
Use of Lachancea thermotolerans for the Bioacidification of White Grape Musts: Assays from the Bench to the Cellar Scale

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bench Scale Fermentations
2.2. Cellar Scale Experiments
2.2.1. Chasselas Vinification
2.2.2. Divona Vinification
2.3. Flow Cytometry (FCM)
2.4. Wine Analysis by Enzymatic Methods and Fourier Transform Infrared (FT-IR)
2.4.1. Bench Scale Fermentations
2.4.2. Cellar Scale Assays
2.5. Statistical Analysis
3. Results
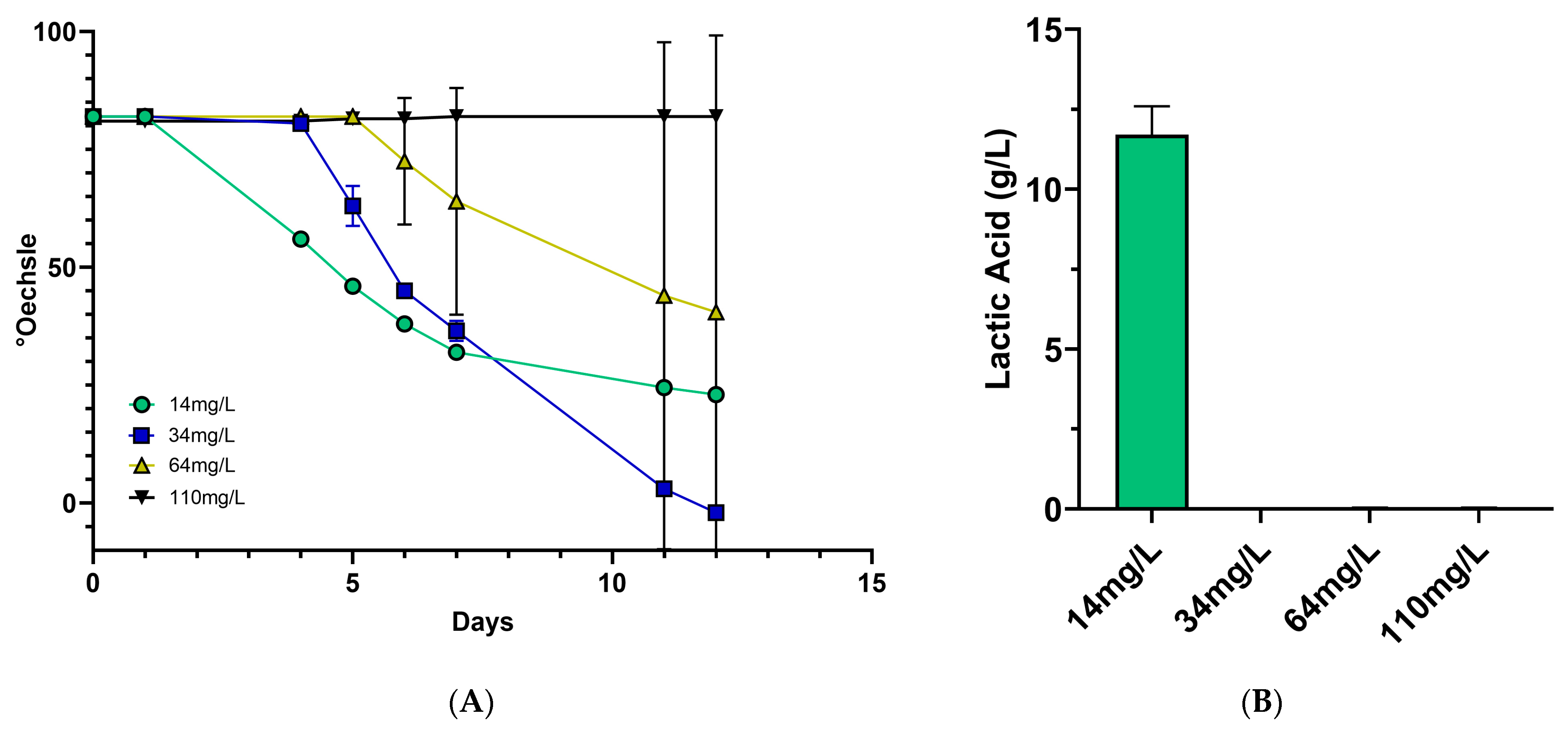
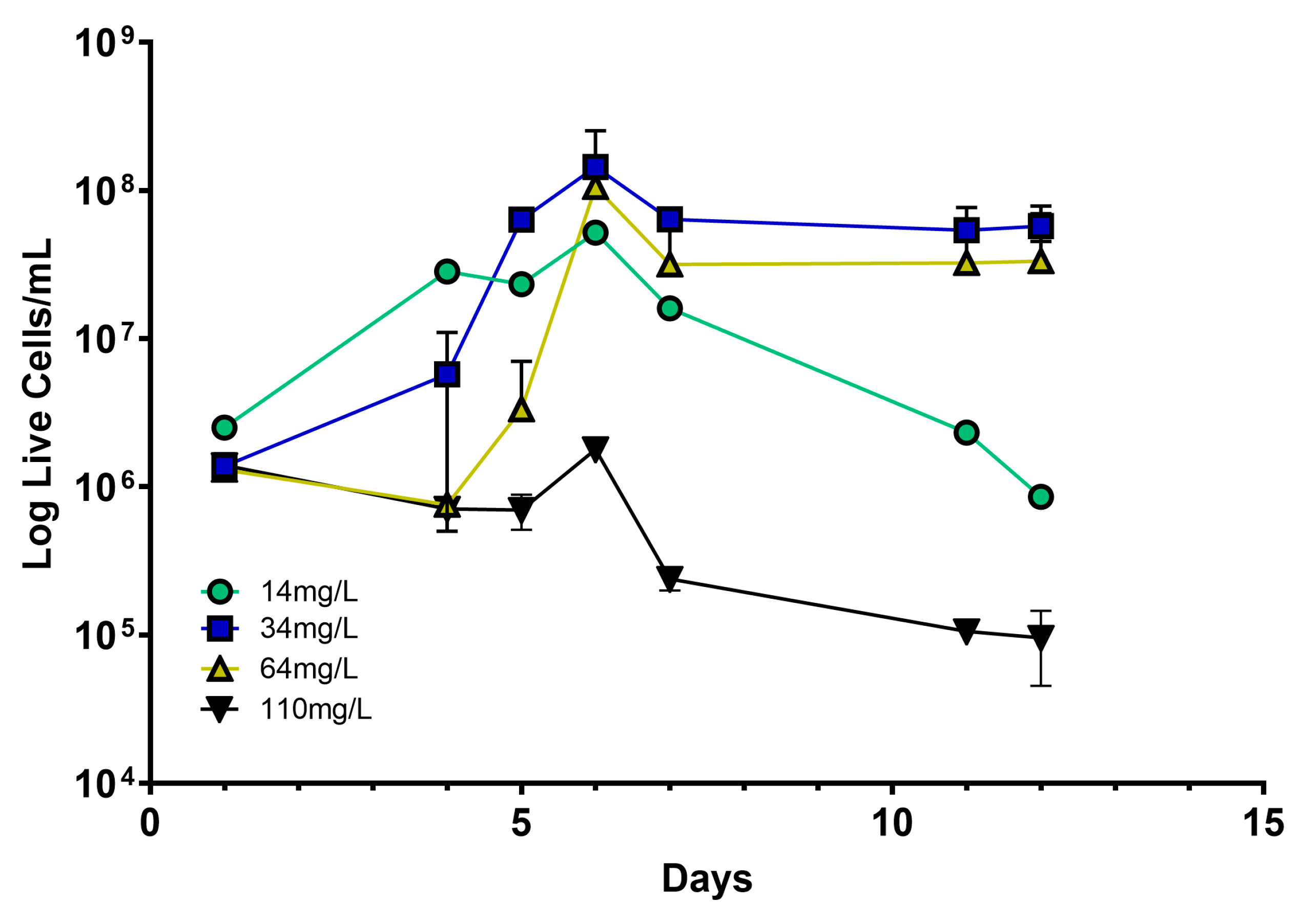
3.1. Increasing Sulfite Concentration Prevents Lt Development and LA Production
3.2. Nitrogen Level Does Not Affect LA Production in Bench Scale Experiments
3.3. Plateau of LA Production Reached after Three Days
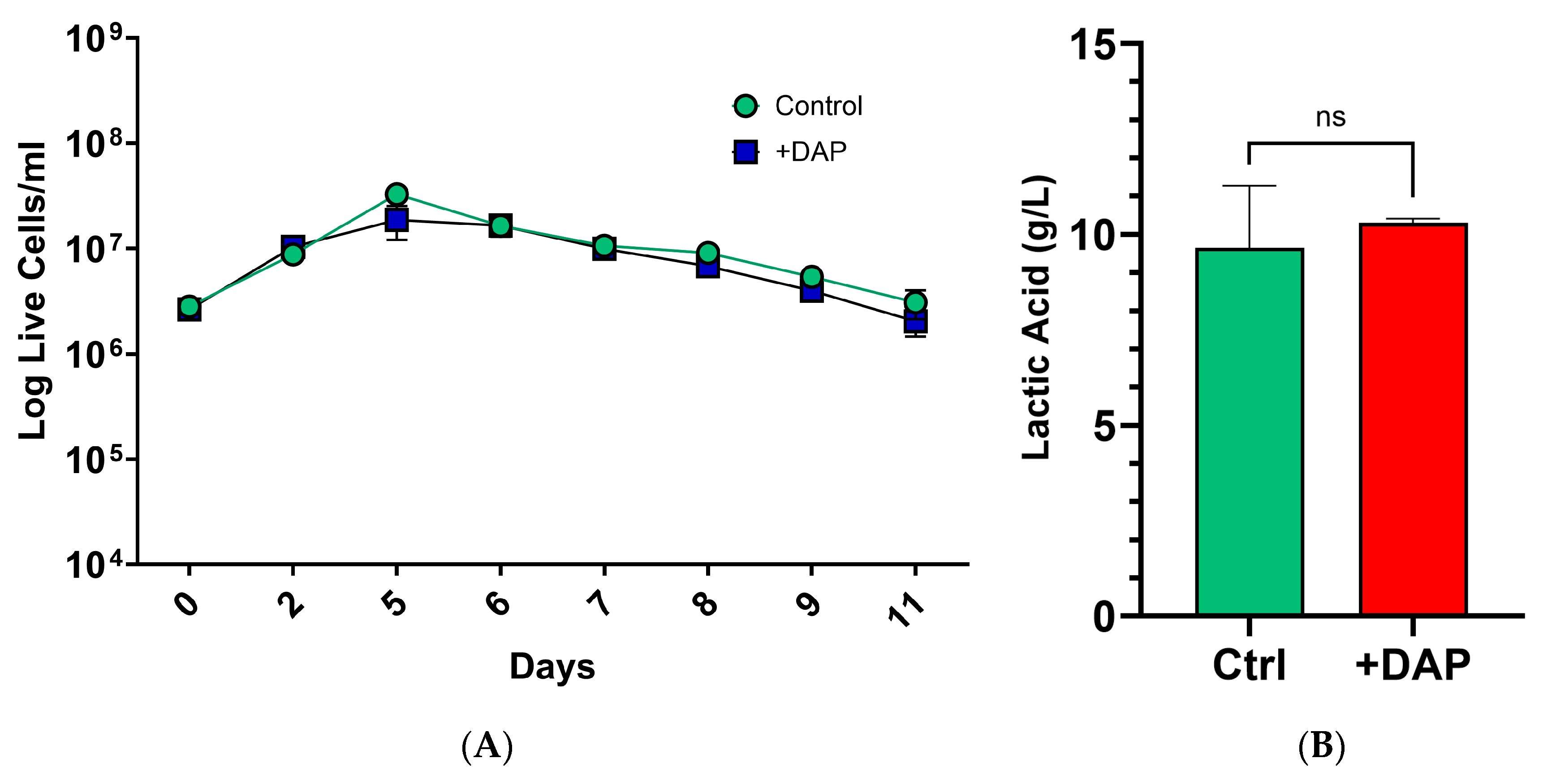
3.4. Time-Dependent Sequential Sc Inoculation Protocols Minimally Affect LA Production by Lt
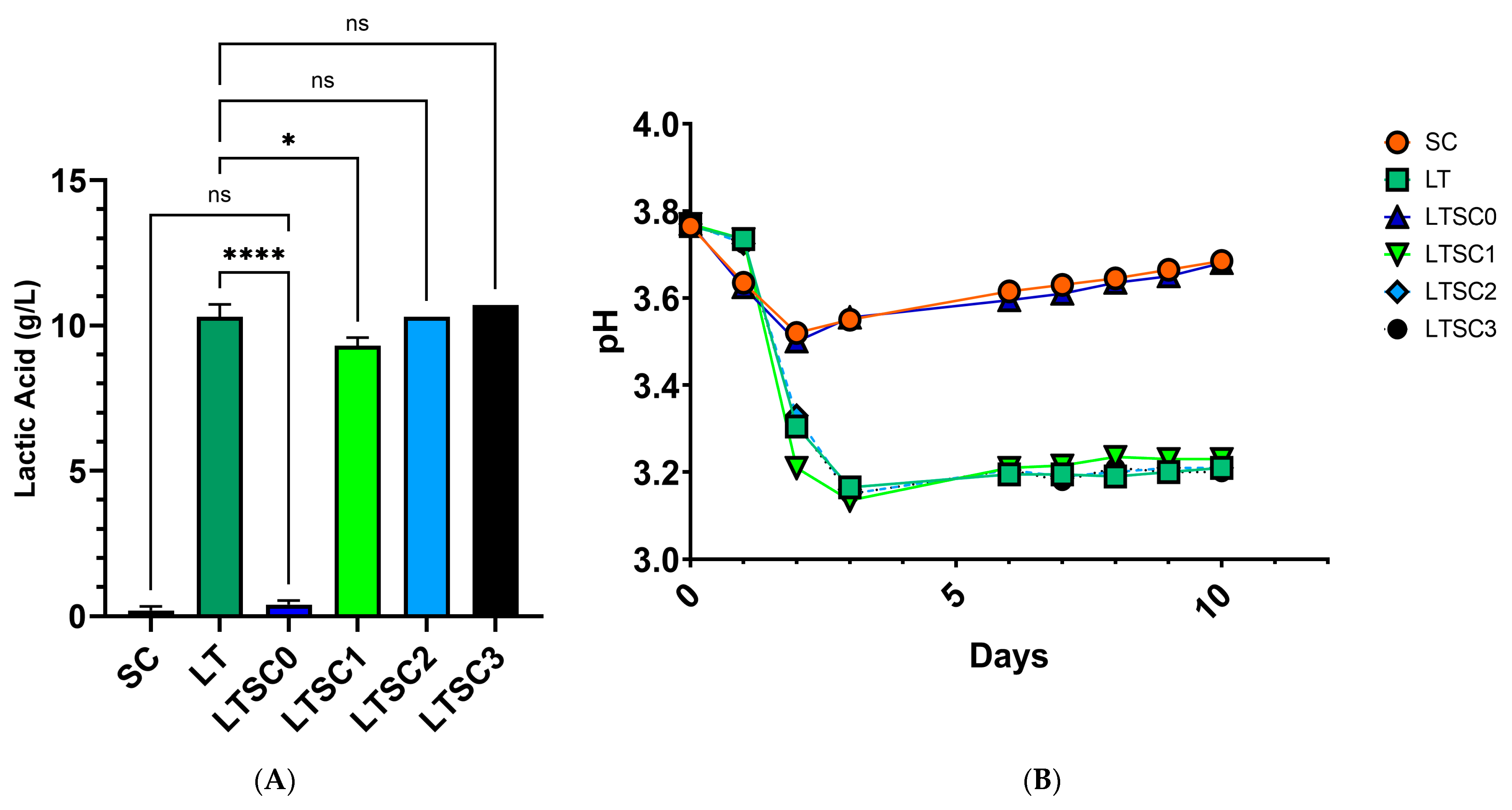
3.5. Alternative Sc Inoculation Protocols Provide Acceptable Acidification in Bench-Scale Fermentation
3.6. Cellar Scale Assays Did Not Show Consistent Acidification When Using 12 h Seq and Mixing Protocols
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rogiers, S.Y.; Greer, D.H.; Liu, Y.; Baby, T.; Xiao, Z. Impact of climate change on grape berry ripening: An assessment of adaptation strategies for the Australian vineyard. Front. Plant Sci. 2022, 13, 1094633. