


Enhanced Biogas Production from Glucose and Glycerol by Artificial Consortia of Anaerobic Sludge with Immobilized Yeast


Abstract
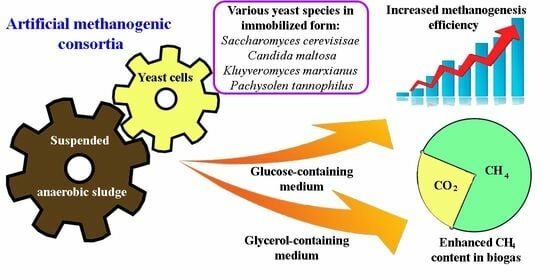
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Yeast Cells Immobilization
2.3. Biogas Production by Artificial Consortia
2.4. Static Test of Artificial Consortia in Methanogenic Treatment of Wastewater
2.5. Analytical Methods
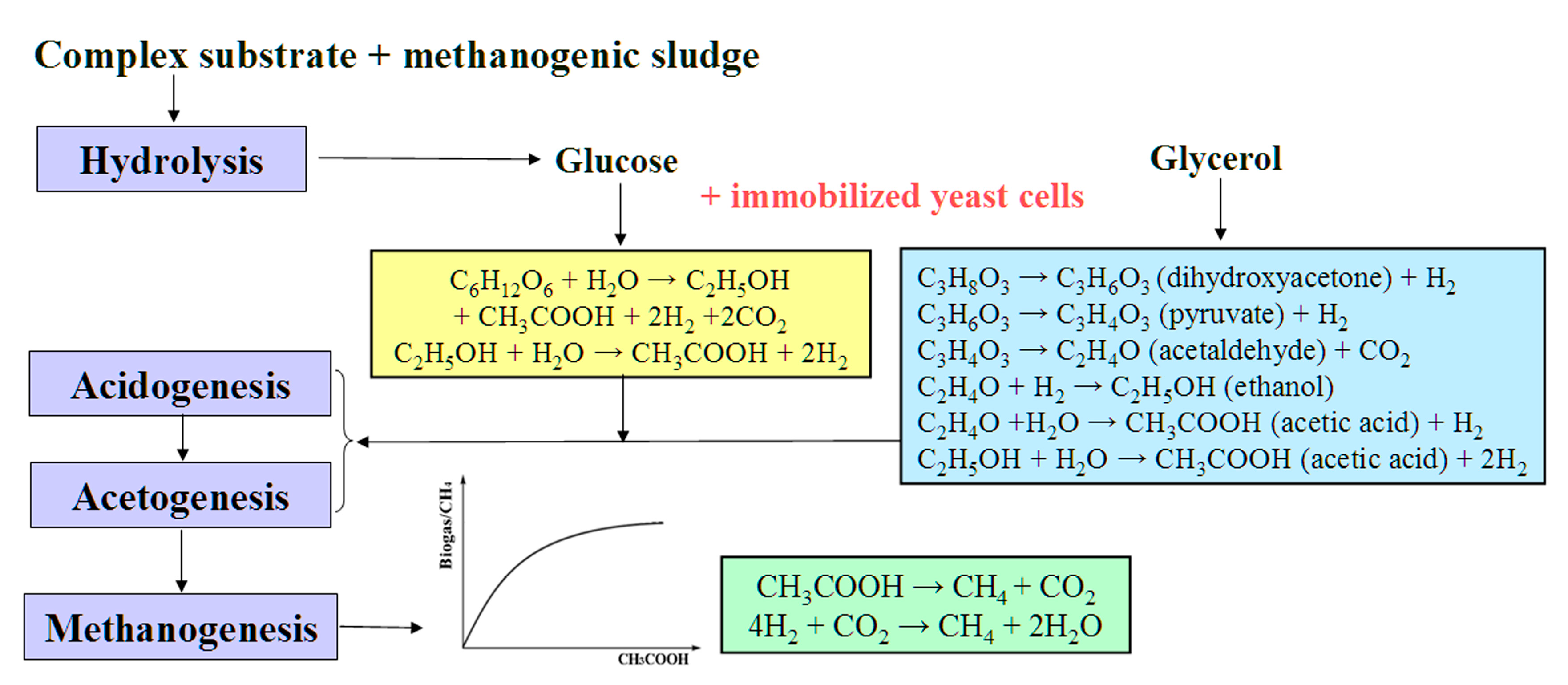
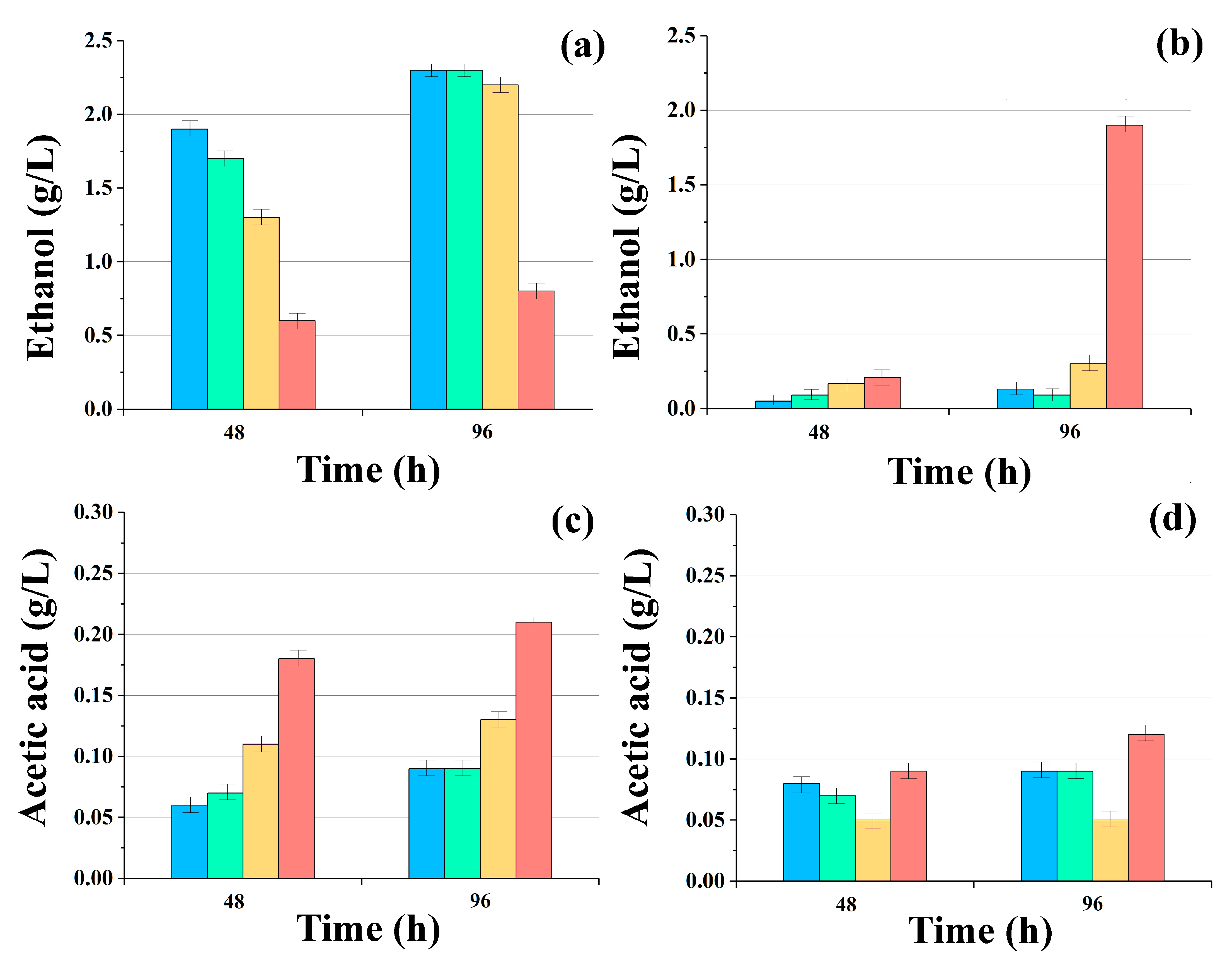
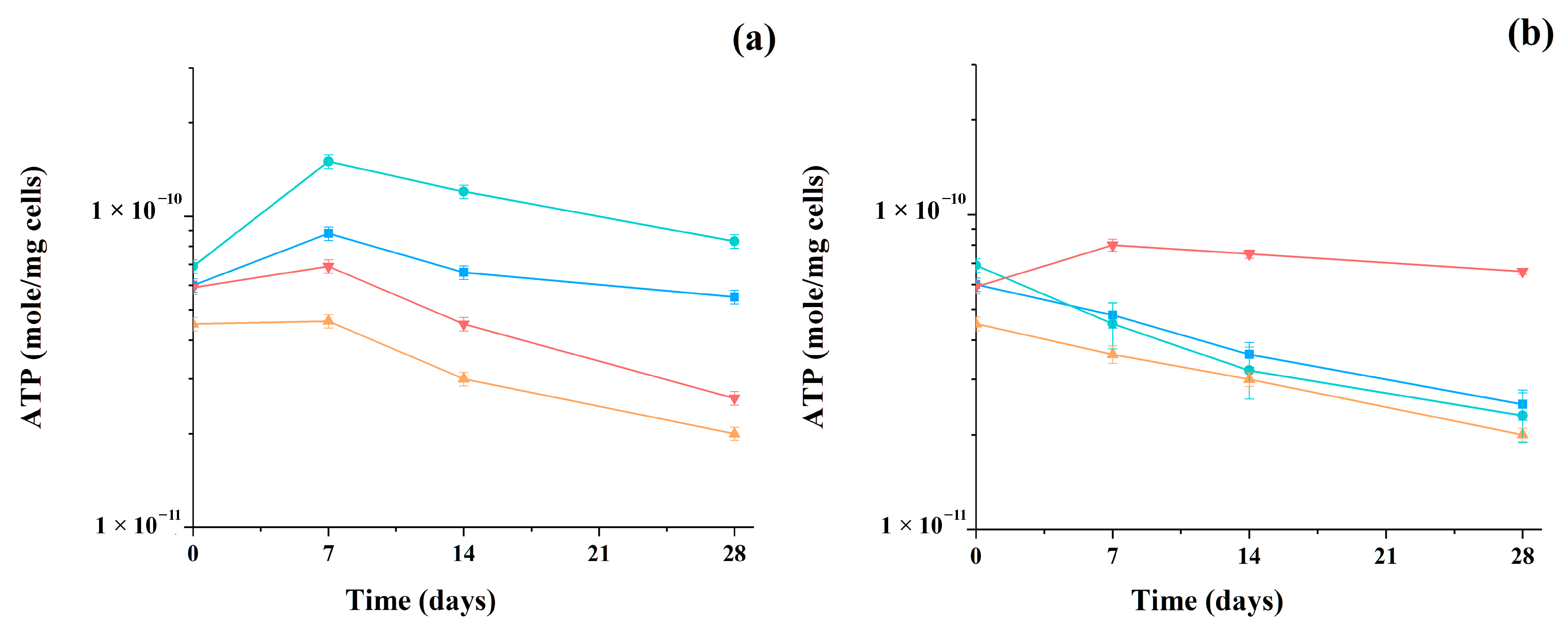
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AS | Anaerobic sludge |
| ATP | Adenosine triphosphate |
| COD | Chemical oxygen demand |
| DMSO | Dimethyl sulfoxide |
| SCFA | Short-chain fatty acids |
| VS | Volatile solids |
References
- Lawson, C.E.; Harcombe, W.R.; Hatzenpichler, R.; Lindemann, S.R.; Löffler, F.E.; O’Malley, M.A.; Martín, H.G.; Pfleger, B.F.; Raskin, L.; Venturelli, O.S.; et al. Common principles and best practices for engineering microbiomes. Nat. Rev. Microbiol. 2019, 17, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Lü, H.; Wei, J.-L.; Tang, G.-X.; Chen, Y.-S.; Huang, Y.-H.; Hu, R.; Mo, C.; Zhao, H.-M.; Xiang, L.; Li, Y.-W.; et al. Microbial consortium degrading of organic pollutants: Source, degradation efficiency, pathway, mechanism and application. J. Clean. Prod. 2024, 451, 141913. [Google Scholar] [CrossRef]
- Adamu, K.S.; Bichi, Y.H.; Nasiru, A.Y.; Babangida, A.M.; Umar, M.M.; Usman, G.; Muhammad, R. Synthetic microbial consortia in bioremediation and biodegradation. Int. J. Res. Sci. Innov. Appl. Sci. 2023, 8, 232–241. [Google Scholar] [CrossRef]
- Mohanty, S.S.; Kumar, A. Enhanced degradation of anthraquinone dyes by microbial monoculture and developed consortium through the production of specific enzymes. Sci. Rep. 2021, 11, 7678. [Google Scholar] [CrossRef]
- Elumalai, P.; Parthipan, P.; Huang, M.; Muthukumar, B.; Cheng, L.; Govarthanan, M.; Rajasekar, A. Enhanced biodegradation of hydrophobic organic pollutants by the bacterial consortium: Impact of enzymes and biosurfactants. Environ. Pollut. 2021, 289, 117956. [Google Scholar] [CrossRef] [PubMed]
