
Enhancing Single-Cell Protein Yield Through Grass-Based Substrates: A Study of Lolium perenne and Kluyveromyces marxianus

Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Material
2.2. Pretreatment and Press Juice Preparation
2.3. Microbial Reduction Methods
2.4. Microorganism, Preculture
2.5. Preparation of Media and Agar Plate
2.6. Optimization Experiments in Shake Flasks
2.7. Analytical Methods
3. Results and Discussion
3.1. Parameter Optimization for Fermentation
3.1.1. Optimizing Microbial Reduction Methods for Fermentation
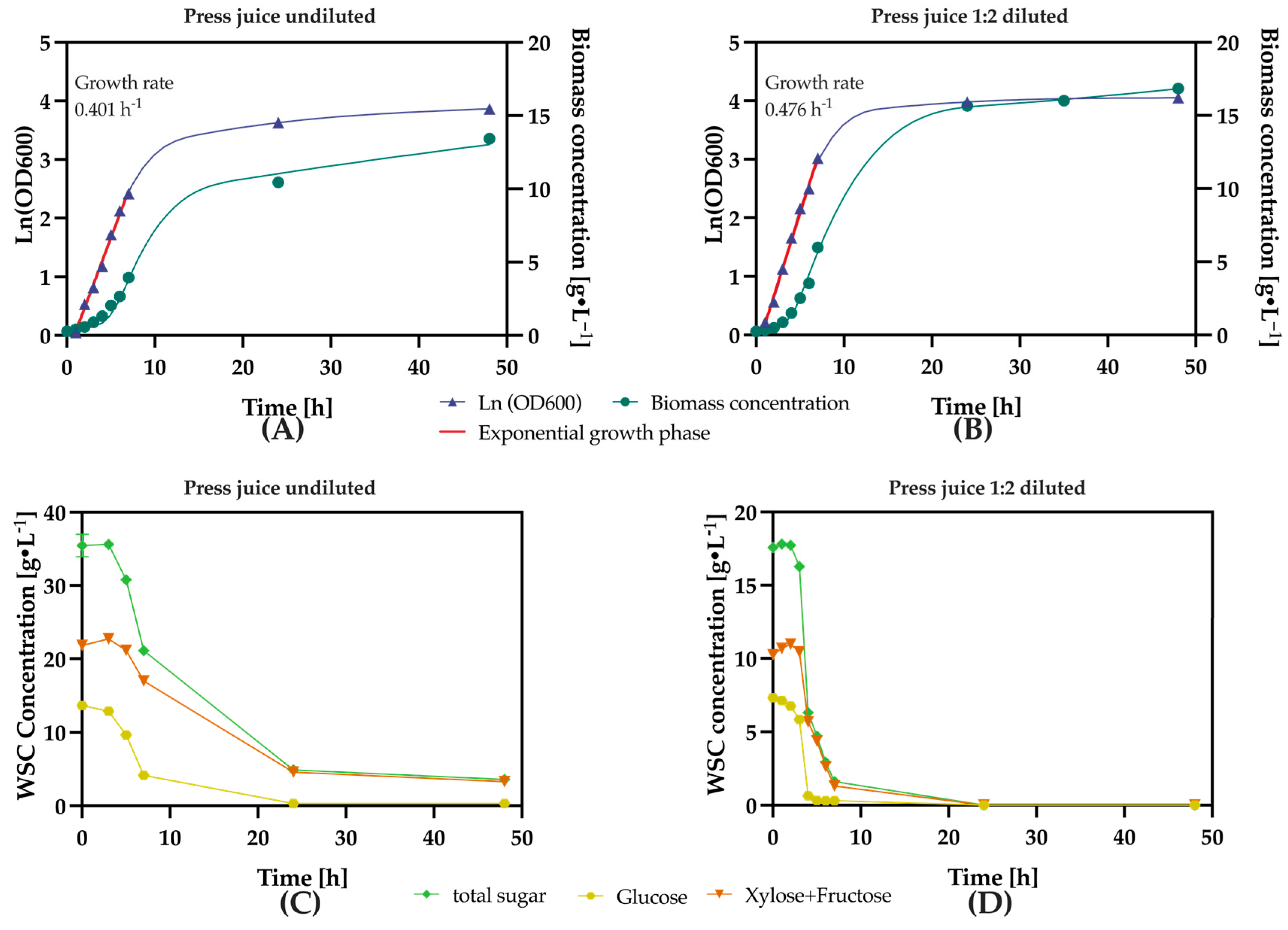
3.1.2. Optimizing Press Juice Dilution Ratios for Fermentation
3.1.3. Optimizing Temperature for Fermentation
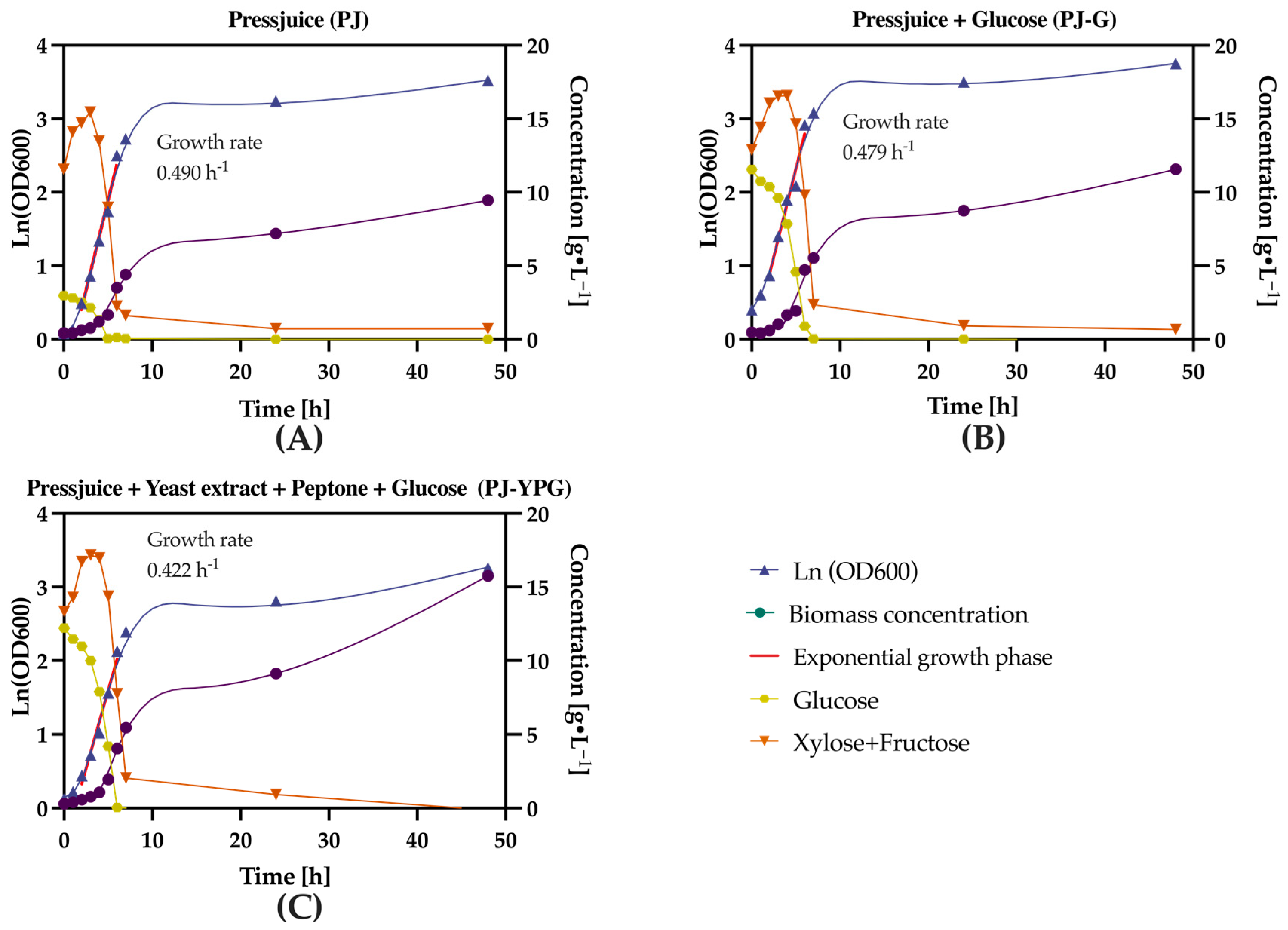
3.2. Effects of Press Juice Supplementation on K. marxianus Growth Rate, Biomass Yield, and Substrate Consumption
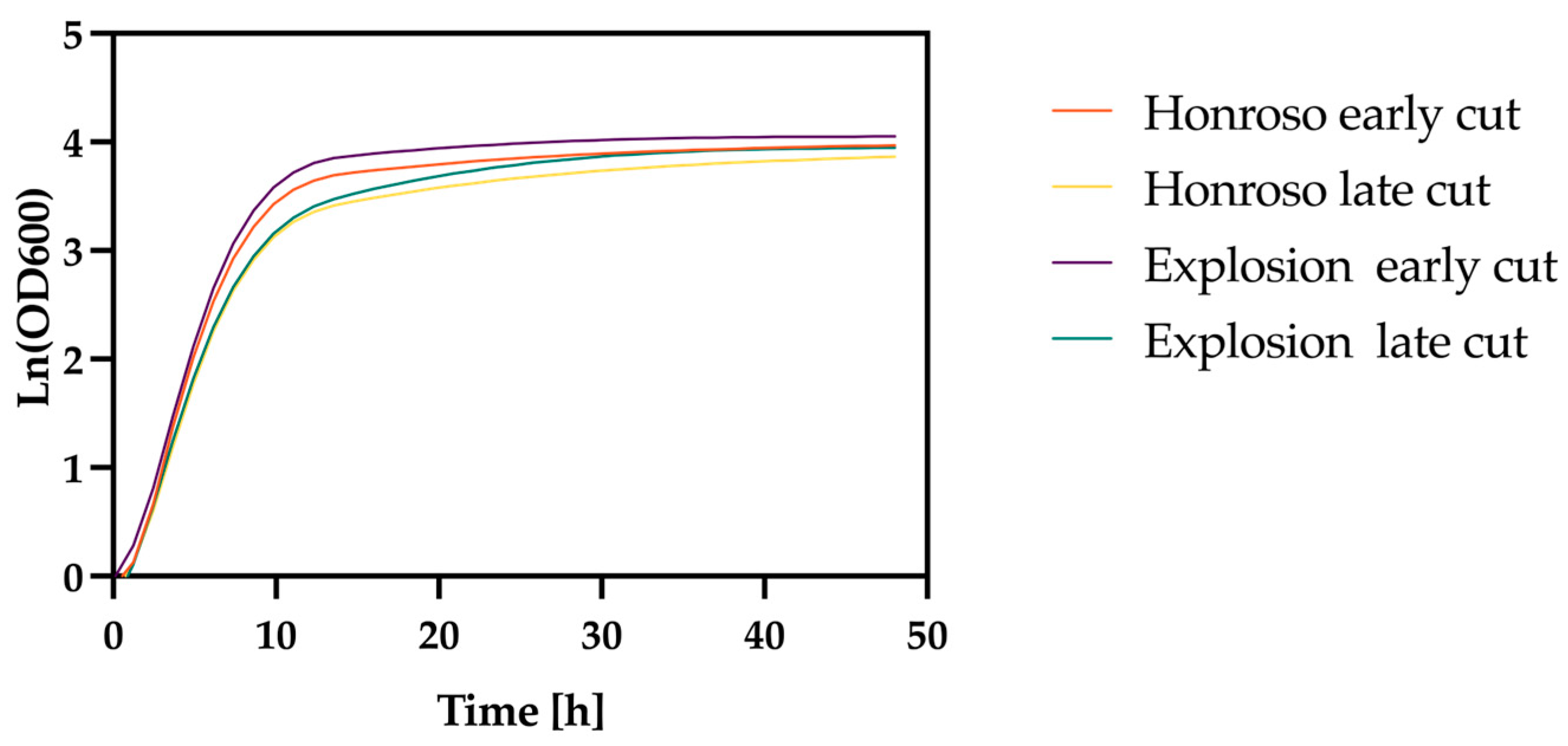
3.3. Effect of Lolium perenne Varieties and Harvest Time on Biomass Production
3.4. Evaluation of Field Yield and Harvest Timing Effects on Biomass and SCP Production in Different Lolium perenne Varieties
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwardson, W.; Lewis, C.W.; Slesser, M. Energy and Environmental Implications of Novel Protein Production Systems. Agric. Environ. 1981, 6, 1–21. [Google Scholar] [CrossRef]
- Najafpour, G.D. Single-Cell Protein. In Biochemical Engineering and Biotechnology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 417–434. ISBN 978-0-444-63357-6. [Google Scholar]
- Suman, G.; Nupur, M.; Anuradha, S.; Pradeep, B. Single Cell Protein Production: A Review. Int. J. Curr. Microbiol. Appl. Sci. (IJCMAS) 2015, 4, 251–262. [Google Scholar]
- Sekoai, P.T.; Roets-Dlamini, Y.; O’Brien, F.; Ramchuran, S.; Chunilall, V. Valorization of Food Waste into Single-Cell Protein: An Innovative Technological Strategy for Sustainable Protein Production. Microorganisms 2024, 12, 166. [Google Scholar] [CrossRef]