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; González, C.; Escott, C. Non-Saccharomyces as biotools to control the production of off-flavors in wines. Molecules 2021, 26, 4571. [Google Scholar] [CrossRef]
- Blackford, M.; Roesle-Fuchs, J.; Rienth, M.; Dienes-Nagy, A.; Cleroux, M.; Spring, J.L.; Bourdin, G.; Lorenzini, F. Evolution des acides dans les baies et dans les mouts au cours des dernieres millésimes. Rev. Suisse Vitic. Arboric. Hortic. 2019, 51, 290–299. [Google Scholar]
- Payan, C.; Gancel, A.L.; Jourdes, M.; Christmann, M.; Teissedre, P.L. Wine acidification methods: A review. Oeno One 2023, 57, 113–126. [Google Scholar] [CrossRef]
- Vecchio, R.; Parga-Dans, E.; Gonzàlez, P.A.; Annunziata, A. Why consumers drink natural wine? Consumer perception and information about natural wine. Agric. Food Econ. 2021, 9, 22. [Google Scholar] [CrossRef]
- Vicente, J.; Baran, Y.; Navascués, E.; Santos, A.; Calderon, F.; Marquina, D.; Rauhut, D.; Benito, S. Biological management of acidity in wine industry: A review. Int. J. Food Microbiol. 2022, 375, 109726. [Google Scholar] [CrossRef]
- Comuzzo, P.; Del Fresno, J.M.; Voce, S.; Loira, I.; Morata, A. Emerging biotechnologies and non-thermal technologies for winemaking in a context of global warming. Front. Microbiol. 2023, 14, 1273940. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Wendu, T.; Bañuelos, M.A.; González, C.; Suárez Lepe, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef]