- Tondro, H.; Musivand, S.; Zilouei, H.; Bazarganipour, M.; Zargoosh, K. Biological production of hydrogen and acetone- butanol-ethanol from sugarcane bagasse and rice straw using co-culture of Enterobacter aerogenes and Clostridium acetobutylicum. Biomass Bioenergy 2020, 142, 105818. [Google Scholar] [CrossRef]
- Cao, Z.; Yan, W.; Ding, M.; Yuan, Y. Construction of microbial consortia for microbial degradation of complex compounds. Front. Bioeng. Biotechnol. 2022, 10, 1051233. [Google Scholar] [CrossRef]
- Efremenko, E.; Stepanov, N.; Maslova, O.; Senko, O.; Aslanli, A.; Lyagin, I. “Unity and Struggle of Opposites” as a basis for the functioning of synthetic bacterial immobilized consortium that continuously degrades organophosphorus pesticides. Microorganisms 2022, 10, 1394. [Google Scholar] [CrossRef]
- Efremenko, E.; Senko, O.; Stepanov, N.; Aslanli, A.; Maslova, O.; Lyagin, I. Quorum sensing as a trigger that improves characteristics of microbial biocatalysts. Microorganism 2023, 11, 1395. [Google Scholar] [CrossRef]
- Pourcelot, E.; Vigna, A.; Marlin, T.; Galeote, V.; Nidelet, T. Design of a new model yeast consortium for ecological studies of enological fermentation. Peer Community J. 2025, 5, e5. [Google Scholar] [CrossRef]
- Darvishi, F.; Rafatiyan, S.; Abbaspour Motlagh Moghaddam, M.H.; Atkinson, E.; Ledesma-Amaro, R. Applications of synthetic yeast consortia for the production of native and non-native chemicals. Crit. Rev. Biotechnol. 2024, 44, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, F.; Bäumler, M.; Arulrajah, P.; García Lima, J.D.J.; Hauke, S.; Stock, A.; Weuster-Botz, D. Artificial microbial consortia for bioproduction processes. Eng. Life Sci. 2023, 23, e2100152. [Google Scholar] [CrossRef] [PubMed]
- Malik, K.; Salama, E.-S.; El-Dalatony, M.M.; Jalalah, M.; Harraz, F.A.; Al-Assiri, M.S.; Zheng, Y.; Sharma, P.; Li, X. Co-fermentation of immobilized yeasts boosted bioethanol production from pretreated cotton stalk lignocellulosic biomass: Long-term investigation. Ind. Crops Prod. 2021, 159, 113122. [Google Scholar] [CrossRef]
- Kong, Z.; Li, L.; Kurihara, R.; Kubota, K.; Li, Y.-Y. Anaerobic treatment of N, N-dimethylformamide-containing wastewater by co-culturing two sources of inoculum. Water Res. 2018, 139, 228–239. [Google Scholar] [CrossRef]