- Tropea, A.; Ferracane, A.; Albergamo, A.; Potortì, A.G.; Lo Turco, V.; Di Bella, G. Single Cell Protein Production through Multi Food-Waste Substrate Fermentation. Fermentation 2022, 8, 91. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The Yeast Kluyveromyces marxianus and Its Biotechnological Potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef]
- Leonel, L.V.; Arruda, P.V.; Chandel, A.K.; Felipe, M.G.A.; Sene, L. Kluyveromyces marxianus: A Potential Biocatalyst of Renewable Chemicals and Lignocellulosic Ethanol Production. Crit. Rev. Biotechnol. 2021, 41, 1131–1152. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Ji, L.; Xu, Y.; Xu, S.; Lin, Y.; Iqbal, H.M.N.; Cheng, H. Bioprospecting Kluyveromyces marxianus as a Robust Host for Industrial Biotechnology. Front. Bioeng. Biotechnol. 2022, 10, 851768. [Google Scholar] [CrossRef]
- Schwan, R.F.; Rose, A.H. Polygalacturonase Production by Kluyveromyces marxianus: Effect of Medium Composition. J. Appl. Bacteriol. 1994, 76, 62–67. [Google Scholar] [CrossRef]
- Struyf, N.; Vandewiele, H.; Herrera-Malaver, B.; Verspreet, J.; Verstrepen, K.J.; Courtin, C.M. Kluyveromyces marxianus Yeast Enables the Production of Low FODMAP Whole Wheat Breads. Food Microbiol. 2018, 76, 135–145. [Google Scholar] [CrossRef]
- Koukoumaki, D.I.; Papanikolaou, S.; Ioannou, Z.; Mourtzinos, I.; Sarris, D. Single-Cell Protein and Ethanol Production of a Newly Isolated Kluyveromyces marxianus Strain through Cheese Whey Valorization. Foods 2024, 13, 1892. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Bezawada, J.; Ajila, C.M.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Mixed Culture of Kluyveromyces marxianus and Candida krusei for Single-Cell Protein Production and Organic Load Removal from Whey. Bioresour. Technol. 