- Vicente, J.; Navascués, E.; Calderon, F.; Santos, A.; Marquina, D.; Benito, S. An integrative view of the role of Lachancea thermotolerans in wine technology. Foods 2021, 10, 2878. [Google Scholar] [CrossRef]
- Furet-Gavallet, C. Les levures qui acidifient les vins sont imprévisibles. Vitisphere 2022. [Google Scholar]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Sainz, F.; Pardo, J.; Ruiz, A.; Exposito, D.; Armero, R.; Querol, A.; Guillamon, J.M. Use of non-conventional yeasts to increase total acidity in the Cava base wines. LWT—Food Sci. Technol. 2022, 158, 113183. [Google Scholar] [CrossRef]
- Li, J.; Yuan, M.; Meng, N.; Li, H.; Sun, J.; Sun, B. Influence of nitrogen status on fermentation performances of non-Saccharomyces yeasts: A review. Food Sci. Hum. Wellness 2024, 13, 556–567. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Benito, A.; Calderón, F.; Palomero, F.; Benito, S. Quality and composition of Airén wines fermented by sequential inoculation of Lachancea thermotolerans and Saccharomyces cerevisiae. Food Technol. Biotechnol. 2016, 54, 135–144. [Google Scholar] [CrossRef]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.P.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on chemical composition and sensory profiles of Viognier wines. J. Fungi 2022, 8, 474. