ( ), and hippuric acid (
), and hippuric acid ( ). Different letters for the same compound indicate statistically significant differences (p > 0.05).
), galactose (), citric acid (), orotic acid (), lactic acid (), acetic acid (), and hippuric acid (). Different letters for the same compound indicate statistically significant differences (p > 0.05).
). Different letters for the same compound indicate statistically significant differences (p > 0.05).
), galactose (), citric acid (), orotic acid (), lactic acid (), acetic acid (), and hippuric acid (). Different letters for the same compound indicate statistically significant differences (p > 0.05).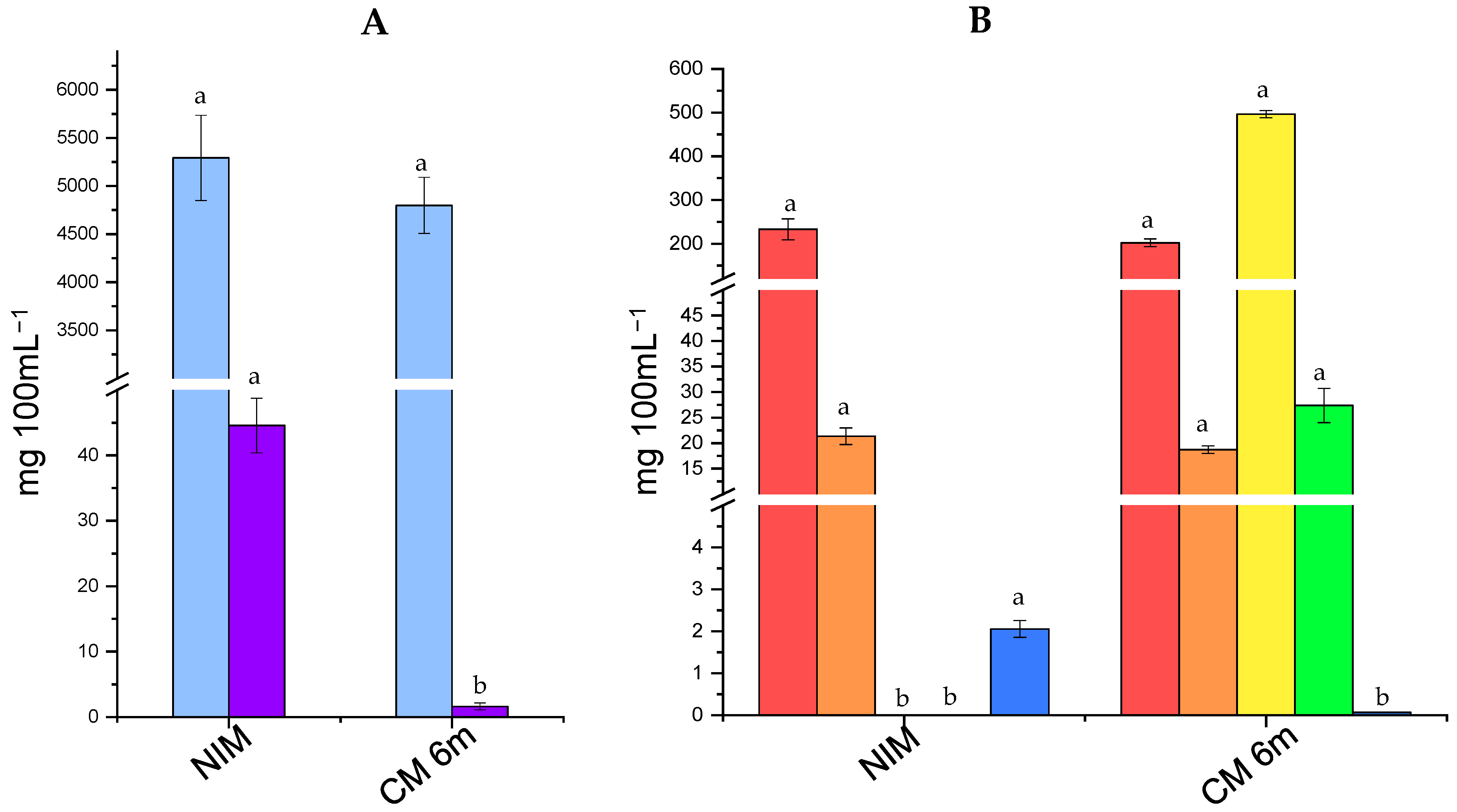
 ), galactose (
), galactose ( ), citric acid (
), citric acid ( ), orotic acid (
), orotic acid ( ), lactic acid (
), lactic acid ( ), acetic acid (
), acetic acid ( ), and hippuric acid (
), and hippuric acid ( ). Abbreviations: NRA,
cheese with no ripening agent; RA -20, cheese manufactured with L73
freeze-dried cells stored at −20 °C for 6 months as a ripening agent; RA 5,
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
a ripening agent. Bars for the same compound with the same letter did not
differ significantly (p > 0.05).