2014, 164, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, D.D.V.; De Arruda, P.V.; Vicente, F.M.C.F.; Sene, L.; Da Silva, S.S.; Das Graças De Almeida Felipe, M. Evaluation of Fermentative Potential of Kluyveromyces marxianus ATCC 36907 in Cellulosic and Hemicellulosic Sugarcane Bagasse Hydrolysates on Xylitol and Ethanol Production. Ann. Microbiol. 2015, 65, 687–694. [Google Scholar] [CrossRef]
- Dong, L.; Wu, Y.; Li, M.; Zhang, C.; Cao, J.; Ledesma-Amaro, R.; Zhao, W.; Kang, D. Exploring the Fermentation Potential of Kluyveromyces marxianus NS127 for Single-Cell Protein Production. Fermentation 2025, 11, 70. [Google Scholar] [CrossRef]
- Bajpai, P. Single Cell Protein Production from Lignocellulosic Biomass; SpringerBriefs in Molecular Science; Springer: Singapore, 2017; ISBN 978-981-10-5872-1. [Google Scholar]
- Patelski, P.; Berłowska, J.; Balcerek, M.; Dziekońska-Kubczak, U.; Pielech-Przybylska, K.; Dygas, D.; Jędrasik, J. Conversion of Potato Industry Waste into Fodder Yeast Biomass. Processes 2020, 8, 453. [Google Scholar] [CrossRef]
- Malairuang, K.; Krajang, M.; Rotsattarat, R.; Chamsart, S. Intensive Multiple Sequential Batch Simultaneous Saccharification and Cultivation of Kluyveromyces marxianus SS106 Thermotolerant Yeast Strain for Single-Step Ethanol Fermentation from Raw Cassava Starch. Processes 2020, 8, 898. [Google Scholar] [CrossRef]
- Nunta, R.; Techapun, C.; Sommanee, S.; Mahakuntha, C.; Porninta, K.; Punyodom, W.; Phimolsiripol, Y.; Rachtanapun, P.; Wang, W.; Zhuang, X.; et al. Valorization of Rice Straw, Sugarcane Bagasse and Sweet Sorghum Bagasse for the Production of Bioethanol and Phenylacetylcarbinol. Sci. Rep. 2023, 13, 727. [Google Scholar] [CrossRef]
- Serra, A.-A.; Couée, I.; Heijnen, D.; Michon-Coudouel, S.; Sulmon, C.; Gouesbet, G. Genome-Wide Transcriptional Profiling and Metabolic Analysis Uncover Multiple Molecular Responses of the Grass Species Lolium perenne Under Low-Intensity Xenobiotic Stress. Front. Plant Sci. 2015, 6, 1124. [Google Scholar] [CrossRef]