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Puyo, M.; Simonin, S.; Bach, B.; Klein, G.; Alexandre, H.; Tourdot-Maréchal, R. Bio-protection in oenology by Metschnikowia pulcherrima: From field results to scientific inquiry. Front. Microbiol. 2023, 14, 1252973. [Google Scholar] [CrossRef]
- Simonin, S.; Honoré-Chedozeau, C.; Monnin, L.; David-Vaizant, V.; Bach, B.; Alexandre, H.; Chatelet, B.; Tourdot-Maréchal, R. Bioprotection on Chardonnay grape: Limits and impacts of settling parameters. Aust. J. Grape Wine Res. 2022, 2022, 1489094. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. Metschnikowia pulcherrima in cold clarification: Biocontrol activity and aroma enhancement in Verdicchio wine. Fermentation 2023, 9, 302. [Google Scholar] [CrossRef]
- Sizzano, F.; Blackford, M.; Berthoud, H.; Amiet, L.; Bailly, S.; Vuichard, F.; Monnard, C.; Bieri, S.; Spring, J.L.; Barth, Y.; et al. Bioprospecting of a Metschnikowia pulcherrima indigenous strain for Chasselas winemaking in 2022 vintage. Foods 2023, 12, 4485. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Escott, C.; Vaquero, C.; del Fresno, J.M.; Guamis, B.; Loira, I.; Bañuelos, M.A.; Palomero, F.; Lopez, C.; Gonzaléz, C.; et al. Improving the implantation of non-Saccharomyces yeasts in winemaking by UHPH processing. In Proceedings of the BIO Web of Conferences, 44th World Congress of Vine and Wine, Jerez, Cadiz, Spain, 5–9 June 2023; Volume 68, p. 02001. [Google Scholar]
