
Influence of Fermented Mulberry Leaves as an Alternative Animal Feed Source on Product Performance and Gut Microbiome in Pigs

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fermented Mulberry Leaves
2.2. Experimental Design, Animals, and Diets
2.3. Slaughter Procedure, Sample Collection, and Processing
2.4. Determination of Product Performance
2.5. Chemical Composition Determination
2.6. Antioxidant Index Determination
2.7. Economic Benefit Calculations
2.8. Gene Sequencing
2.9. Odorous Compound Determination
2.10. Data Analysis
3. Results
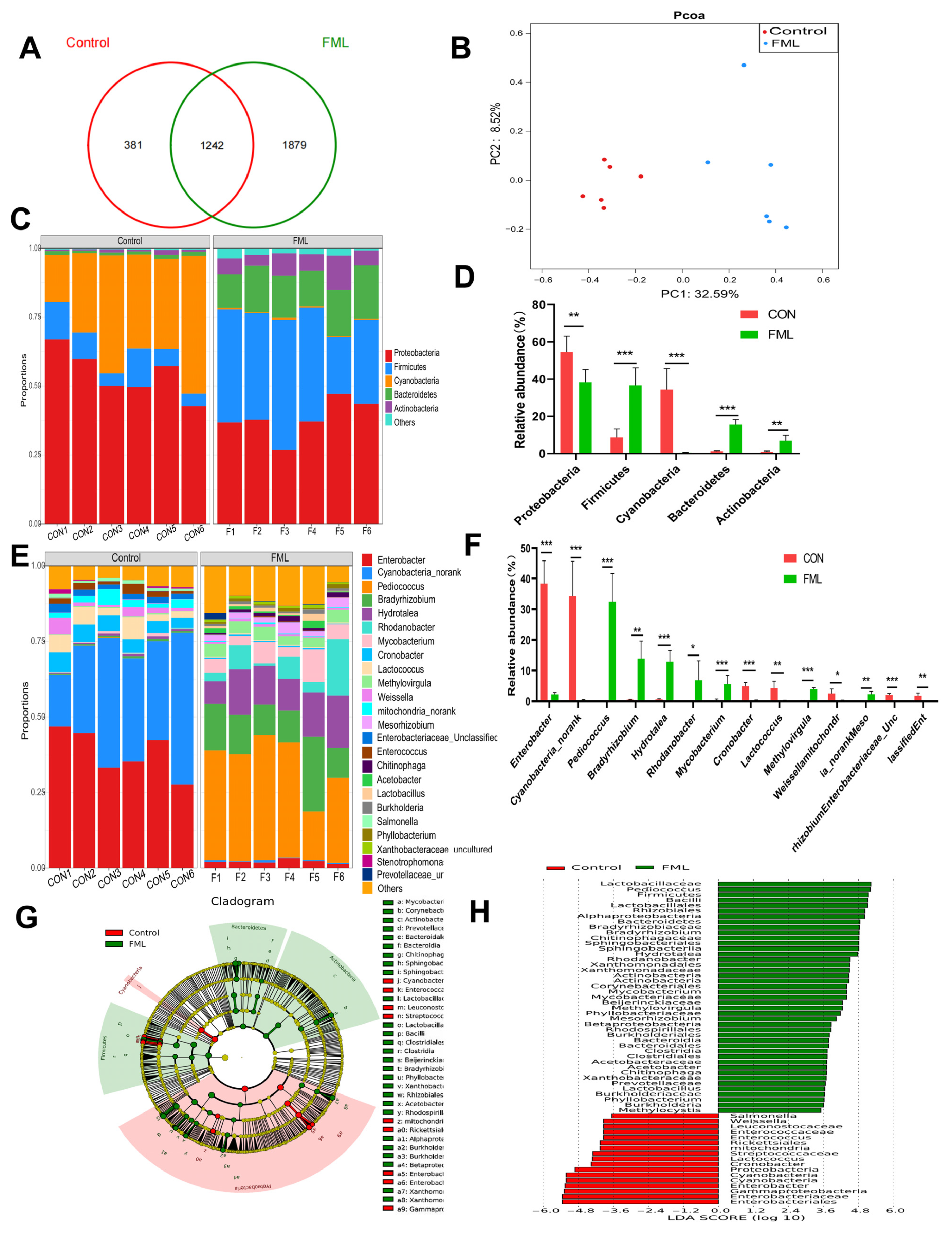
3.1. Fermented Mulberry Leaves
3.1.1. Overview of the Microbial Community in FML
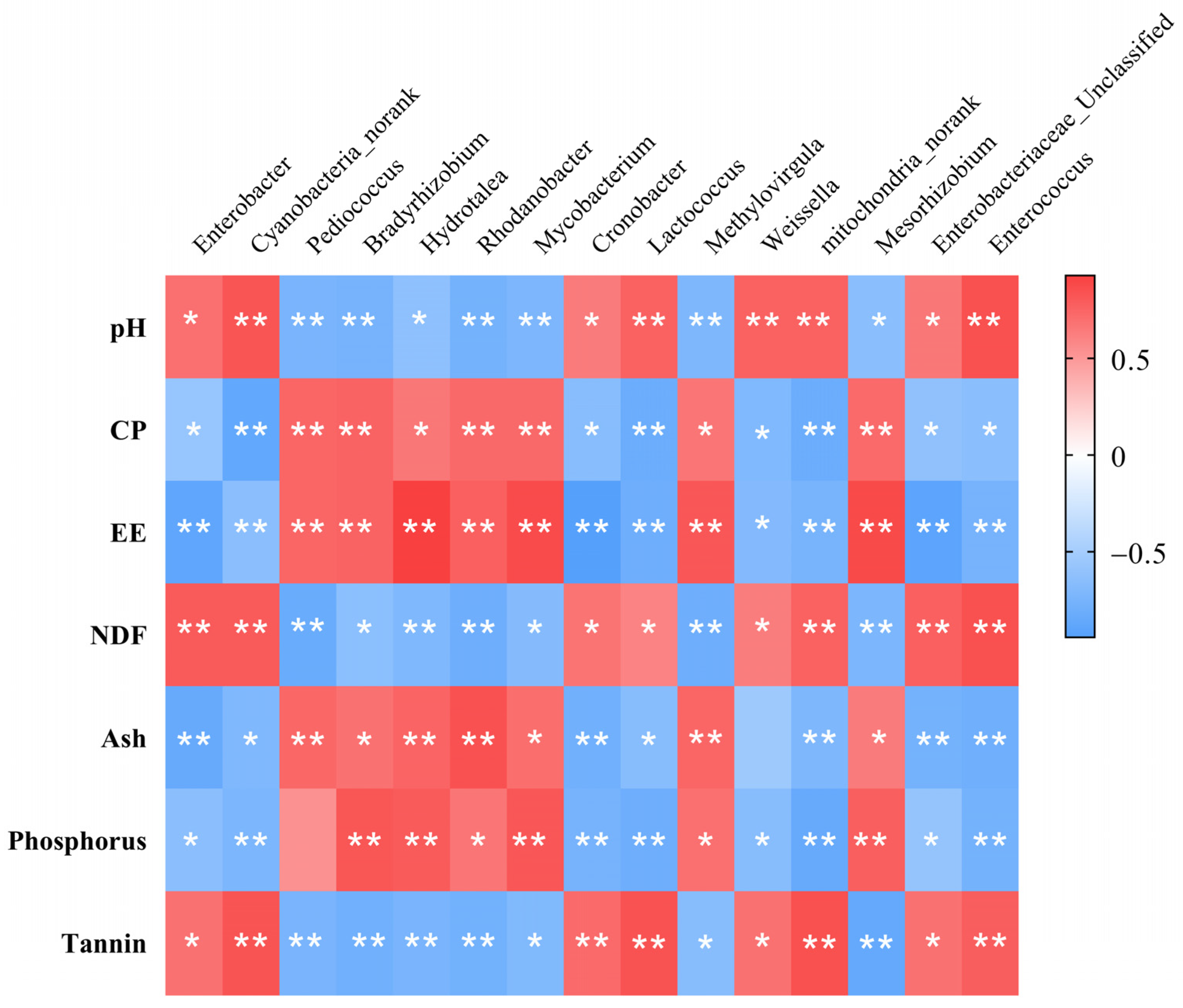
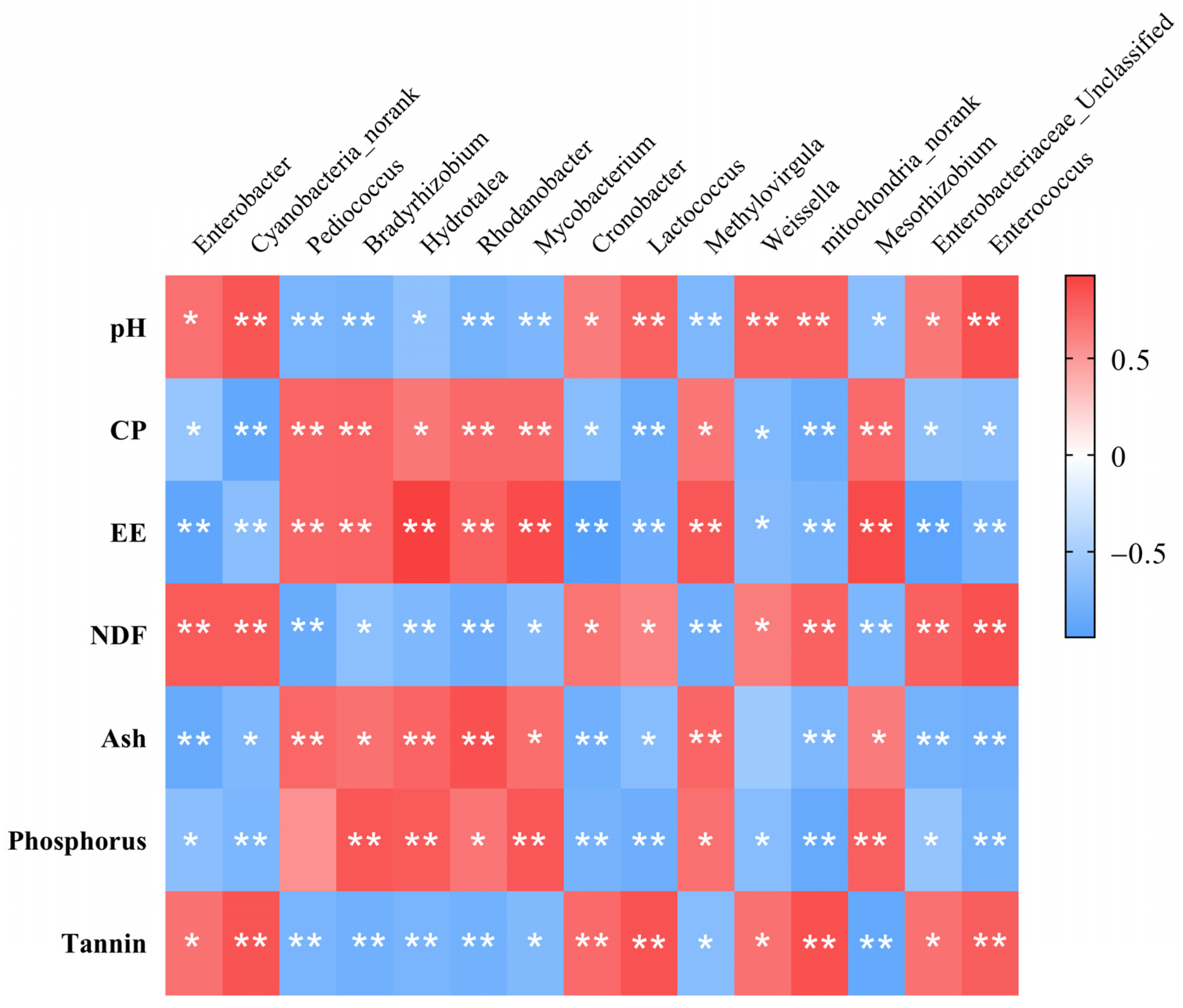
3.1.2. Spearman’s Correlation Analysis
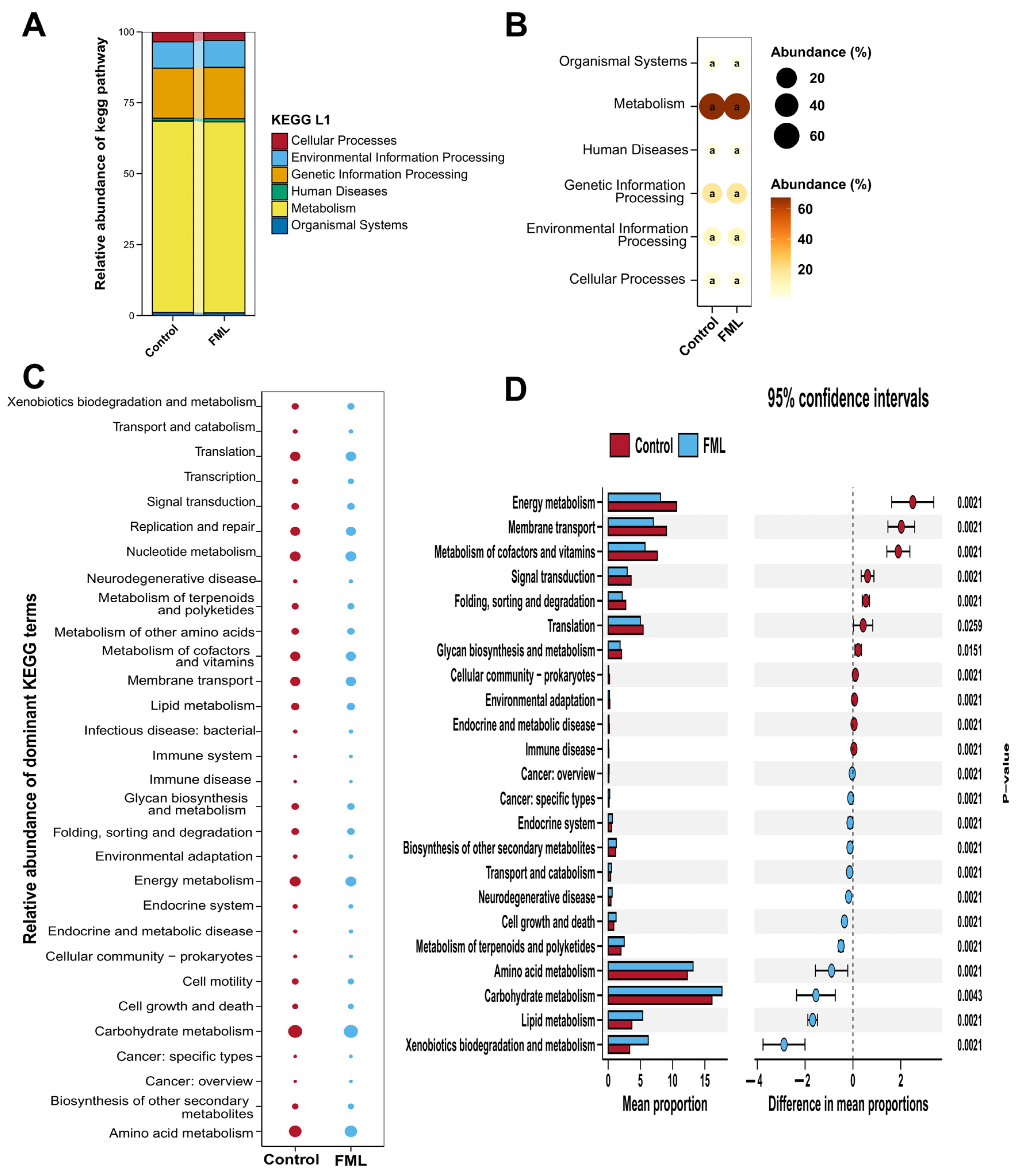
3.1.3. Prediction of the Microbial Community Functions in FML
3.2. Carcass Performance and Meat Quality
3.3. Muscle Chemical Composition and Antioxidant Capacity
3.4. Growth Performance and Economic Benefits
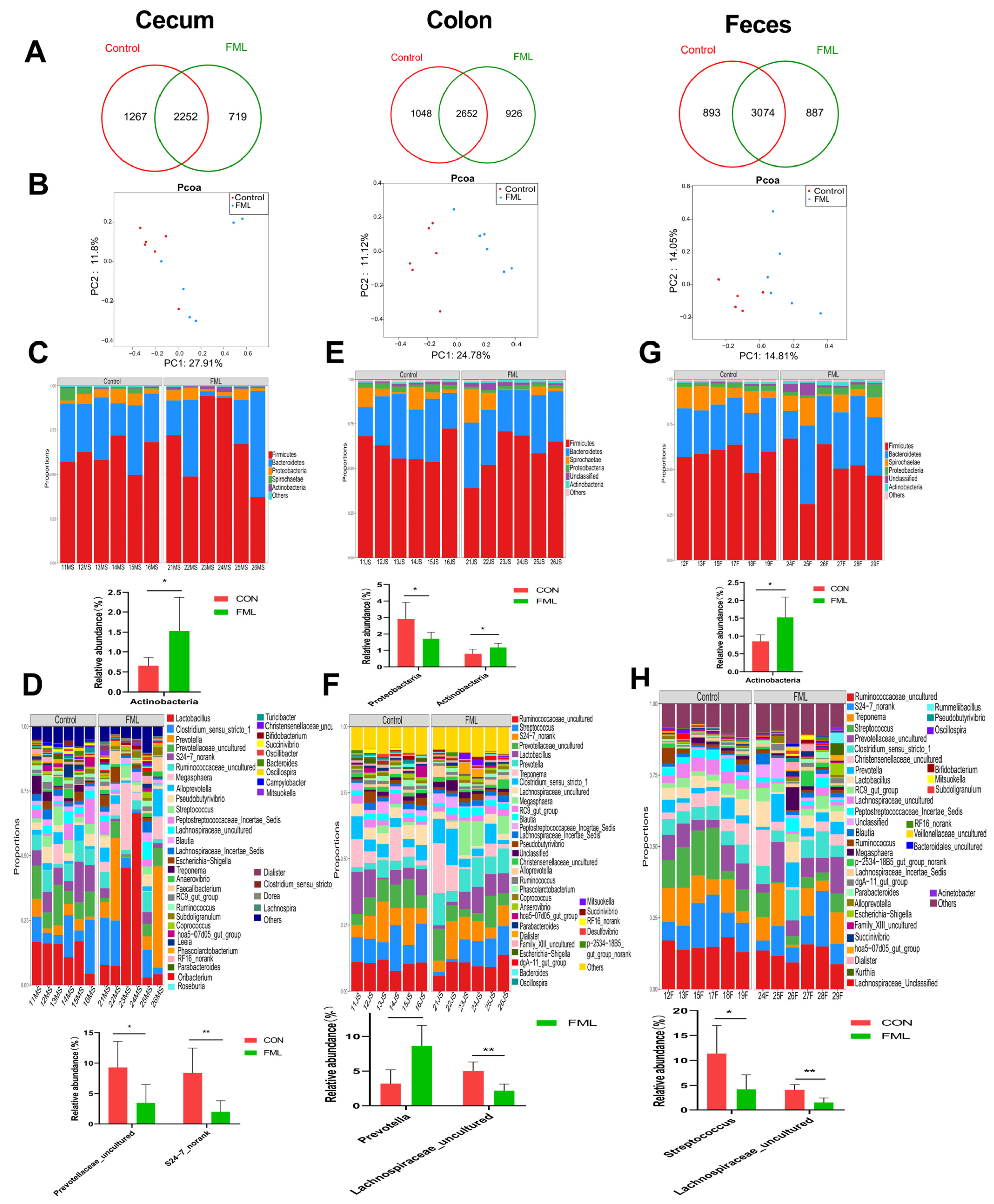
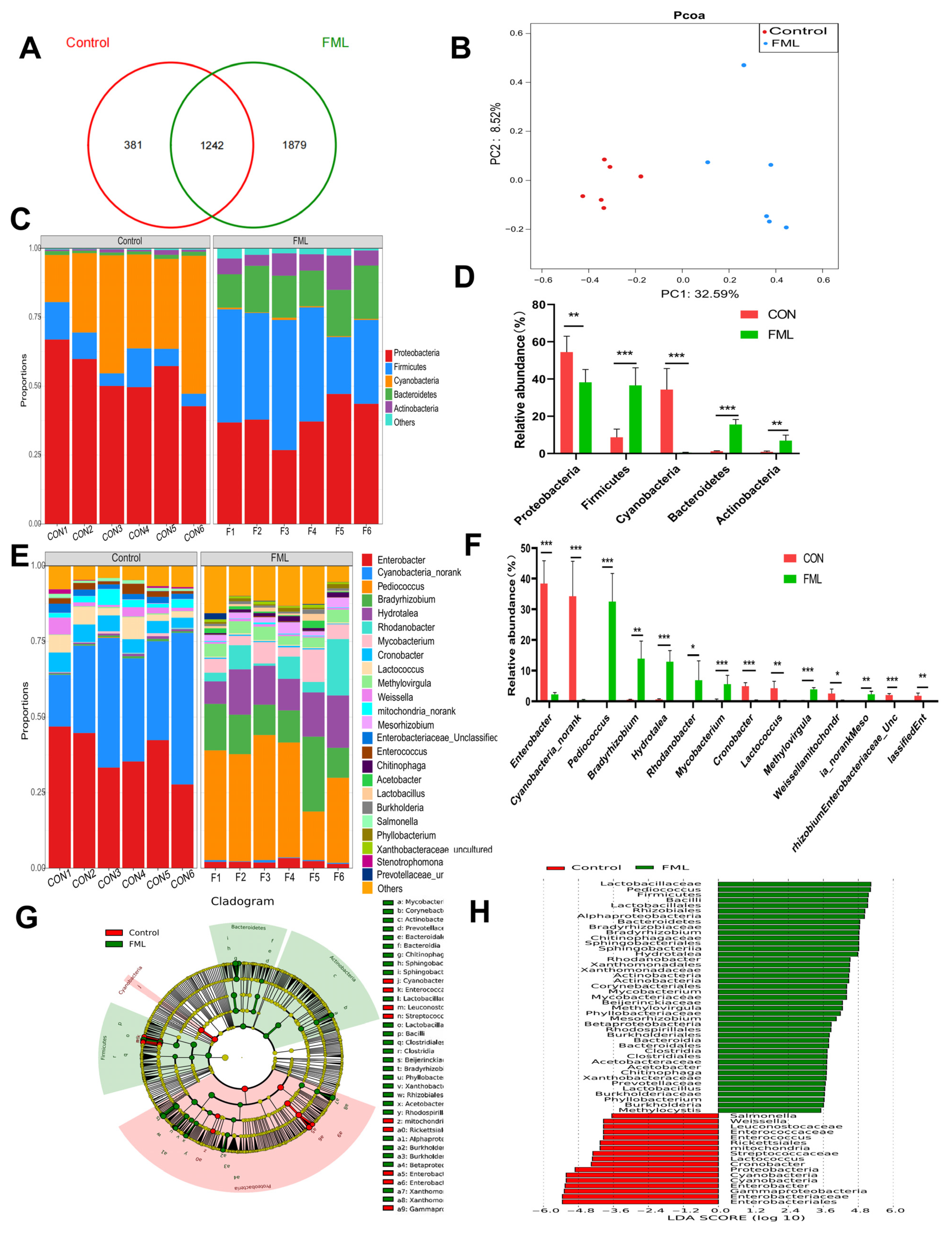
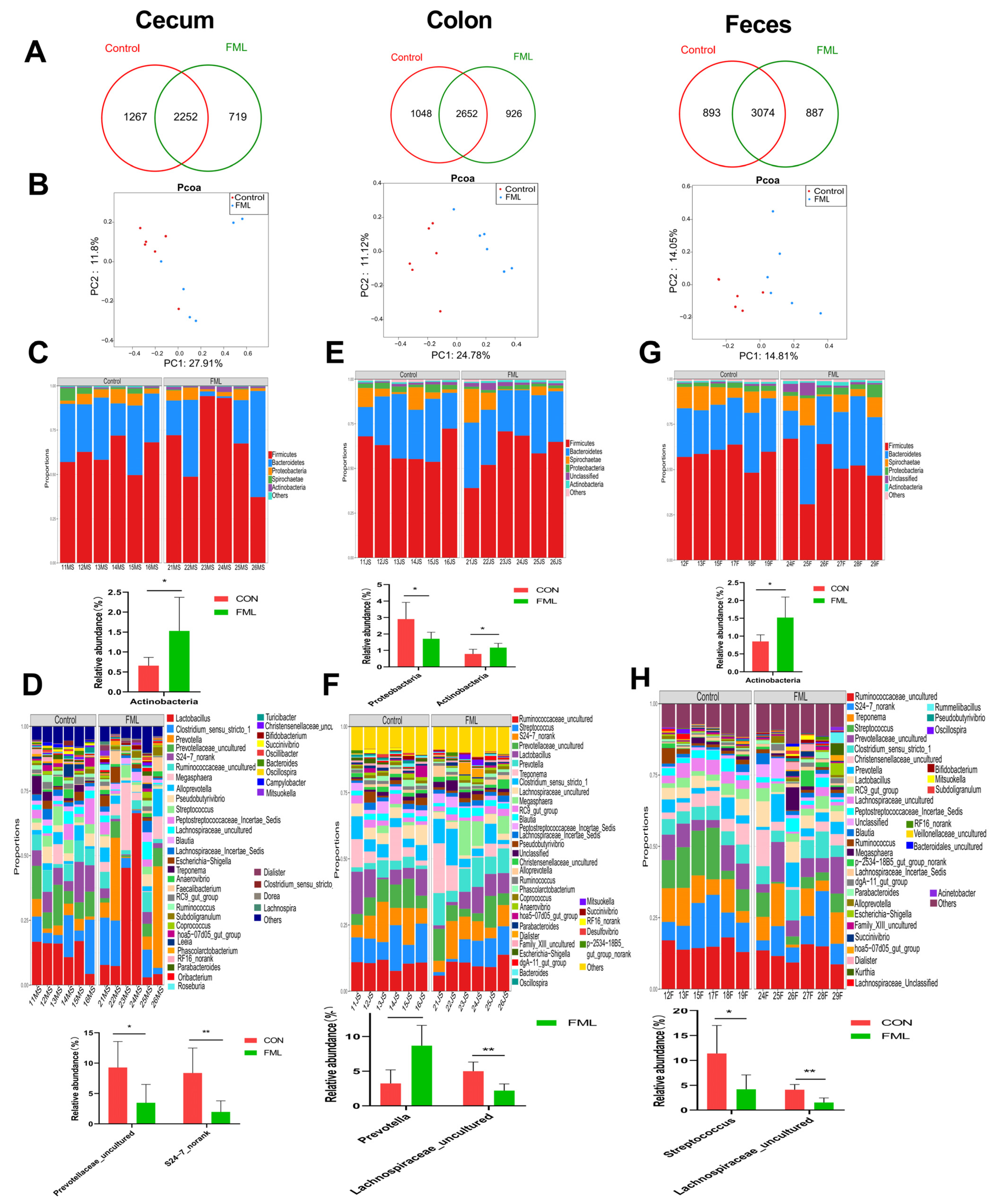
3.5. Overview of the Gut Microbial Community
3.6. LEfSe Analysis
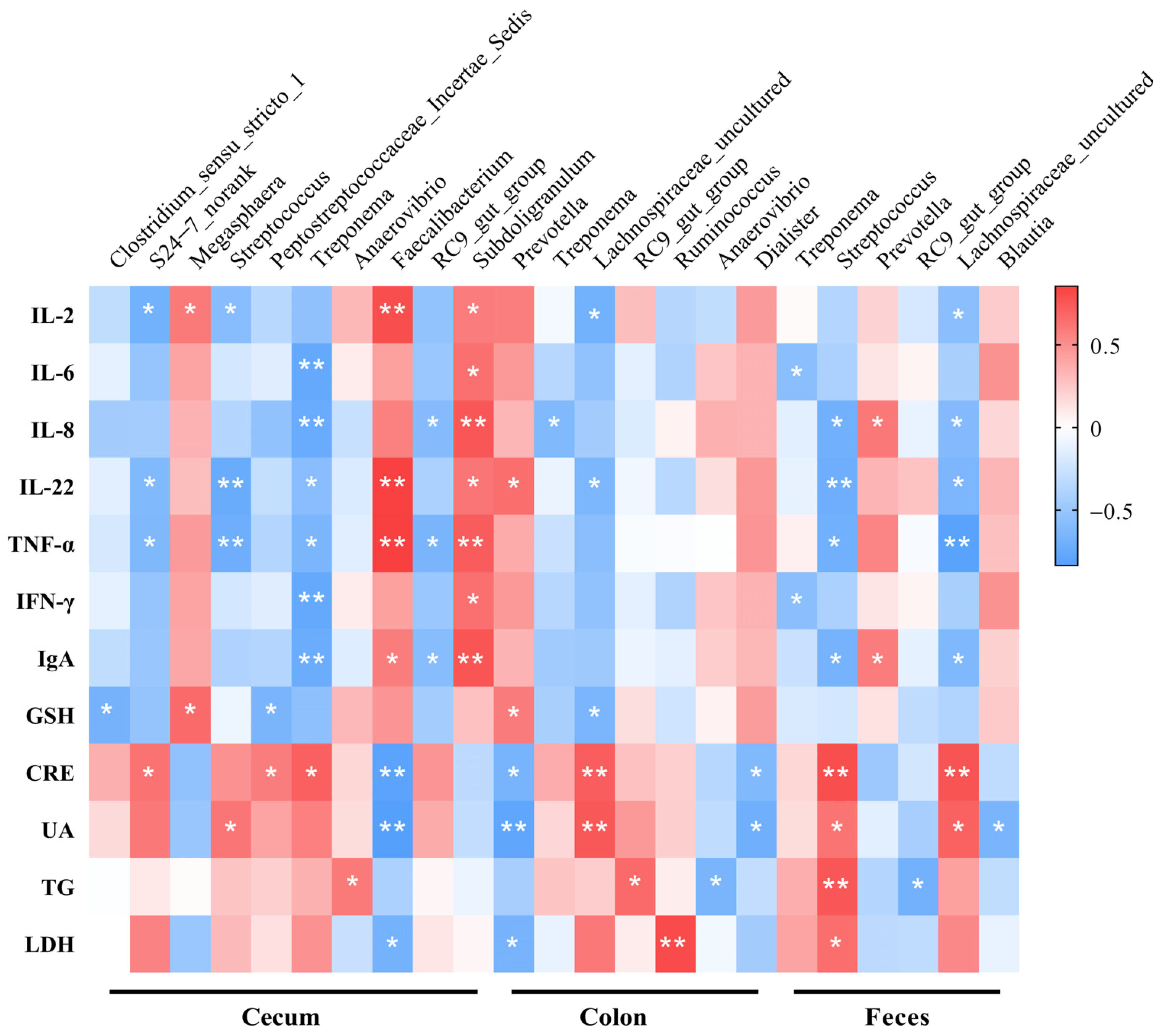
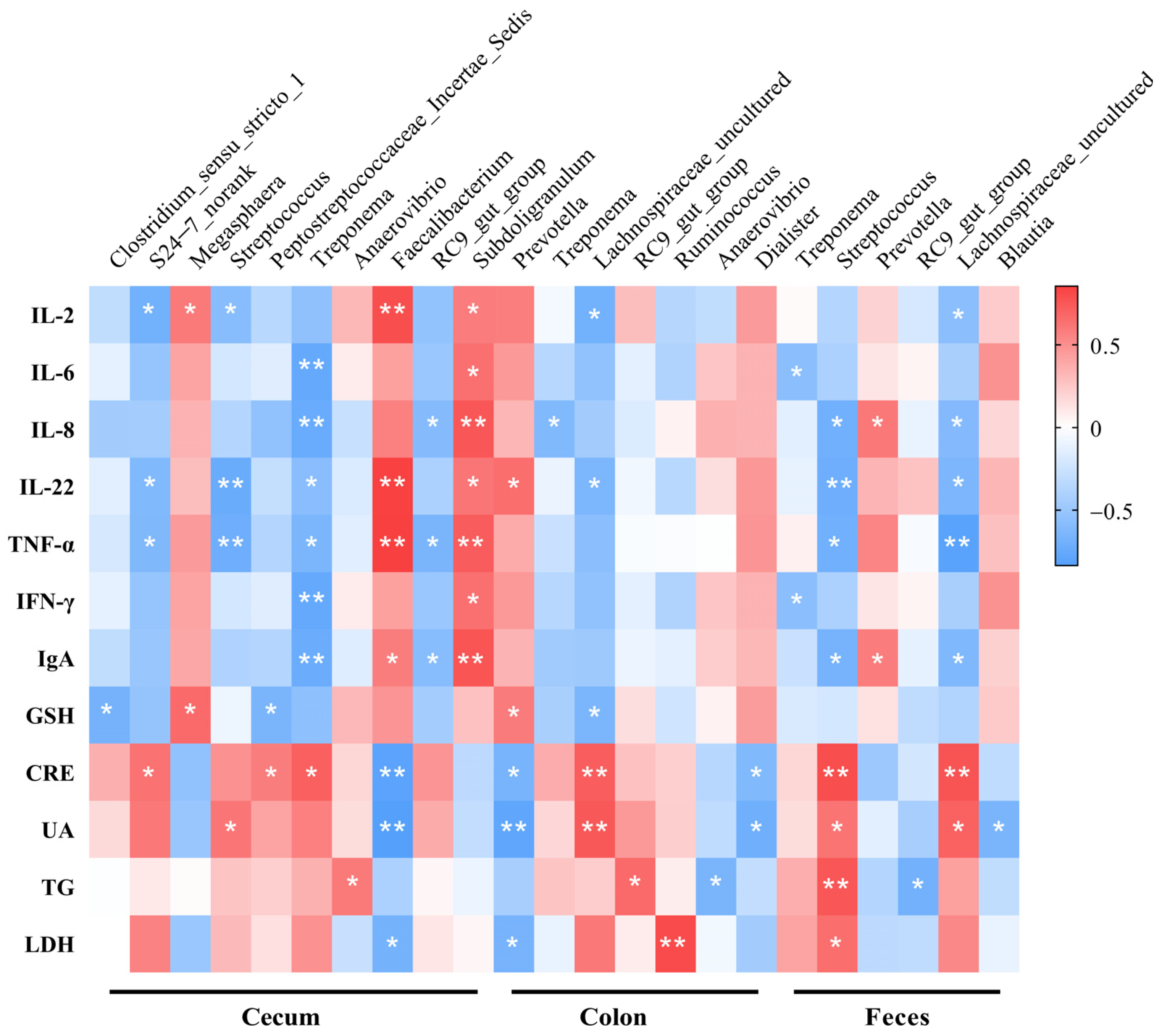
3.7. Spearman’s Correlations between Microorganisms and Blood Parameters
3.8. Prediction of Microbial Community Function in the Gut
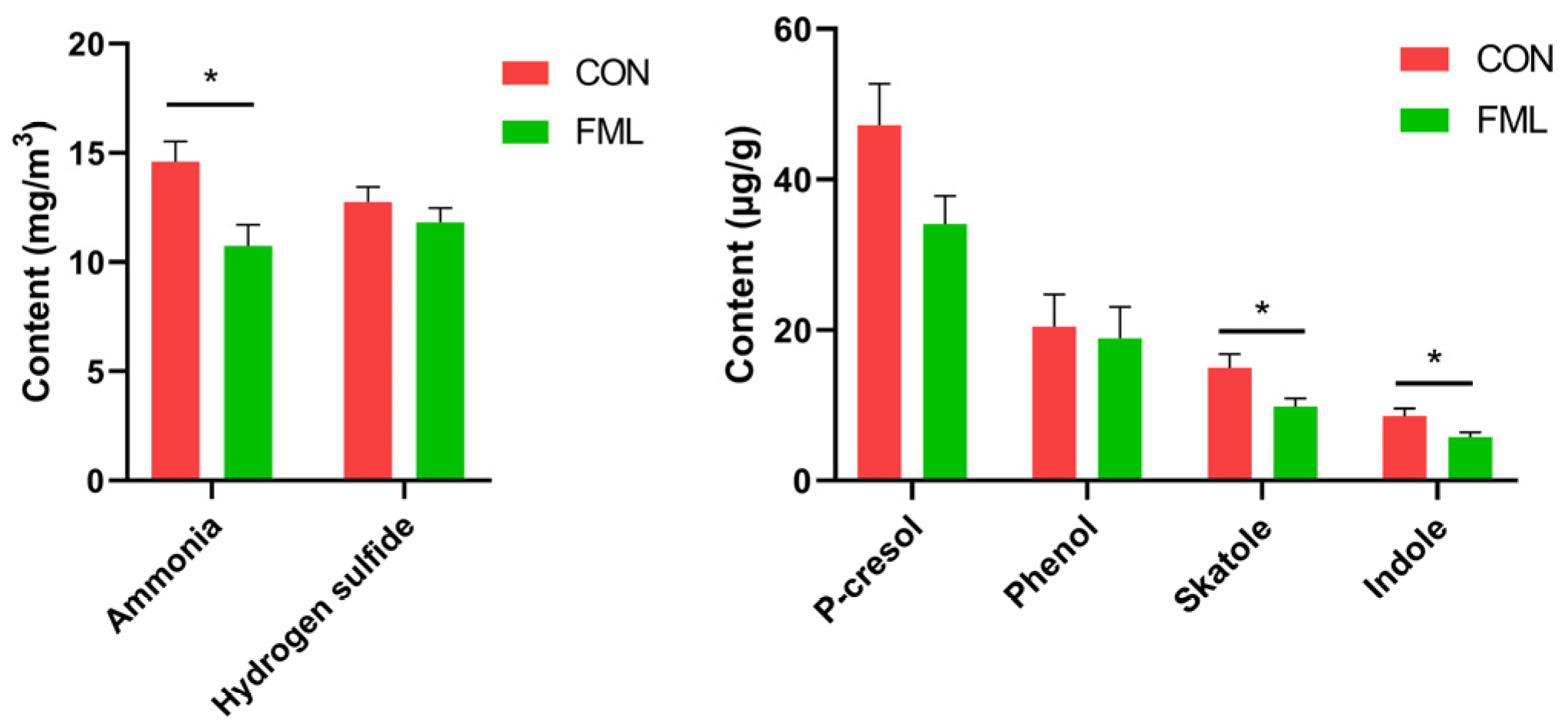
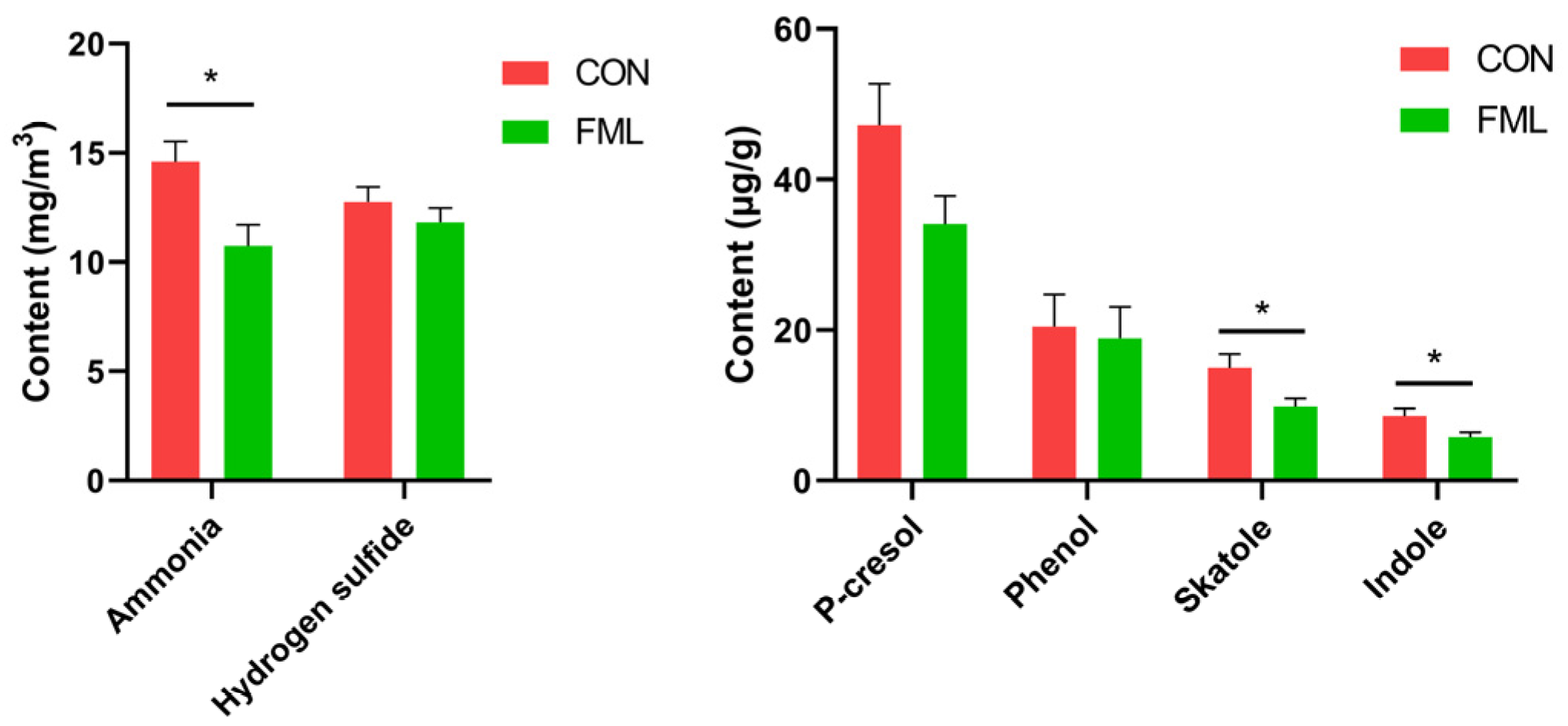
3.9. Odorous Compounds
4. Discussion
4.1. Fermented Mulberry Leaves
4.2. Carcass Performance and Meat Quality
4.3. Muscle Chemical Composition and Antioxidant Capacity
4.4. Economic Benefits
4.5. Gut Microbial Community
4.6. Odorous Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ali, B.M.; van Zanten, H.H.E.; Berentsen, P.; Bastiaansen, J.W.M.; Bikker, P.; Lansink, A.O. Environmental and economic impacts of using co-products in the diets of finishing pigs in Brazil. J. Clean. Prod. 2017, 162, 247–259. [Google Scholar] [CrossRef]
- Hassan, F.; Arshad, M.A.; Li, M.; Rehman, M.S.; Loor, J.J.; Huang, J. Potential of mulberry leaf biomass and its flavonoids to improve production and health in ruminants: Mechanistic insights and prospects. Animals 2020, 10, 2076. [Google Scholar] [CrossRef]
- Ma, G.; Chai, X.; Hou, G.; Zhao, F.; Meng, Q. Phytochemistry, bioactivities and future prospects of mulberry leaves: A review. Food Chem. 2022, 372, 131335. [Google Scholar] [CrossRef]