- Alengebawy, A.; Ran, Y.; Osman, A.I.; Jin, K.; Samer, M.; Ai, P. Anaerobic digestion of agricultural waste for biogas production and sustainable bioenergy recovery: A review. Environ. Chem. Lett. 2024, 22, 2641–2668. [Google Scholar] [CrossRef]
- Chew, K.R.; Leong, H.Y.; Khoo, K.S.; Vo, D.V.N.; Anjum, H.; Chang, C.K.; Show, P.L. Effects of anaerobic digestion of food waste on biogas production and environmental impacts: A review. Environ. Chem. Lett. 2021, 19, 2921–2939. [Google Scholar] [CrossRef]
- Gao, M.; Zhang, S.; Ma, X.; Guan, W.; Song, N.; Wang, Q.; Wu, C. Effect of yeast addition on the biogas production performance of a food waste anaerobic digestion system. R. Soc. Open Sci. 2020, 7, 200443. [Google Scholar] [CrossRef] [PubMed]
- Zupančič, G.D.; Škrjanec, I.; Logar, R.M. Anaerobic co-digestion of excess brewery yeast in a granular biomass reactor to enhance the production of biomethane. Bioresour. Technol. 2012, 124, 328–337. [Google Scholar] [CrossRef]
- Zupančič, G.D.; Panjičko, M.; Zelić, B. Biogas production from brewer’s yeast using an anaerobic sequencing batch reactor. FTB 2017, 55, 187–196. [Google Scholar] [CrossRef]
- Primaloka, A.D.; Ardhannari, L.; Matin, H.H.; Sumardiono, S. Study of biogas production from cassava industrial waste by anaerobic process. MATEC Web Conf. 2018, 156, 03052. [Google Scholar] [CrossRef]
- Tang, X.; Liao, C.; Zhou, S.; Chen, C.; Li, L.; Lu, G.; Xuang, X.; Zhang, M.; Chen, C.; Li, P. Potential of perennial sorghum for biogas production: Pretreatment with yeast-contained inoculants during anaerobic storage. Fuel 2024, 359, 130365. [Google Scholar] [CrossRef]
- Syaichurrozi, I.; Rusdi, R.; Hidayat, T.; Bustomi, A. Kinetics studies impact of initial pH and addition of yeast Saccharomyces cerevisiae on biogas production from tofu wastewater in Indonesia. Int. J. Eng. Trans. B Appl. 2016, 29, 1037–1046. [Google Scholar]
- Islas-Espinoza, M.; de las Heras, A.; Vázquez-Chagoyán, J.C.; Salem, A. Anaerobic cometabolism of fruit and vegetable wastes using mammalian fecal inoculums: Fast assessment of biomethane production. J. Clean. Prod. 2017, 141, 1411–1418. [Google Scholar] [CrossRef]