- Guindal, A.M.; Gonzalez, R.; Tronchoni, J.; Roodink, J.S.; Morales, P. Directed evolution of Saccharomyces cerevisiae for low volatile acidity during winemaking under aerobic conditions. Food Microbiol. 2023, 114, 104282. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Densitometry (°Oe) | pH | Sugars (g/L) | EtOH (% v/v) | Titratable Acidity (g/L) | Lactic Acid (g/L) | Malic Acid (g/L) | Tartaric Acid (g/L) | YAN (mg/L) | |
|---|---|---|---|---|---|---|---|---|---|
| Control (Sc) | 74/<0 | 3.70/3.65 | ND/<1 | ND/10.4 | 4.50/5.12 | 0.3/1.07 | 1.70/0.62 | ND/1.89 | 283/ND |
| Bioac. (Lt + Sc) | 78/<0 | 3.73/3.66 | ND/<1 | ND/11.1 | 4.35/5.33 | 0.23/1.14 | 1.90/0.81 | ND/1.89 | 310/ND |
| Densitometry (°Oe) | pH | Sugars (g/L) | EtOH (% v/v) | Titratable Acidity (g/L) | Lactic Acid (g/L) | Malic Acid (g/L) | Tartaric Acid (g/L) | YAN (mg/L) | |
|---|---|---|---|---|---|---|---|---|---|
| Control (Sc) | 93/<0 | 3.40/3.44 | 233.3 < 1 | ND/13.5 | 4.7/5.5 | 0/0 | 0.8/0.8 | 6.1/4.5 | 303.2/ND |
| Bioac. (Lt + Sc) | 93/<0 | 3.40/3.42 | 234.2 < 1 | ND/13.4 | 4.7/5.7 | 0/0.03 | 0.80.88 | 6.0/4.4 | 301.5/ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sizzano, F.; Bianconi, V.; Blackford, M.; Bieri, S.; Vuichard, F.; Monnard, C.; Amiet, L.; Spring, J.