- Bocian, A.; Kosmala, A.; Rapacz, M.; Jurczyk, B.; Marczak, Ł.; Zwierzykowski, Z. Differences in Leaf Proteome Response to Cold Acclimation between Lolium perenne Plants with Distinct Levels of Frost Tolerance. J. Plant Physiol. 2011, 168, 1271–1279. [Google Scholar] [CrossRef]
- Waldron, K.W. Advances in Biorefineries: Biomass and Waste Supply Chain Exploitation; Woodhead Publishing Series in Energy; Elsevier/Woodhead Publishing: Amsterdam, The Netherlands; Boston, MA, USA, 2014; ISBN 978-0-85709-521-3. [Google Scholar]
- Koschuh, W.; Povoden, G.; Thang, V.H.; Kromus, S.; Kulbe, K.D.; Novalin, S.; Krotscheck, C. Production of Leaf Protein Concentrate from Ryegrass (Lolium perenne x Multiflorum) and Alfalfa (Medicago sauva Subsp. Sativa). Comparison between Heat Coagulation/Centrifugation and Ultrafiltration. Desalination 2004, 163, 253–259. [Google Scholar] [CrossRef]
- Özköse, A.; Arslan, D.; Acar, A. The Comparison of the Chemical Composition, Sensory, Phenolic and Antioxidant Properties of Juices from Different Wheatgrass and Turfgrass Species. Not. Bot. Horti Agrobo 2016, 44, 499–507. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Open Agrar Repositorium: Greifswald, Germany, 2018. [Google Scholar] [CrossRef]
- Varriale, L.; Hengsbach, J.-N.; Guo, T.; Kuka, K.; Tippkötter, N.; Ulber, R. Sustainable Production of Lactic Acid Using a Perennial Ryegrass as Feedstock—A Comparative Study of Fermentation at the Bench- and Reactor-Scale, and Ensiling. Sustainability 2024, 16, 8054. [Google Scholar] [CrossRef]
- Wei, M.; Xiong, S.; Chen, F.; Geladi, P.; Eilertsen, L.; Myronycheva, O.; Lestander, T.A.; Thyrel, M. Energy Smart Hot-Air Pasteurisation as Effective as Energy Intense Autoclaving for Fungal Preprocessing of Lignocellulose Feedstock for Bioethanol Fuel Production. Renew. Energy 2020, 155, 237–247. [Google Scholar] [CrossRef]