- Bartkiene, E.; Mozuriene, E.; Lele, V.; Zokaityte, E.; Gruzauskas, R.; Jakobsone, I.; Juodeikiene, G.; Ruibys, R.; Bartkevics, V. Changes of bioactive compounds in barley industry by-products during submerged and solid state fermentation with antimicrobial Pediococcus acidilactici strain LUHS29. Food Sci. Nutr. 2020, 8, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.Y.; Peng, S.; Li, Z.M.; Song, M.; Yu, M.; Deng, D.; Tian, Z.M.; Rong, T.; Liu, Z.C.; Ma, X.Y. Effects of fermented mulberry leaves on growth performance, plasma and urine metabolites of finishing pigs. Chin. J. Anim. Nutr. 2022, 34, 5620–5629. [Google Scholar]
- Tian, S.; Wang, M.; Liu, C.; Zhao, H.; Zhao, B. Mulberry leaf reduces inflammation and insulin resistance in type 2 diabetic mice by TLRs and insulin Signalling pathway. BMC Complem. Altern. Med. 2019, 19, 326. [Google Scholar] [CrossRef]
- Oku, T.; Yamada, M.; Nakamura, M.; Sadamori, N.; Nakamura, S. Inhibitory effects of extractives from leaves of Morus alba on human and rat small intestinal disaccharidase activity. Br. J. Nutr. 2006, 95, 933–938. [Google Scholar] [CrossRef]
- Cai, M.; Mu, L.; Wang, Z.; Liu, J.; Liu, T.; Wanapat, M.; Huang, B. Assessment of mulberry leaf as a potential feed supplement for animal feeding in P.R. China. Asian-Australas J. Anim. Sci. 2019, 32, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Peng, Y.; Wu, D.; Hu, J.; Shi, X.E.; Yang, G.; Li, X. Morus nigra L. leaves improve the meat quality in finishing pigs. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1904–1911. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish. Immun. 2019, 87, 820–828. [Google Scholar] [CrossRef]
- Peng, W.; Talpur, M.Z.; Zeng, Y.; Xie, P.; Li, J.; Wang, S.; Wang, L.; Zhu, X.; Gao, P.; Jiang, Q.; et al. Influence of fermented feed additive on gut morphology, immune status, and microbiota in broilers. BMC Vet. Res. 2022, 18, 218. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.Y.; Wang, C.P.; Peng, S.; Deng, D.; Tian, Z.M.; Lu, H.J.; Yu, M.; Liu, Z.C.; Ma, X.Y. Optimization of technology and evaluation of nutritional value on fermented mulberry leaves. Chin. J. Anim. Nutr. 2022, 34, 6110–6120. [Google Scholar]