- Syaichurrozi, I.; Basyir, M.F.; Farraz, R.M.; Rusdi, R. A preliminary study: Effect of initial pH and Saccharomyces cerevisiae addition on biogas production from acid-pretreated Salvinia molesta and kinetics. Energy 2020, 207, 118226. [Google Scholar] [CrossRef]
- Fang, H.; Shi, Y.; Li, D.; Song, L.; Li, Y.Y.; Liu, R.; Yuan, D.; Niu, Q. Synergistic co-digestion of waste commercial yeast and chicken manure: Kinetic simulation, DOM variation and microbial community assessment. Renew. Energy 2020, 162, 2272–2284. [Google Scholar] [CrossRef]
- Moeller, L.; Bauer, A.; Zehnsdorf, A.; Lee, M.Y.; Müller, R.A. Anaerobic co-digestion of waste yeast biomass from citric acid production and waste frying fat. Eng. Life Sci. 2018, 18, 425–433. [Google Scholar] [CrossRef]
- Zhao, S.; Li, P.; Fang, H.; Song, L.; Li, D.; Liu, R.; Niu, Q. Enhancement methane fermentation of Enteromorpha prolifera waste by Saccharomyces cerevisiae, batch kinetic investigation, dissolved organic matter characterization, and synergistic mechanism. Environ. Sci. Pollut. Res. 2020, 27, 16254–16267. [Google Scholar] [CrossRef] [PubMed]
- Jojoa-Unigarro, G.D.; González-Martínez, S. OFMSW Fermentation with different inocula and its effects on methane production. Waste Biomass Valor. 2023, 14, 1461–1476. [Google Scholar] [CrossRef]
- Kazimierowicz, J.; Dębowski, M.; Zieliński, M.; Kasiński, S.; Cruz Sanchez, J. Biotechnological valorization of waste glycerol into gaseous biofuels—A Review. Energies 2024, 17, 338. [Google Scholar] [CrossRef]
- de Mello, B.S.; Pozzi, A.; Rodrigues, B.C.G.; Costa, M.A.M.; Sarti, A. Anaerobic digestion of crude glycerol from biodiesel production for biogas generation: Process optimization and pilot scale operation. Environ. Res. 2024, 244, 117938. [Google Scholar] [CrossRef]
- Vikromvarasiri, N.; Koyama, M.; Kurniawan, W.; Pisutpaisal, N.; Nakasaki, K. Enhancing methane recovery by intermittent substrate feeding and microbial community response in anaerobic digestion of glycerol. Renew. Energy 2023, 204, 106–113. [Google Scholar] [CrossRef]