- Xiong, S.; Martín, C.; Eilertsen, L.; Wei, M.; Myronycheva, O.; Larsson, S.H.; Lestander, T.A.; Atterhem, L.; Jönsson, L.J. Energy-Efficient Substrate Pasteurisation for Combined Production of Shiitake Mushroom (Lentinula edodes) and Bioethanol. Bioresour. Technol. 2019, 274, 65–72. [Google Scholar] [CrossRef] [PubMed]
- DSMZ GmbH UNIVERSAL MEDIUM FOR YEASTS (YM). Available online: https://www.dsmz.de/microorganisms/medium/pdf/DSMZ_Medium186.pdf (accessed on 21 January 2025).
- Winkler, K.; Socher, M.L. Shake Flask Technology. In Encyclopedia of Industrial Biotechnology; Wiley: Hoboken, NJ, USA, 2014; pp. 1–16. ISBN 978-0-471-79930-6. [Google Scholar]
- Matlock, B.C.; Beringer, R.W.; Ash, D.; Allen, M.; Page, A.F. Analyzing Differences in Bacterial Optical Density Measurements Between Spectrophotometers; Thermo Fisher Scientific: Waltham, MA, USA, 2011. [Google Scholar]
- Canale, A.; Valente, M.E.; Ciotti, A. Determination of Volatile Carboxylic Acids (C1–C5i ) and Lactic Acid in Aqueous Acid Extracts of Silage by High Performance Liquid Chromatography. J. Sci. Food Agric. 1984, 35, 1178–1182. [Google Scholar] [CrossRef]
- Tix, J.; Moll, F.; Krafft, S.; Betsch, M.; Tippkötter, N. Hydrogen Production from Enzymatic Pretreated Organic Waste with Thermotoga neapolitana. Energies 2024, 17, 2938. [Google Scholar] [CrossRef]
- Myers, J.A.; Curtis, B.S.; Curtis, W.R. Improving Accuracy of Cell and Chromophore Concentration Measurements Using Optical Density. BMC Biophys. 2013, 6, 4. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Fresenius Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Mæhre, H.; Dalheim, L.; Edvinsen, G.; Elvevoll, E.; Jensen, I.-J. Protein Determination—Method Matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef]