), galactose (), citric acid (), orotic acid (), lactic acid (), acetic acid (), and hippuric acid (). Abbreviations: NRA,
cheese with no ripening agent; RA -20, cheese manufactured with L73
freeze-dried cells stored at −20 °C for 6 months as a ripening agent; RA 5,
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
a ripening agent. Bars for the same compound with the same letter did not
differ significantly (p > 0.05).
). Abbreviations: NRA,
cheese with no ripening agent; RA -20, cheese manufactured with L73
freeze-dried cells stored at −20 °C for 6 months as a ripening agent; RA 5,
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
a ripening agent. Bars for the same compound with the same letter did not
differ significantly (p > 0.05).
), galactose (), citric acid (), orotic acid (), lactic acid (), acetic acid (), and hippuric acid (). Abbreviations: NRA,
cheese with no ripening agent; RA -20, cheese manufactured with L73
freeze-dried cells stored at −20 °C for 6 months as a ripening agent; RA 5,
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
a ripening agent. Bars for the same compound with the same letter did not
differ significantly (p > 0.05).
 ), cheese with no ripening agent; RA -20 (
), cheese with no ripening agent; RA -20 ( ), cheese
manufactured with L73 freeze-dried cells stored at −20 °C for 6 months as
ripening agent; RA 5 (
), cheese
manufactured with L73 freeze-dried cells stored at −20 °C for 6 months as
ripening agent; RA 5 ( ), cheese manufactured with L73 freeze-dried cells stored
at 5 °C for 6 months as ripening agent.
), cheese with no ripening agent; RA -20 (), cheese
manufactured with L73 freeze-dried cells stored at −20 °C for 6 months as