- Bułkowska, K.; Mikucka, W.; Pokój, T. Enhancement of biogas production from cattle manure using glycerine phase as a co-substrate in anaerobic digestion. Fuel 2022, 317, 123456. [Google Scholar] [CrossRef]
- Gebreegziabher, B.W.; Dubale, A.A.; Adaramola, M.S.; Morken, J. Advancing anaerobic digestion of biodiesel byproducts: A comprehensive review. Bioenerg. Res. 2025, 18, 15. [Google Scholar] [CrossRef]
- Stepanov, N.; Efremenko, E. Immobilised cells of Pachysolen tannophilus yeast for ethanol production from crude glycerol. N. Biotechnol. 2017, 34, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Ugarova, N.; Koksharov, M.; Lomakina, G. Reagent for Adenosine-5-Triphosphate Determination. Patent RU 2420594, 20 May 2009. [Google Scholar]
- Stepanov, N.; Senko, O.; Perminova, I.; Efremenko, E. A new approach to assess the effect of various humic compounds on the metabolic activity of cells participating in methanogenesis. Sustainability 2019, 11, e3158. [Google Scholar] [CrossRef]
- Sisman-Aydin, G.; Simsek, K. Municipal wastewater effects on the performance of nutrient removal, and lipid, carbohydrate, and protein productivity of blue-green algae Chroococcus turgidus. Sustainability 2022, 14, 17021. [Google Scholar] [CrossRef]
- Nwokolo, N.; Mukumba, P.; Obileke, K.; Enebe, M. Waste to Energy: A focus on the impact of substrate type in biogas production. Processes 2020, 8, 1224. [Google Scholar] [CrossRef]
- Kazimierowicz, J.; Dębowski, M.; Zieliński, M.; Ignaciuk, A.; Mlonek, S.; Cruz Sanchez, J. The biosynthesis of liquid fuels and other value-added products based on waste glycerol—A comprehensive review and bibliometric analysis. Energies 2024, 17, 3035. [Google Scholar] [CrossRef]
- Efremenko, E.; Senko, O.; Stepanov, N.; Maslova, O.; Lomakina, G.Y.; Ugarova, N. Luminescent analysis of ATP: Modern objects and processes for sensing. Chemosensors 2022, 10, 493. [Google Scholar] [CrossRef]
- Strotmann, U.J.; Eismann, F.; Hauth, B.; Bias, W.R. An integrated test strategy for the assessment of anaerobic biodegradability of wastewaters. Chemosphere 1993, 26, 2241–2254. [Google Scholar] [CrossRef]
