
Bioprospecting for a Wild Strain of Sporisorium scitamineum for the Valorization of Sugarcane Molasses into Mannosylerythritol Lipids and Cellobiose Lipids

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Chemicals
2.3. Isolation and Genetic Analysis of Wild-Type S. scitamineum
2.3.1. Isolation of S. scitamineum
2.3.2. DNA Extraction, PCR Amplification, and Sequencing
2.3.3. Phylogenetic Analysis
2.4. Culture Conditions
2.4.1. Stock Cultures
2.4.2. Seed Cultures
2.4.3. Co-Production of MELs and CBLs
2.5. Isolation and Purification of MELs and CBLs
2.6. Analytical Methods
2.6.1. Microscope Imaging
2.6.2. UV-Spectrometry
2.6.3. High-Performance Liquid Chromatography
2.7. Calculations
2.8. Statistical Analysis
3. Results
3.1. Isolation and Identification of Wild-Type S. scitamineum
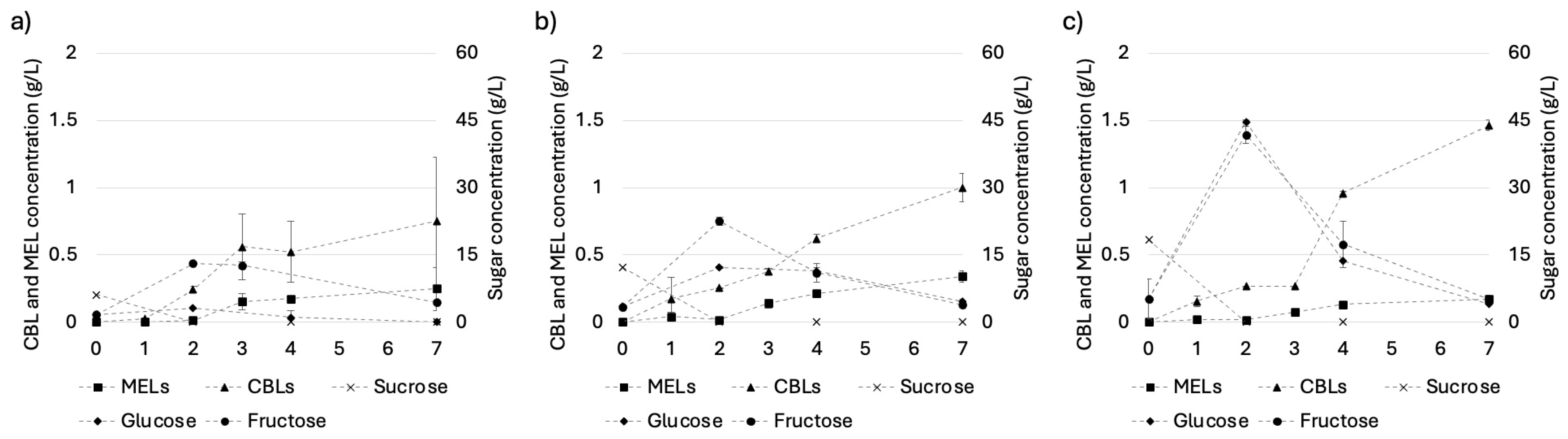
3.2. The Co-Production of MELs and CBLs from Pure Hydrophilic Carbon Sources by Wild-Type S. scitamineum
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Formann, S.; Hahn, A.; Janke, L.; Stinner, W.; Sträuber, H.; Logroño, W.; Nikolausz, M. Beyond Sugar and Ethanol Production: Value Generation Opportunities Through Sugarcane Residues. Front. Energy Res. 2020, 8, 267. [Google Scholar] [CrossRef]
- Dinesh Babu, K.S.; Janakiraman, V.; Palaniswamy, H.; Kasirajan, L.; Gomathi, R.; Ramkumar, T.R. A short review on sugarcane: Its domestication, molecular manipulations and future perspectives. Genet. Resour. Crop Evol. 2022, 69, 2623. [Google Scholar] [CrossRef] [PubMed]
- Hawaz, E.; Tafesse, M.; Tesfaye, A.; Kiros, S.; Beyene, D.; Kebede, G.; Boekhout, T.; Groenwald, M.; Theelen, B.; Degefe, A.; et al. Optimization of bioethanol production from sugarcane molasses by the response surface methodology using Meyerozyma caribbica isolate MJTm3. Ann. Microbiol. 2023, 73, 2. [Google Scholar] [CrossRef]
- Luo, R.; Zhang, Y.; Wang, F.; Liu, K.; Huang, G.; Zheng, N.; Wang, J. Effects of Sugar Cane Molasses Addition on the Fermentation Quality, Microbial Community, and Tastes of Alfalfa Silage. Animals 2021, 11, 355. [Google Scholar] [CrossRef]
- Bento, H.B.S.; Carvalho, A.K.F.; Reis, C.E.R.; De Castro, H.F. Single cell oil production and modification for fuel and food applications: Assessing the potential of sugarcane molasses as culture medium for filamentous fungus. Ind. Crops Prod. 2020, 145, 112141. [Google Scholar] [CrossRef]
- Morita, T.; Ishibashi, Y.; Hirose, N.; Wada, K.; Takahashi, M.; Fukuoka, T.; Imura, T.; Sakai, H.; Abe, M.; Kitamoto, D. Production and Characterization of a Glycolipid Biosurfactant, Mannosylerythritol Lipid B, from Sugarcane Juice by Ustilago scitaminea NBRC 32730. Biosci. Biotechnol. Biochem. 2011, 75, 1371–1376. [Google Scholar] [CrossRef]
- Teixeira-Silva, N.S.; Schaker, P.D.C.; Rody, H.V.S.; Maia, T.; Garner, C.M.; Gassmann, W.; Monteiro-Vitorello, C.B. Leaping into the Unknown World of Sporisorium scitamineum Candidate Effectors. J. Fungi 2020, 6, 339. [Google Scholar] [CrossRef]