- Cui, Y.; Tian, Z.; Deng, D.; Liu, Z.; Wang, G.; Chen, W.; Ma, X. Effects of dietary citrus extract on growth performance, carcass characteristics and meat quality of pigs. J. Anim. Physiol. Anim. Nutr. 2021, 106, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Tian, Z.; Yu, M.; Liu, Z.; Rong, T.; Ma, X. Effect of guanidine acetic acid on meat quality, muscle amino acids, and fatty acids in Tibetan pigs. Front. Vet. Sci. 2022, 9, 998956. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, Z.; Chen, W.; Rong, T.; Wang, G.; Ma, X. Hermetia illucens larvae as a potential dietary protein source altered the microbiota and modulated mucosal immune status in the colon of finishing pigs. J. Anim. Sci. Biotechnol. 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Zhang, C.; Yang, Y.; Mu, C.; Su, Y.; Yu, K.; Zhu, W. Long-term effects of early antibiotic intervention on blood parameters, apparent nutrient digestibility, and fecal microbial fermentation profile in pigs with different dietary protein levels. J. Anim. Sci. Biotechnol. 2017, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Cui, X.; Yang, Y.; Zhang, M.; Jiao, F.; Gan, T.; Lin, Z.; Huang, Y.; Wang, H.; Liu, S.; Bao, L.; et al. Optimized ensiling conditions and microbial community in mulberry leaves silage with inoculants. Front. Microbiol. 2022, 13, 813363. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Wang, C.; He, L.; Zhou, W.; Yang, F.; Zhang, Q. The bacterial community and fermentation quality of mulberry (Morus alba) leaf silage with or without Lactobacillus casei and sucrose. Bioresour. Technol. 2019, 293, 122059. [Google Scholar] [CrossRef]
- Ding, Z.; Bai, J.; Xu, D.; Li, F.; Zhang, Y.; Guo, X. Microbial community dynamics and natural fermentation profiles of ensiled alpine grass elymus nutans prepared from different regions of the Qinghai-Tibetan Plateau. Front. Microbiol. 2020, 11, 855. [Google Scholar] [CrossRef]
- Mcgarvey, J.A.; Franco, R.B.; Palumbo, J.D.; Hnasko, R.; Stanker, L.; Mitloehner, F.M. Bacterial population dynamics during the ensiling of Medicago sativa (alfalfa) and subsequent exposure to air. J. Appl. Microbiol. 2013, 114, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Si, Q.; Wang, Z.; Liu, W.; Liu, M.; Ge, G.; Jia, Y.; Du, S. Influence of cellulase or Lactiplantibacillus plantarum on the ensiling performance and bacterial community in mixed silage of alfalfa and Leymus chinensis. Microorganisms 2023, 11, 426. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Bacterial dynamics of wheat silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef]
- Verni, M.; Wang, C.; Montemurro, M.; De Angelis, M.; Katina, K.; Rizzello, C.G.; Coda, R. Exploring the microbiota of Faba bean: Functional characterization of lactic acid bacteria. Front. Microbiol. 2017, 8, 2461. [Google Scholar] [CrossRef]