-L.; Dorsaz, E.; Pfenninger-Bridy, N.; et al. Use of Lachancea thermotolerans for the Bioacidification of White Grape Musts: Assays from the Bench to the Cellar Scale. Fermentation 2024, 10, 458. https://doi.org/10.3390/fermentation10090458
Sizzano F, Bianconi V, Blackford M, Bieri S, Vuichard F, Monnard C, Amiet L, Spring J-L, Dorsaz E, Pfenninger-Bridy N, et al. Use of Lachancea thermotolerans for the Bioacidification of White Grape Musts: Assays from the Bench to the Cellar Scale. Fermentation. 2024; 10(9):458. https://doi.org/10.3390/fermentation10090458
Chicago/Turabian StyleSizzano, Federico, Valentina Bianconi, Marie Blackford, Stefan Bieri, Frédéric Vuichard, Christine Monnard, Laurent Amiet, Jean-Laurent Spring, Eddy Dorsaz, Nadine Pfenninger-Bridy, and et al. 2024. "Use of Lachancea thermotolerans for the Bioacidification of White Grape Musts: Assays from the Bench to the Cellar Scale" Fermentation 10, no. 9: 458. https://doi.org/10.3390/fermentation10090458
APA StyleSizzano, F., Bianconi, V., Blackford, M., Bieri, S., Vuichard, F., Monnard, C., Amiet, L., Spring, J.-L., Dorsaz, E., Pfenninger-Bridy, N., Simonin, S., Bach, B., & Bourdin, G. (2024). Use of Lachancea thermotolerans for the Bioacidification of White Grape Musts: Assays from the Bench to the Cellar Scale. Fermentation, 10(9), 458. https://doi.org/10.3390/fermentation10090458