- Alves, I.R.; Mahler, C.F.; Oliveira, L.B.; Reis, M.M.; Bassin, J.P. Assessing the use of crude glycerol from biodiesel production as an alternative to boost methane generation by anaerobic co-digestion of sewage sludge. Biomass Bioenergy 2020, 143, 105831. [Google Scholar] [CrossRef]
- Lozinsky, V.I. Cryostructuring of polymeric systems. 55. Retrospective view on the more than 40 years of studies performed in the A.N. Nesmeyanov Institute of organoelement compounds with respect of the cryostructuring processes in polymeric systems. Gels 2020, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Senko, O.; Gladchenko, M.; Maslova, O.; Efremenko, E. Long-term storage and use of artificially immobilized anaerobic sludge as a powerful biocatalyst for conversion of various wastes including those containing xenobiotics to biogas. Catalysts 2019, 9, 326. [Google Scholar] [CrossRef]
- Kallistova, A.; Goel, G.; Nozhevnikova, A. Microbial diversity of methanogenic communities in the systems for anaerobic treatment of organic waste. Microbiology 2014, 83, 462–483. [Google Scholar] [CrossRef]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef]
- Maslova, O.; Senko, O.; Stepanov, N.; Gladchenko, M.; Gaydamaka, S.; Akopyan, A.; Polikarpova, P.; Lysenko, S.; Anisimov, A.; Efremenko, E. Formation and use of anaerobic consortia for the biotransformation of sulfur-containing extracts from pre-oxidized crude oil and oil fractions. Bioresour. Technol. 2021, 319, 124248. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast/Reference | * Conditions | Effects |
|---|---|---|
| Saccharomyces cerevisiae (Angel Yeast Co., Ltd., Hubei, China) [17] | Substrate—Food waste Biocatalysts—Sludge from a rural biogas station (80% by VS) and Yeast (2% VS); 37 °C, 60 rpm, 50 days | 33.2% increase in biogas production |
| Brewing yeast [18] | Substrate—Mixture of 50% brewery wastewater and 50% municipal wastewater (400–700 mg/L COD) Biocatalysts—Granular sludge (20% VS) and yeast (0.7%); 37 °C, pH 6.5, 120 rpm, 228 days | Increased biogas production |
| Brewing yeast [19] | Substrate—Brewery wastewater Biocatalysts—Granular sludge from brewery and yeast (2–22% v/v); 35 °C, pH 6.5, 7 days | 50% increase in biogas production |