- Nionelli, L.; Montemurro, M.; Pontonio, E.; Verni, M.; Gobbetti, M.; Rizzello, C.G. Pro-technological and functional characterization of lactic acid bacteria to be used as starters for hemp (Cannabis sativa L.) sourdough fermentation and wheat bread fortification. Int. J. Food Microbiol. 2018, 279, 14–25. [Google Scholar] [CrossRef]
- Hopper, W.; Mahadevan, A. Utilization of catechin and its metabolites by Bradyrhizobium japonicum. Appl. Microbiol. Biot. 1991, 35, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Peng, Y.; He, J.; Xiao, D.; Chen, C.; Li, F.; Huang, R.; Yin, Y. Dietary mulberry leaf powder affects growth performance, carcass traits and meat quality in finishing pigs. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1934–1945. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Jiang, J.; Yu, J.; Mao, X.; Yu, B.; Chen, D. Effect of dietary supplementation with mulberry (Morus alba L.) leaves on the growth performance, meat quality and antioxidative capacity of finishing pigs. J. Integr. Agr. 2019, 18, 143–151. [Google Scholar] [CrossRef]
- Chen, G.; Su, Y.; Cai, Y.; He, L.; Yang, G. Comparative transcriptomic analysis reveals beneficial effect of dietary mulberry leaves on the muscle quality of finishing pigs. Vet. Med. Sci. 2019, 5, 526–535. [Google Scholar] [CrossRef]
- Chen, Z.; Xie, Y.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Dietary supplementation with Moringa oleifera and mulberry leaf affects pork quality from finishing pigs. J. Anim. Physiol. Anim. Nutr. 2021, 105, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiao, Y.; Xie, J.; Peng, Y.; Li, F.; Chen, C.; Zhang, X.; He, J.; Xiao, D.; Yin, Y. Dietary supplementation with flavonoids from mulberry leaves improves growth performance and meat quality, and alters lipid metabolism of skeletal muscle in a Chinese Hybrid Pig. Anim. Feed. Sci. Technol. 2022, 285, 115211. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Xiao, Y.; Peng, Y.; He, J.; Chen, C.; Xiao, D.; Yin, Y.; Li, F. Mulberry leaf powder regulates antioxidative capacity and lipid metabolism in finishing pigs. Anim. Nutr. 2021, 7, 421–429. [Google Scholar] [CrossRef] [PubMed]