- Hayes, M. Measuring Protein Content in Food: An Overview of Methods. Foods 2020, 9, 1340. [Google Scholar] [CrossRef]
- Rocha, S.; Coimbra, M.; Delgadillo, I. Occurrence of Furfuraldehydes during the Processing of Quercus Suber L. Cork. Simultaneous Determination of Furfural, 5-Hydroxymethylfurfural and 5-Methylfurfural and Their Relation with Cork Polysaccharides. Carbohydr. Polym. 2004, 56, 287–293. [Google Scholar] [CrossRef]
- Liu, Z.L.; Slininger, P.J.; Dien, B.S.; Berhow, M.A.; Kurtzman, C.P.; Gorsich, S.W. Adaptive Response of Yeasts to Furfural and 5-Hydroxymethylfurfural and New Chemical Evidence for HMF Conversion to 2,5-Bis-Hydroxymethylfuran. J. Ind. Microbiol. Biotechnol. 2004, 31, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.; Cunha, J.T.; Domingues, L. Establishment of Kluyveromyces marxianus as a Microbial Cell Factory for Lignocellulosic Processes: Production of High Value Furan Derivatives. JoF 2021, 7, 1047. [Google Scholar] [CrossRef]
- Rugthaworn, P.; Murata, Y.; Machida, M.; Apiwatanapiwat, W.; Hirooka, A.; Thanapase, W.; Dangjarean, H.; Ushiwaka, S.; Morimitsu, K.; Kosugi, A.; et al. Growth Inhibition of Thermotolerant Yeast, Kluyveromyces marxianus, in Hydrolysates from Cassava Pulp. Appl. Biochem. Biotechnol. 2014, 173, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.M.; Ballesteros, I.; Negro, M.J.; Manzanares, P.; Cabanas, A.; Ballesteros, M. Effect of Binary Combinations of Selected Toxic Compounds on Growth and Fermentation of Kluyveromyces marxianus. Biotechnol. Prog. 2004, 20, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Grba, S.; Stehlik-Tomas, V.; Stanzer, D.; Vahčić, N.; Škrlin, A. Selection of Yeast Strain Kluyveromyces marxianus for Alcohol and Biomass Production on Whey. Chem. Biochem. Eng. Q. 2002, 16, 13–16. [Google Scholar]