| S. cerevisiae NKL [20] | Substrate—Cassava wastes 1% (w/v) with 0.4% urea (w/v) Biocatalysts—Cow rumen (8–20% w/v) and yeast (0.08–0.15% w/v); 26–33 °C, pH 7.0, 16 days | Increased biogas production |
| S. cerevisiae [21] | Substrate—Anaerobically pre-fermented sorghum biomass (500 g/L) Biocatalysts—Sludge from biogas plant and yeast (105 CFU/g wet substrate weight); 39 °C, 7 days | Enhanced production of total biogas (236–251 mL/g VS) and methane (96–105 mL/g VS) |
| S. cerevisiae Ragi [22] | Substrate—Tofu wastewater (COD—576 mg/L, total nitrogen—13.5 mg/L, pH 4.2) Biocatalysts—Rumen anaerobic consortium (10% v/v) and Yeast (1 g); pH 8.0, 8 days | 150% increase in biogas production |
| S. cerevisiae [23] | Substrate—Mixture of kitchen wastes (vegetables and fruits)—20 g/L Biocatalysts—Microbial consortia from fresh mixed mammal feces (20–500 g/L) and yeast with cellulolytic activity—400 mg/L (4 × 108 cells); 25, 39 or 55 °C, 96 h | S. cerevisiae improved cellulolytic activity in kitchen waste digestion and provided an increased biogas yield |
| S. cerevisiae [24] | Substrate—Acid-pretreated free-floating aquatic plant Salvinia molesta—10 g Biocatalysts—Cow rumen anaerobic consortium (25 mL) and yeast (1 g) in 130 mL H2O; pH 7.0, 30 °C, 30 days | Increase in biogas production (up to 113.7 mg/g VS) and CH4 content (up to 84%) |
| S. cerevisiae [25] | Substrate—Chicken manure—1.2 g Biocatalysts—Sludge from UASB (upflow AS blanket) reactor (5 g) and yeast (0.48–1.07 g/g VS); 35 °C, 120 rpm, 500 h | The highest biogas yield (470.95 mL/g VS) with 77.5% CH4 was observed at a yeast concentration of 0.69 g/g VS (4.3 times higher than the control without yeast cells) |
| Yarrowia lipolytica H181 from citric acid production [26] | Substrate—Waste frying fat Biocatalysts—AS from biogas plant treated maize silage (0.21–0.45 (VS/VS substrate) and yeast (5% w/w); pH 7.7–8.0, 70 days | Increase in biogas yield (1.45 m3/kg VS) with CH4 content—67 ± 4% |
| Wastewater | COD, g/L | Content of Main Components, % | pH | ||
|---|---|---|---|---|---|
| Proteins | Lipids | Carbohydrates | |||
| Milk plant | 1.4 ± 0.1 | 27.0 ± 2.5 | 41.6 ± 2.7 | 31.4 ± 3.5 | 6.8 |
| Bagasse hydrolysate | 42 ± 2.5 | 14.1 ± 1.2 | 11.7 ± 0.0 | 75.2 ± 3.5 | 5.0 |