ripening agent; RA 5 (), cheese manufactured with L73 freeze-dried cells stored
at 5 °C for 6 months as ripening agent.
), cheese manufactured with L73 freeze-dried cells stored
at 5 °C for 6 months as ripening agent.
), cheese with no ripening agent; RA -20 (), cheese
manufactured with L73 freeze-dried cells stored at −20 °C for 6 months as
ripening agent; RA 5 (), cheese manufactured with L73 freeze-dried cells stored
at 5 °C for 6 months as ripening agent.

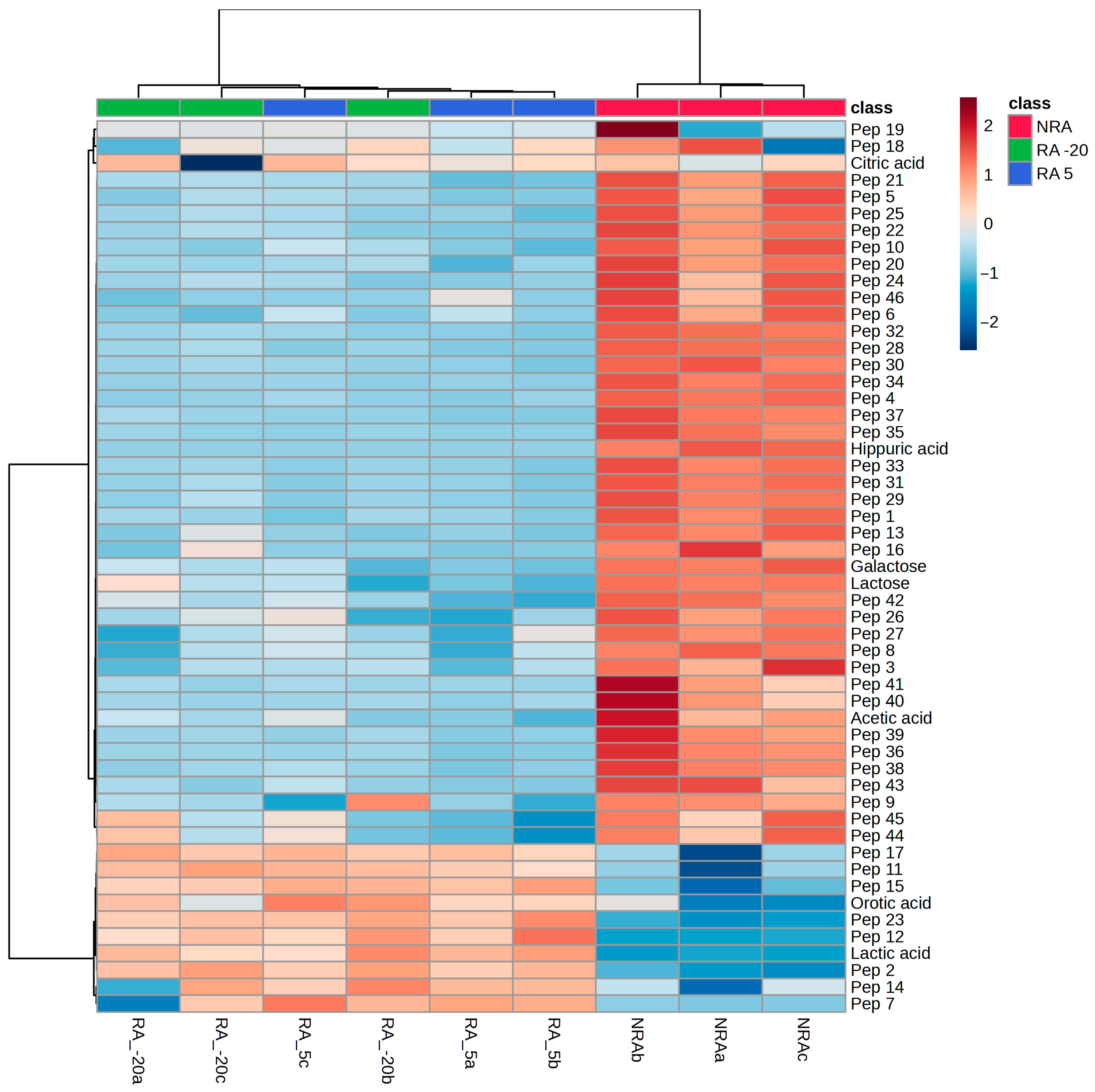
 ), cheese with no
ripening agent; RA -20 (
), cheese with no
ripening agent; RA -20 ( ), cheese manufactured with L73 freeze-dried cells
stored at −20 °C for 6 months as ripening agent; RA 5 (
), cheese manufactured with L73 freeze-dried cells
stored at −20 °C for 6 months as ripening agent; RA 5 ( ),
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
ripening agent.
), cheese with no
ripening agent; RA -20 (), cheese manufactured with L73 freeze-dried cells
stored at −20 °C for 6 months as ripening agent; RA 5 (),
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
ripening agent.
),
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
ripening agent.
), cheese with no
ripening agent; RA -20 (), cheese manufactured with L73 freeze-dried cells
stored at −20 °C for 6 months as ripening agent; RA 5 (),
cheese manufactured with L73 freeze-dried cells stored at 5 °C for 6 months as
ripening agent.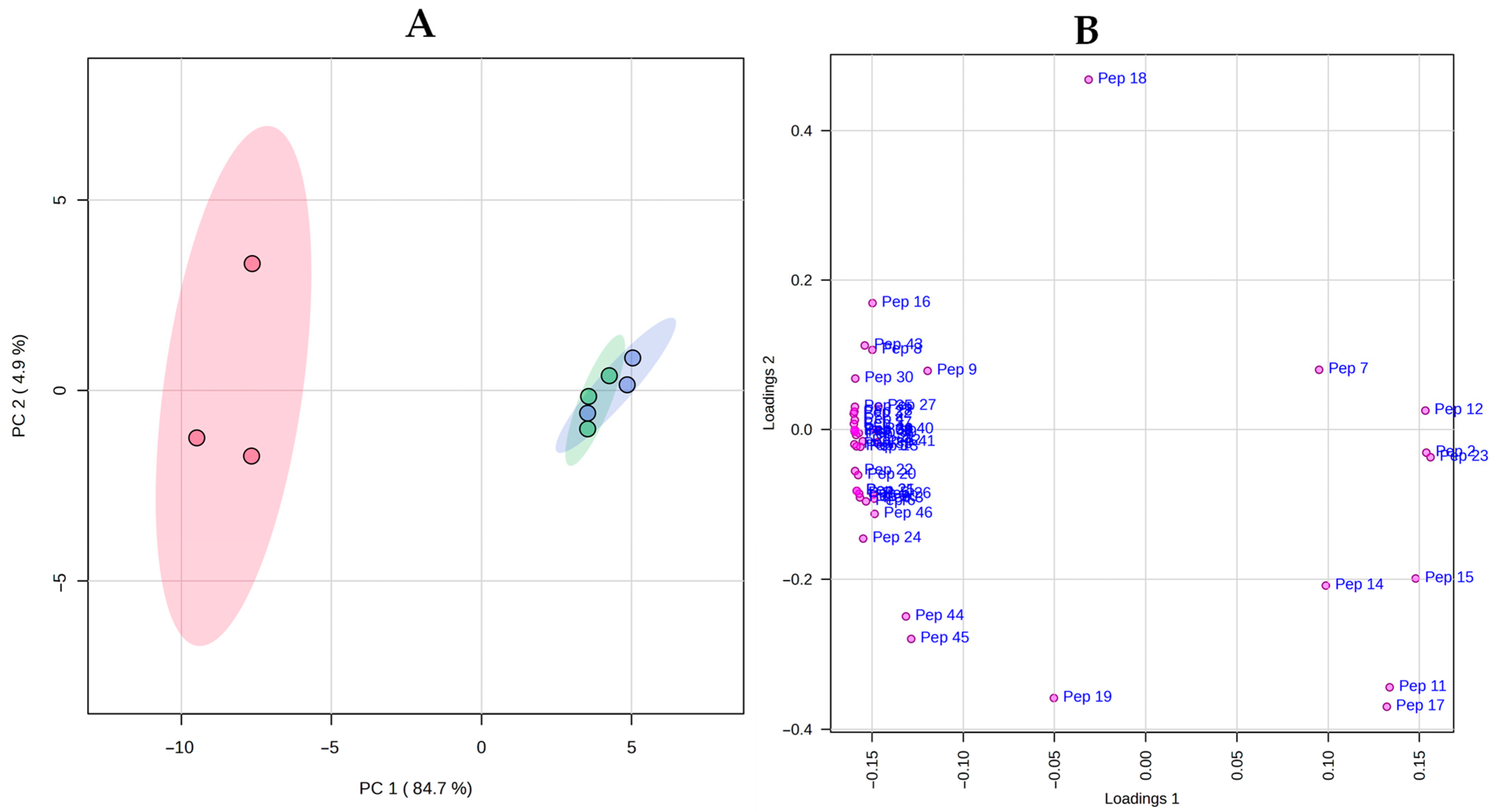

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | NRA | RA -20 | RA 5 |
|---|---|---|---|
| Moisture (%, w/w) | 33.6 ± 0.7 a | 34.7 ± 0.7 a | 34.7 ± 0.9 a |
| Fat (%, w/w) | 34.5 ± 0.0 a | 34.6 ± 0.6 a | 34.5 ± 0.4 a |