- D’Helft, J.; Caccialanza, R.; Derbyshire, E.; Maes, M. Relevance of ω-6 GLA added to ω-3 PUFAs supplements for ADHD: A narrative review. Nutrients 2022, 14, 3273. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R. Vitamin E research: Past, now and future. Free Radic. Biol. Med. 2021, 177, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Meng, Q.; Zhou, B.; Zhou, Z. Effect of ensiled mulberry leaves and sun-dried mulberry fruit pomace on the fecal bacterial community composition in finishing steers. BMC Microbiol. 2017, 17, 97. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, Q.; Zhong, S.; Chen, Y.; Yang, Y. Comparison of rumen microbiota and serum biochemical indices in white cashmere goats fed ensiled or sun-dried mulberry leaves. Microorganisms 2020, 8, 981. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Ding, Y.; Zeng, Q.; Li, X.; Song, Z.; Fan, Z.; He, X. Effects of fermented forage mulberry powder on antioxidant performance and intestinal function of Ningxiang pigs. Chin. J. Anim. Nutr. 2019, 31, 4303–4313. [Google Scholar]
- Wang, J.; Qin, C.; He, T.; Qiu, K.; Sun, W.; Zhang, X.; Jiao, N.; Zhu, W.; Yin, J. Alfalfa-containing diets alter luminal microbiota structure and short chain fatty acid sensing in the caecal mucosa of pigs. J. Anim. Sci. Biotechnol. 2018, 9, 11. [Google Scholar] [CrossRef]
- Pu, G.; Li, P.; Du, T.; Niu, Q.; Fan, L.; Wang, H.; Liu, H.; Li, K.; Niu, P.; Wu, C.; et al. Adding appropriate fiber in diet increases diversity and metabolic capacity of distal gut microbiota without altering fiber digestibility and growth rate of finishing pig. Front. Microbiol. 2020, 11, 533. [Google Scholar] [CrossRef]
- Cui, S.; Guo, W.; Chen, C.; Tang, X.; Zhao, J.; Mao, B.; Zhang, H. Metagenomic analysis of the effects of Lactiplantibacillus plantarum and fructooligosaccharides (FOS) on the fecal microbiota structure in mice. Foods 2022, 11, 1187. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Wu, J.; Tan, Z.; Wang, S.; Yazawa, T. Different intestinal microbiota with growth stages of three-breed hybrid pig. Biomed. Res. Int. 2022, 2022, 5603451. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, K.; Xiong, K. Fecal microbial changes in response to finishing pigs directly fed with fermented feed. Front. Vet. Sci. 2022, 9, 894909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, C.; Schroyen, M.; Everaert, N.; Ma, T.; Zhang, H. Effects of the inclusion of fermented mulberry leaves and Branches in the gestational diet on the performance and gut microbiota of sows and their offspring. Microorganisms 2021, 9, 604. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Mu, A.; Haslam, N.; Schwartz, O.S.; Simmons, J.G. Feeling down? A systematic review of the gut microbiota in anxiety/depression and irritable bowel syndrome. J. Affect. Disord. 2020, 266, 429–446. [Google Scholar] [CrossRef]