- Bhuiyan, S.A.; Magarey, R.C.; McNeil, M.D.; Aitken, K.S. Sugarcane Smut, Caused by Sporisorium scitamineum, a Major Disease of Sugarcane: A Contemporary Review. Phytopathology 2021, 111, 1905–1917. [Google Scholar] [CrossRef]
- Oraby, A.; Werner, N.; Sungur, Z.; Zibek, S. Factors Affecting the Synthesis of Cellobiose Lipids by Sporisorium scitamineum. Front. Bioeng. Biotechnol. 2020, 8, 555647. [Google Scholar] [CrossRef]
- Morita, T.; Ishibashi, Y.; Fukuoka, T.; Imura, T.; Sakai, H.; Abe, M.; Kitamoto, D. Production of glycolipid biosurfactants, mannosylerythritol lipids, by a smut fungus, Ustilago scitaminea NBRC 32730. Biosci. Biotechnol. Biochem. 2009, 73, 788–792. [Google Scholar] [CrossRef]
- Kitamoto, D.; Fukuoka, T.; Saika, A.; Morita, T. Glycolipid Biosurfactants, Mannosylerythritol Lipids: Distinctive Interfacial Properties and Applications in Cosmetic and Personal Care Products. J. Oleo Sci. 2021, 71, ess21358. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.L.S.; Feuser, P.E.; Carciofi, B.A.M.; de Andrade, C.J.; de Oliveira, D. Mannosylerythritol lipids: Antimicrobial and biomedical properties. Appl. Microbiol. Biotechnol. 2020, 104, 2297–2318. [Google Scholar] [CrossRef]
- Kulakovskaya, E.; Baskunov, B.; Zvonarev, A. The Antibiotic and Membrane-damaging Activities of Cellobiose Lipids and Sophorose Lipids. J. Oleo Sci. 2014, 63, 701–707. [Google Scholar] [CrossRef]
- Imura, T.; Kawamura, D.; Ishibashi, Y.; Morita, T.; Sato, S.; Fukuoka, T.; Kikkawa, Y.; Kitamoto, D. Low molecular weight gelators based on biosurfactants, cellobiose lipids by cryptococcus humicola. J. Oleo Sci. 2012, 61, 659–664. [Google Scholar] [CrossRef]
- Imura, T.; Yamamoto, S.; Yamashita, C.; Taira, T.; Minamikawa, H.; Morita, T.; Kitamoto, D. Aqueous Gel Formation from Sodium Salts of Cellobiose Lipids. J. Oleo Sci. 2014, 63, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Moldes, A.B.; Rodríguez-López, L.; Rincón-Fontán, M.; López-Prieto, A.; Vecino, X.; Cruz, J.M. Synthetic and Bio-Derived Surfactants Versus Microbial Biosurfactants in the Cosmetic Industry: An Overview. Int. J. Mol. Sci. 2021, 22, 2371. [Google Scholar] [CrossRef] [PubMed]
- Jimoh, A.A.; Lin, J. Biosurfactant: A new frontier for greener technology and environmental sustainability. Ecotoxicol. Environ. Saf. 2019, 184, 109607. [Google Scholar] [CrossRef]
- Shaban, S.M.; Kang, J.; Kim, D.H. Surfactants: Recent advances and their applications. Compos. Commun. 2020, 22, 100537. [Google Scholar] [CrossRef]
- Jahan, R.; Bodratti, A.M.; Tsianou, M.; Alexandridis, P. Biosurfactants, natural alternatives to synthetic surfactants: Physicochemical properties and applications. Adv. Colloid Interface Sci. 2020, 275, 102061. [Google Scholar] [CrossRef]
- Yu, C.; Qi, J.; Han, H.; Wang, P.; Liu, C. Progress in pathogenesis research of Ustilago maydis, and the metabolites involved along with their biosynthesis. Mol. Plant Pathol. 2023, 24, 495. [Google Scholar] [CrossRef]
- Seliger, H. PCR protocols—A guide to methods and applications. Trends Biotechnol. 1990, 8, 335. [Google Scholar] [CrossRef]