- Sampaio, J.P.; Spencer-Martins, I. Adaptive Growth at High Temperatures of the Lactose-fermenting Yeast Kluyveromyces marxianus Var. Marxianus. J. Basic. Microbiol. 1989, 29, 61–64. [Google Scholar] [CrossRef]
- Rocha, S.N.; Abrahão-Neto, J.; Gombert, A.K. Physiological Diversity within the Kluyveromyces marxianus Species. Antonie Van Leeuwenhoek 2011, 100, 619–630. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Ploidy | Breeding Companies (City, Country) | BBCH | Harvest Time | |
|---|---|---|---|---|---|
| Arvicola | 4× | Freudenberger (Krefeld, Germany) | 65 | 3 June 2022 | |
| Honroso | 2× | DSV (Lippstadt, Germany) | 31 | early cut (E) | 17 May 2022 |
| 41 | late cut (L) | 31 May 2022 | |||
| Explosion | 4× | DSV (Lippstadt, Germany) | 31 | early cut (E) | 17 May 2022 |
| 41 | late cut (L) | 31 May 2022 | |||
| Abbreviation | Samples | Press Juice Dilution | Substance Concentration |
|---|---|---|---|
| PJ | Press juice | Diluted 1:2 with deionized water | - |
| PJ-Y | Press juice + Yeast extract | Yeast extract 3 g·L−1 | |
| PJ-P | Press juice + Peptone | Peptone 5 g·L−1 | |
| PJ-G | Press juice + Glucose | Glucose 10 g·L−1 | |
| PJ-YP | Press juice + Yeast extract + Peptone | Yeast extract 3 g·L−1 Peptone 5 g·L−1 | |
| PJ-YPG | Press juice + Yeast extract + Peptone + Glucose | Yeast extract 3 g·L−1 Peptone 5 g·L−1 Glucose 10 g·L−1 |
| Samples | Growth Rate [h−1] | Maximum Biomass Concentration [g·L−1] | Increase in Biomass |
|---|---|---|---|
| PJ | 0.490 | 9.42 | - |
| PJ-Y | 0.511 | 10.47 | 11% |
| PJ-P | 0.512 | 10.25 | 8.8% |
| PJ-G | 0.479 | 11.58 | 23% |
| PJ-YP | 0.497 | 11.80 | 25% |
| PJ-YPG | 0.422 | 16.00 | 71% |
| Samples | Growth Rate [h−1] | Maximum Biomass Concentration [g·L−1] | Biomass Yield Coefficient [g·g−1] |
|---|---|---|---|
| ExplosionE | 0.483 | 16.62 ± 0.49 | 0.996 ± 0.049 |
| ExplosionL | 0.482 | 15.05 ± 0.90 | 1.174 ± 0.102 |
| HonrosoE | 0.433 | 15.32 ± 0.23 | 0.958 ± 0.038 |