- Guo, X.; Huang, C.; Xu, J.; Xu, H.; Liu, L.; Zhao, H.; Wang, J.; Huang, W.; Peng, W.; Chen, Y.; et al. Gut microbiota is a potential biomarker in inflammatory bowel disease. Front. Nutr. 2022, 8, 818902. [Google Scholar] [CrossRef]
- Minamoto, Y.; Minamoto, T.; Isaiah, A.; Sattasathuchana, P.; Buono, A.; Rangachari, V.R.; McNeely, I.H.; Lidbury, J.; Steiner, J.M.; Suchodolski, J.S. Fecal short-chain fatty acid concentrations and dysbiosis in dogs with chronic enteropathy. J. Vet. Int. Med. 2019, 33, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Qin, W.; Xu, Y.; Yang, W.; Chen, Y.; Huang, J.; Zhao, J.; Ma, L. Dietary quercetin supplementation attenuates diarrhea and intestinal damage by regulating gut microbiota in weanling piglets. Oxid. Med. Cell. Longev. 2021, 2021, 6221012. [Google Scholar] [CrossRef] [PubMed]
- Khosroshahi, H.T.; Abedi, B.; Ghojazadeh, M.; Samadi, A.; Jouyban, A. Effects of fermentable high fiber diet supplementation on gut derived and conventional nitrogenous product in patients on maintenance hemodialysis: A randomized controlled trial. Nutr. Metab. 2019, 16, 18. [Google Scholar] [CrossRef]
- Wang, Z.; Bai, Y.; Pi, Y.; Gerrits, W.J.J.; de Vries, S.; Shang, L.; Tao, S.; Zhang, S.; Han, D.; Zhu, Z.; et al. Xylan alleviates dietary fiber deprivation-induced dysbiosis by selectively promoting Bifidobacterium pseudocatenulatum in pigs. Microbiome 2021, 9, 227. [Google Scholar] [CrossRef]
- Gong, L.; Cao, W.; Chi, H.; Wang, J.; Zhang, H.; Liu, J.; Sun, B. Whole cereal grains and potential health effects: Involvement of the gut microbiota. Food Res. Int. 2018, 103, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus subtilis DSM 32315 manipulates intestinal structure and microbial composition in broiler chickens. Sci. Rep. 2018, 8, 15358. [Google Scholar] [CrossRef] [PubMed]
- Li, I.; Wu, S.; Liou, J.; Liu, H.; Chen, J.; Chen, C. Effects of Deinococcus spp. supplement on egg quality traits in laying hens. Poult. Sci. 2018, 97, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Bottacini, F.; van Sinderen, D.; Ventura, M. Omics of bifidobacteria: Research and insights into their health-promoting activities. Biochem. J. 2017, 474, 4137–4152. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lu, M.; Wang, J.; Zhang, H.; Qiu, K.; Qi, G.; Wu, S. Dietary oregano essential oil supplementation improves intestinal functions and alters gut microbiota in late-phase laying hens. J. Anim. Sci. Biotechnol. 2021, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Tao, L.; Liu, H.; Yang, G. Effects of fermented feed on growth performance, immune organ indices, serum biochemical parameters, cecal odorous compound production, and the microbiota community in broilers. Poult. Sci. 2023, 102, 102629. [Google Scholar] [CrossRef]
- Recharla, N.; Kim, K.; Park, J.; Jeong, J.; Jeong, Y.; Lee, H.; Hwang, O.; Ryu, J.; Baek, Y.; Oh, Y.; et al. Effects of amino acid composition in pig diet on odorous compounds and microbial characteristics of swine excreta. J. Anim. Sci. Technol. 2017, 59, 28. [Google Scholar] [CrossRef]
- Jha, R.; Berrocoso, J.F.D. Dietary fiber and protein fermentation in the intestine of swine and their interactive effects on gut health and on the environment: A review. Anim. Feed Sci. Technol. 2016, 212, 18–26. [Google Scholar] [CrossRef]
- Kerr, B.J.; Trabue, S.L.; van Weelden, M.B.; Andersen, D.S.; Pepple, L.M. Impact of narasin on manure composition, microbial ecology, and gas emissions from finishing pigs fed either a corn-soybean meal or a corn-soybean meal-dried distillers grains with solubles diets. J. Anim. Sci. 2018, 96, 1317–1329. [Google Scholar] [CrossRef]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef]
- Li, X.; Jensen, B.B.; Canibe, N. The mode of action of chicory roots on skatole production in entire male pigs is neither via reducing the population of skatole-producing bacteria nor via increased butyrate production in the hindgut. Appl. Environ. Microbiol. 2019, 85, e02327-18. [Google Scholar] [CrossRef] [PubMed]
- Vhile, S.G.; Kjos, N.P.; Sørum, H.; Øverland, M. Feeding Jerusalem artichoke reduced skatole level and changed intestinal microbiota in the gut of entire male pigs. Animal 2012, 6, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Pi, Y.; Gao, K.; Peng, Y.; Mu, C.L.; Zhu, W.Y. Antibiotic-induced alterations of the gut microbiota and microbial fermentation in protein parallel the changes in host nitrogen metabolism of growing pigs. Animal 2019, 13, 262–272. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | CON | FML | p-Value |
|---|---|---|---|
| Carcass performance | |||
| Slaughter weight (kg) | 132.73 ± 1.63 | 128.23 ± 13.35 | 0.431 |
| Carcass weight (kg) | 96.84 ± 3.14 | 90.82 ± 9.01 | 0.153 |