- McTaggart, A.R.; Shivas, R.G.; Geering, A.D.W.; Callaghan, B.; Vánky, K.; Scharaschkin, T. Soral synapomorphies are significant for the systematics of the Ustilago-Sporisorium-Macalpinomyces complex (Ustilaginaceae). Persoonia 2012, 29, 63–77. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Konishi, M.; Fukuoka, T.; Imura, T.; Kitamoto, D. Production of glycolipid biosurfactants, mannosylerythritol lipids, by Pseudozyma siamensis CBS 9960 and their interfacial properties. J. Biosci. Bioeng. 2008, 105, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Ishibashi, Y.; Fukuoka, T.; Imura, T.; Sakai, H.; Abe, M.; Kitamoto, D. Production of Glycolipid Biosurfactants, Mannosylerythritol Lipids, Using Sucrose by Fungal and Yeast Strains, and Their Interfacial Properties. Biosci. Biotechnol. Biochem. 2009, 73, 2352–2355. [Google Scholar] [CrossRef]
- Arnstein, H.R.V.; Dawson, R.M.C.; Elliott, D.C.; Elliott, W.H.; Jones, K.M. Data for Biochemical Research, 3rd ed.; Clarendon Press: Oxford, UK, 1988; Volume 234, p. 506. [Google Scholar]
- Valkenburg, A.D.; Teke, G.M.; Pott, R.W.M.; van Rensburg, E. The fed-batch production of mannosylerythritol lipids by Ustilago maydis DSM 4500 from hydrophilic carbon sources. Bioprocess. Biosyst. Eng. 2024, 47, 2043–2054. [Google Scholar] [CrossRef]
- Oraby, A.; Rupp, S.; Zibek, S. Techno-Economic Analysis as a Driver for Optimisation of Cellobiose Lipid Fermentation and Purification. Front. Bioeng. Biotechnol. 2022, 10, 913351. [Google Scholar] [CrossRef]
- Morita, T.; Ishibashi, Y.; Fukuoka, T.; Imura, T.; Sakai, H.; Abe, M.; Kitamoto, D. Production of Glycolipid Biosurfactants, Cellobiose Lipids, by Cryptococcus humicola JCM 1461 and Their Interfacial Properties. Biosci. Biotechnol. Biochem. 2011, 75, 1597–1599. [Google Scholar] [CrossRef]
- Zhou, B.; Hao, Q.; Liang, Y.; Kong, E. Protein palmitoylation in cancer: Molecular functions and therapeutic potential. Mol. Oncol 2023, 17, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Martín Ortega, A.M.; Segura Campos, M.R. Macronutrients and micronutrients in cancer prevention and treatment. Oncol. Funct. Nutr. Phytochem. Med. Plants 2021, 99–124. [Google Scholar] [CrossRef]
- Wada, K.; Koike, H.; Fujii, T.; Morita, T. Targeted transcriptomic study of the implication of central metabolic pathways in mannosylerythritol lipids biosynthesis in Pseudozyma antarctica T-34. PLoS ONE 2020, 15, e0227295. [Google Scholar] [CrossRef]
- Beck, A.; Werner, N.; Zibek, S. Mannosylerythritol Lipids: Biosynthesis, Genetics, and Production Strategies. In Biobased Surfactants; Elsevier: Amsterdam, The Netherlands, 2019; pp. 121–167. [Google Scholar]
- Hewald, S.; Josephs, K.; Bölker, M. Genetic analysis of biosurfactant production in Ustilago maydis. Appl. Environ. Microbiol 2005, 71, 3033–3040. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Snetselaar, K.; Mims, C. Infection of maize stigmas by Ustilago maydis: Light and electron microscopy. Phytopathology 1993, 83, 843. [Google Scholar] [CrossRef]