| HonrosoL | 0.429 | 13.79 ± 0.49 | 1.199 ± 0.060 |
| YM Media | 0.460 | 5.22 ± 0.17 | 0.510 ± 0.030 |
| Samples | Field Yield [t WM·ha−1] | Press Juice Yield * [m3·ha−1] | Maximum Biomass Concentration [g·L−1] | Biomass Yield [kg·ha−1] |
|---|---|---|---|---|
| ExplosionE | 15.9 ± 1.9 | 6.8 ± 0.8 | 16.62 ± 0.49 | 113.3 ± 13.9 |
| ExplosionL | 16.9 ± 0.4 | 7.3 ± 0.2 | 15.05 ± 0.90 | 109.3 ± 7.1 |
| HonrosoE | 12.3 ± 0.2 | 5.3 ± 0.1 | 15.32 ± 0.23 | 81.0 ± 1.8 |
| HonrosoL | 14.2 ± 0.2 | 6.1 ± 0.3 | 13.79 ± 0.49 | 84.3 ± 4.8 |
| Samples | Biomass Yield [kg·ha−1] | SCP Content [%] | SCP Yield [kg·ha−1] |
|---|---|---|---|
| ExplosionE | 113.3 ± 13.9 | 19.1 ± 0.9 | 21.6 ± 3.7 |
| ExplosionL | 109.3 ± 7.1 | 18.9 ± 1.2 | 20.7 ± 2.7 |
| HonrosoE | 81.0 ± 1.8 | 18.6 ± 1.8 | 15.0 ± 1.8 |
| HonrosoL | 84.3 ± 4.8 | 20.9 ± 0.9 | 17.6 ± 1.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, T.; Bode, J.; Kuka, K.; Tippkötter, N. Enhancing Single-Cell Protein Yield Through Grass-Based Substrates: A Study of Lolium perenne and Kluyveromyces marxianus. Fermentation 2025, 11, 266. https://doi.org/10.3390/fermentation11050266
Guo T, Bode J, Kuka K, Tippkötter N. Enhancing Single-Cell Protein Yield Through Grass-Based Substrates: A Study of Lolium perenne and Kluyveromyces marxianus. Fermentation. 2025; 11(5):266. https://doi.org/10.3390/fermentation11050266
Chicago/Turabian StyleGuo, Tianyi, Joshua Bode, Katrin Kuka, and Nils Tippkötter. 2025. "Enhancing Single-Cell Protein Yield Through Grass-Based Substrates: A Study of Lolium perenne and Kluyveromyces marxianus" Fermentation 11, no. 5: 266. https://doi.org/10.3390/fermentation11050266
APA StyleGuo, T., Bode, J., Kuka, K., & Tippkötter, N. (2025). Enhancing Single-Cell Protein Yield Through Grass-Based Substrates: A Study of Lolium perenne and Kluyveromyces marxianus. Fermentation, 11(5), 266. https://doi.org/10.3390/fermentation11050266