| Carcass yield (%) | 72.97 ± 2.63 | 70.86 ± 1.79 | 0.136 |
| Abdominal fat weight (kg) | 2.63 ± 1.05 | 1.71 ± 0.55 | 0.088 |
| Back fat thickness (mm) | 29.91 ± 6.31 | 27.56 ± 6.22 | 0.531 |
| Loin muscle area (cm2) | 60.93 ± 2.03 | 63.03 ± 1.70 | 0.646 |
| Meat quality | |||
| L* 45 min | 48.06 ± 1.84 | 48.28 ± 1.07 | 0.805 |
| L* 24 h | 58.56 ± 3.99 | 59.79 ± 1.38 | 0.503 |
| a* 45 min | 17.28 ± 0.85 | 17.30 ± 0.85 | 0.956 |
| a* 24 h | 17.01 ± 1.27 | 17.18 ± 1.53 | 0.833 |
| b* 45 min | 5.87 ± 0.41 | 6.22 ± 0.45 | 0.191 |
| b* 24 h | 7.48 ± 0.76 | 8.13 ± 1.00 | 0.232 |
| pH 45 min | 6.22 ± 0.17 | 6.16 ± 0.11 | 0.502 |
| pH 24 h | 5.56 ± 0.18 | 5.52 ± 0.11 | 0.677 |
| Drip loss 24 h (%) | 2.29 ± 0.12 | 2.21 ± 0.20 | 0.434 |
| Shear force (N) | 64.76 ± 14.23 | 60.23 ± 6.02 | 0.489 |
| Marbling score | 2.70 ± 0.32 | 2.54 ± 0.73 | 0.639 |
| Items | CON | FML | p-Value |
|---|---|---|---|
| Weight gain (kg) | 49.20 ± 2.71 | 49.36 ± 3.14 | 0.970 |
| Feed intake (kg) | 173.97 ± 5.72 | 178.88 ± 5.54 | 0.556 |
| Feed cost (CNY/pig) | 644.69 ± 23.59 | 597.44 ± 18.50 | 0.155 |
| Income from weight gain (CNY/pig) | 1176.86 ± 64.94 | 1180.62 ± 75.00 | 0.970 |
| Gross profit (CNY/pig) | 532.17 ± 43.39 | 583.18 ± 58.51 | 0.485 |
| Feed cost of weight gain (CNY/kg) | 13.22 ± 0.30 | 12.26 ± 0.42 | 0.078 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Y.; Liu, Z.; Deng, D.; Tian, Z.; Song, M.; Lu, Y.; Yu, M.; Ma, X. Influence of Fermented Mulberry Leaves as an Alternative Animal Feed Source on Product Performance and Gut Microbiome in Pigs. Fermentation 2024, 10, 215. https://doi.org/10.3390/fermentation10040215
Cui Y, Liu Z, Deng D, Tian Z, Song M, Lu Y, Yu M, Ma X. Influence of Fermented Mulberry Leaves as an Alternative Animal Feed Source on Product Performance and Gut Microbiome in Pigs. Fermentation. 2024; 10(4):215. https://doi.org/10.3390/fermentation10040215
Chicago/Turabian StyleCui, Yiyan, Zhichang Liu, Dun Deng, Zhimei Tian, Min Song, Yusheng Lu, Miao Yu, and Xianyong Ma. 2024. "Influence of Fermented Mulberry Leaves as an Alternative Animal Feed Source on Product Performance and Gut Microbiome in Pigs" Fermentation 10, no. 4: 215. https://doi.org/10.3390/fermentation10040215
APA StyleCui, Y., Liu, Z., Deng, D., Tian, Z., Song, M., Lu, Y., Yu, M., & Ma, X. (2024). Influence of Fermented Mulberry Leaves as an Alternative Animal Feed Source on Product Performance and Gut Microbiome in Pigs. Fermentation, 10(4), 215. https://doi.org/10.3390/fermentation10040215