- Sipaúba-Tavares, L.H.; Tedesque, M.G.; Colla, L.C.; Millan, R.N.; Scardoeli-Truzzi, B. Effect of untreated and pretreated sugarcane molasses on growth performance of Haematococcus pluvialis microalgae in inorganic fertilizer and macrophyte extract culture media. Braz. J. Biol. 2022, 82, e263282. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source | CBLs | MELs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Titre (g/L) | Yield per Biomass Formed (g/g) | Yield per Carbon Consumed (g/g) | Yield per Urea Consumed (g/g) | Production Rate (g/L·d) | Titre (g/L) | Yield per Biomass Formed (g/g) | Yield per Carbon Consumed (g/g) | Yield per Urea Consumed (g/g) | Production Rate (g/L·d) | |
| 50 g/L sucrose | 2.727 ± 0.680 | 0.701 ± 0.785 | 0.075 ± 0.019 | 4.544 ± 1.133 | 0.085 | 0.236 ± 0.046 | 0.061 ± 0.053 | 0.007 ± 0.001 | 0.394 ± 0.077 | 0.006 |
| 50 g/L glucose | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 |
| 50 g/L fructose | 0.711 ± 0.615 | 0.208 ± 1.091 | 0.064 ± 0.055 | 1.186 ± 1.025 | 0.077 | 0.022 ± 0.033 | 0.007 ± 0.058 | 0.002 ± 0.003 | 0.037 ± 0.054 | 0.004 |
| Carbon Source | CBLs | MELs | ||||||
|---|---|---|---|---|---|---|---|---|
| Titre (g/L) | Yield per Carbon Consumed (g/g) | Yield per Urea Consumed (g/g) | Production Rate (g/L·d) | Titre (g/L) | Yield per Carbon Consumed (g/g) | Yield per Urea Consumed (g/g) | Production Rate (g/L·d) | |
| 36 g/L sugarcane molasses | 0.750 ± 0.478 | 0.138 ± 0.088 | 1.250 ± 0.797 | 0.107 | 0.246 ± 0.160 | 0.045 ± 0.030 | 0.411 ± 0.267 | 0.035 |
| 72 g/L sugarcane molasses | 1.001 ± 0.106 | 0.088 ± 0.009 | 1.668 ± 0.176 | 0.143 | 0.339 ± 0.044 | 0.030 ± 0.004 | 0.565 ± 0.074 | 0.048 |
| 108 g/L sugarcane molasses | 1.463 ± 0.039 | 0.072 ± 0.002 | 2.439 ± 0.065 | 0.209 | 0.170 ± 0.032 | 0.008 ± 0.002 | 0.283 ± 0.054 | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valkenburg, A.D.; van der Merwe, B.; Teke, G.M.; van Rensburg, E.; Pott, R.W.M. Bioprospecting for a Wild Strain of Sporisorium scitamineum for the Valorization of Sugarcane Molasses into Mannosylerythritol Lipids and Cellobiose Lipids. Fermentation 2025, 11, 384. https://doi.org/10.3390/fermentation11070384
Valkenburg AD, van der Merwe B, Teke GM, van Rensburg E, Pott RWM. Bioprospecting for a Wild Strain of Sporisorium scitamineum for the Valorization of Sugarcane Molasses into Mannosylerythritol Lipids and Cellobiose Lipids. Fermentation. 2025; 11(7):384. https://doi.org/10.3390/fermentation11070384
Chicago/Turabian StyleValkenburg, André D., Breyten van der Merwe, George M. Teke, Eugéne van Rensburg, and Robert W. M. Pott. 2025. "Bioprospecting for a Wild Strain of Sporisorium scitamineum for the Valorization of Sugarcane Molasses into Mannosylerythritol Lipids and Cellobiose Lipids" Fermentation 11, no. 7: 384. https://doi.org/10.3390/fermentation11070384
APA StyleValkenburg, A. D., van der Merwe, B., Teke, G. M., van Rensburg, E., & Pott, R. W. M. (2025). Bioprospecting for a Wild Strain of Sporisorium scitamineum for the Valorization of Sugarcane Molasses into Mannosylerythritol Lipids and Cellobiose Lipids. Fermentation, 11(7), 384. https://doi.org/10.3390/fermentation11070384