| Protein (%, w/w) | 26.4 ± 0.6 a | 26.6 ± 0.8 a | 26.2 ± 0.9 a |
| pH | 5.45 ± 0.01 a | 4.87 ± 0.03 b | 4.87 ± 0.03 b |
| Parameter | NRA | RA -20 | RA 5 |
|---|---|---|---|
| Total LAB (log cfu g−1) | 9.0 ± 0.1 a | 8.7 ± 0.1 a | 8.6 ± 0.1 a |
| Lactobacilli (log cfu g−1) | <4 | 7.6 ± 0.1 | 7.1 ± 0.5 |
| Enterococci (log cfu g−1) | <3 | <3 | <3 |
| Molds and yeasts (log cfu g−1) | <2 | <2 | <2 |
| Coliforms (log cfu g−1) | <1 | <1 | <1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batistela, M.E.; Bergamini, C.V.; Ale, E.C.; Peralta, G.H. Unlocking the Potential of Lacticaseibacillus rhamnosus 73 as a Ripening Agent in Semi-Hard Cheese After Freeze-Drying and a Six-Month Storage Period. Fermentation 2025, 11, 409. https://doi.org/10.3390/fermentation11070409
Batistela ME, Bergamini CV, Ale EC, Peralta GH. Unlocking the Potential of Lacticaseibacillus rhamnosus 73 as a Ripening Agent in Semi-Hard Cheese After Freeze-Drying and a Six-Month Storage Period. Fermentation. 2025; 11(7):409. https://doi.org/10.3390/fermentation11070409
Chicago/Turabian StyleBatistela, Mara E., Carina V. Bergamini, Elisa C. Ale, and Guillermo H. Peralta. 2025. "Unlocking the Potential of Lacticaseibacillus rhamnosus 73 as a Ripening Agent in Semi-Hard Cheese After Freeze-Drying and a Six-Month Storage Period" Fermentation 11, no. 7: 409. https://doi.org/10.3390/fermentation11070409
APA StyleBatistela, M. E., Bergamini, C. V., Ale, E. C., & Peralta, G. H. (2025). Unlocking the Potential of Lacticaseibacillus rhamnosus 73 as a Ripening Agent in Semi-Hard Cheese After Freeze-Drying and a Six-Month Storage Period. Fermentation, 11(7), 409. https://doi.org/10.3390/fermentation11070409