| Meat processing | 4.0 ± 0.1 | 10.0 ± 0.3 | 35.0 ± 2.5 | 1.0 ± 0.1 | 6.0 |
| Consortia | CH4, mL/day | |
|---|---|---|
| Glucose | Glycerol | |
| AS (control without yeast) | 1.2 ± 0.06 | 1.1 ± 0.06 |
| AS + S. cerevisiae | 2.8 ± 0.14 | 1.9 ± 0.10 |
| AS + C. maltosa | 2.7 ± 0.13 | 2.2 ± 0.11 |
| AS + Kluyveromyces marxianus | 2.4 ± 0.12 | 2.0 ± 0.10 |
| AS + Pachysolen tannophilus | 1.6 ± 0.06 | 2.9 ± 0.15 |
| Sample of Wastewater | Initial COD, g/L | Type of Anaerobic Biocatalyst * | COD Removal, % | Efficiency of Biogas Production, % | CH4 Content, % |
|---|---|---|---|---|---|
| Wastewater from milk plant | 1.4 ± 0.1 | AS | 72 ± 2 | 47 ± 2 | 41 ± 2 |
| AS + K | 84 ± 3 | 63 ± 3 | 50 ± 3 | ||
| Wastewater with bagasse hydrolysate | 8.5 ± 0.3 | AS | 49 ± 2 | 41 ± 2 | 32 ± 1 |
| AS + S | 67 ± 3 | 65 ± 3 | 52 ± 3 | ||
| Wastewater with crude glycerol | 7.0 ± 0.2 | AS | 32 ± 1 | 22 ± 1 | 20 ± 1 |
| AS + P | 54 ± 2 | 47 ± 3 | 45 ± 2 | ||
| Wastewater from meat processing | 4.0 ± 0.1 | AS | 58 ± 2 | 42 ± 2 | 37 ± 1 |
| AS + C | 75 ± 3 | 64 ± 3 | 53 ± 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stepanov, N.; Senko, O.; Aslanli, A.; Maslova, O.; Efremenko, E. Enhanced Biogas Production from Glucose and Glycerol by Artificial Consortia of Anaerobic Sludge with Immobilized Yeast. Fermentation 2025, 11, 352. https://doi.org/10.3390/fermentation11060352
Stepanov N, Senko O, Aslanli A, Maslova O, Efremenko E. Enhanced Biogas Production from Glucose and Glycerol by Artificial Consortia of Anaerobic Sludge with Immobilized Yeast. Fermentation. 2025; 11(6):352. https://doi.org/10.3390/fermentation11060352
Chicago/Turabian StyleStepanov, Nikolay, Olga Senko, Aysel Aslanli, Olga Maslova, and Elena Efremenko. 2025. "Enhanced Biogas Production from Glucose and Glycerol by Artificial Consortia of Anaerobic Sludge with Immobilized Yeast" Fermentation 11, no. 6: 352. https://doi.org/10.3390/fermentation11060352
APA StyleStepanov, N., Senko, O., Aslanli, A., Maslova, O., & Efremenko, E. (2025). Enhanced Biogas Production from Glucose and Glycerol by Artificial Consortia of Anaerobic Sludge with Immobilized Yeast. Fermentation, 11(6), 352. https://doi.org/10.3390/fermentation11060352