Debaryomyces hansenii Strains from Traditional Chinese Dry-Cured Ham as Good Aroma Enhancers in Fermented Sausage

Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of a Yeast Strain Starter
2.2. Preparation of Dry-Cured Fermented Sausages
2.3. Chemical Analysis
2.4. Texture Parameters
2.5. Free Fatty Acids (FFAs)
2.6. Free Amino Acid
2.7. Volatile Compounds
2.8. Sensory Evaluation
2.9. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Analysis
3.2. Texture
3.3. Free Amino Acids
3.4. Free Fatty Acids
3.5. Flavor
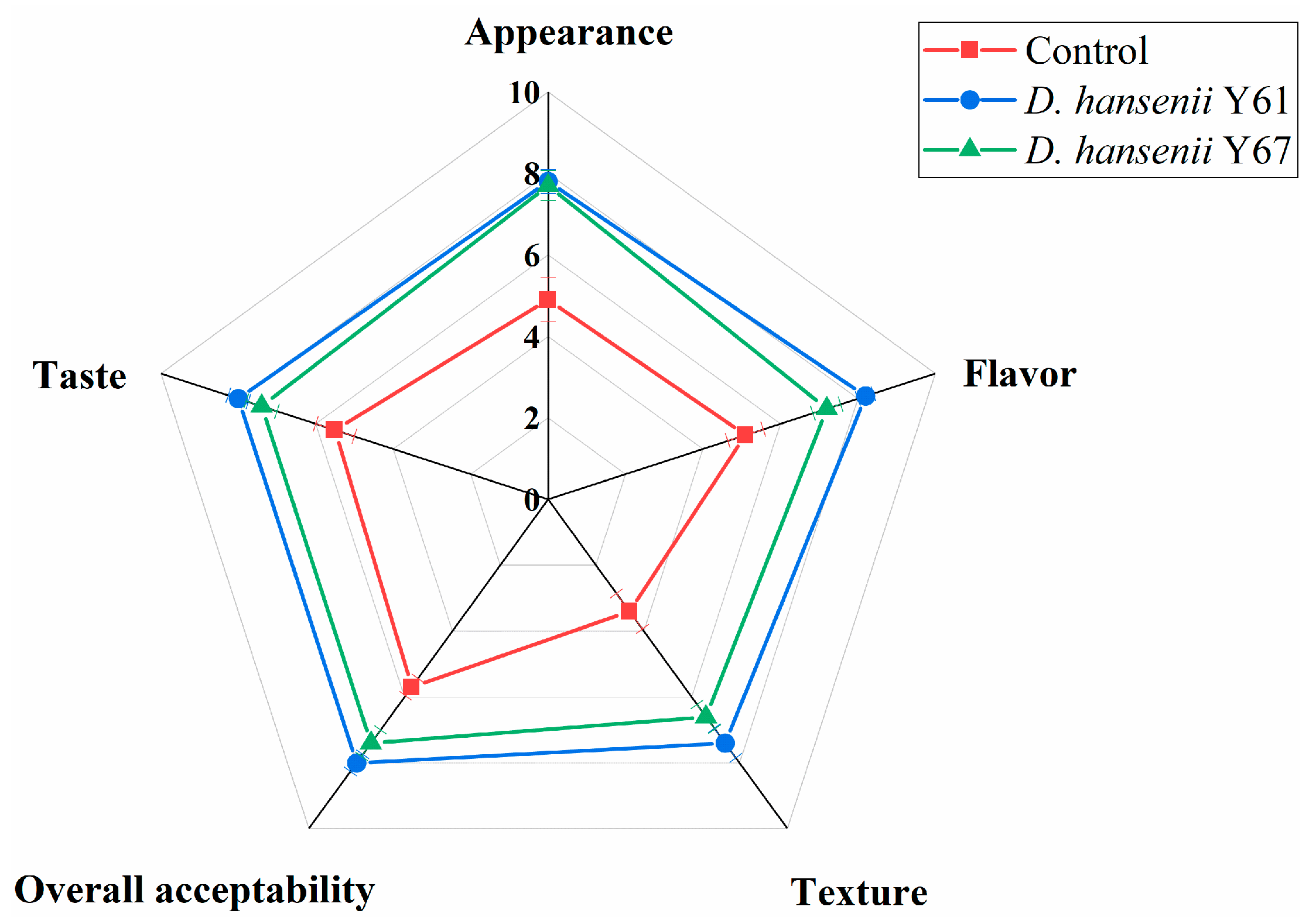
3.6. Sensory Evaluation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arihara, K. Probiotics. In Handbook of Fermented Meat and Poultry; Toldrá, F., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 155–160. [Google Scholar]
- Perea-Sanz, L.; López-Díez, J.J.; Belloch, C.; Flores, M. Counteracting the effect of reducing nitrate/nitrite levels on dry fermented sausage aroma by Debaryomyces hansenii inoculation. Meat Sci. 2020, 164, 108103. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Moreno, L.; Ruiz-Pérez, F.; Rodríguez-Castro, E.; Ramos, J. Debaryomyces hansenii is a real tool to improve a diversity of characteristics in sausages and dry-meat products. Microorganisms 2021, 9, 1512. [Google Scholar] [CrossRef] [PubMed]
- Núñez, F.; Rodríguez, M.M.; Córdoba, J.J.; Bermúdez, M.E.; Asensio, M.A. Yeast population during ripening of dry-cured Iberian ham. Int. J. Food Microbiol. 1996, 29, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Cano-García, L.; Belloch, C.; Flores, M. Impact of Debaryomyces hansenii strains inoculation on the quality of slow dry-cured fermented sausages. Meat Sci. 2014, 96, 1469–1477. [Google Scholar] [CrossRef]
- Flores, M.; Moncunill, D.; Montero, R.; López-Díez, J.J.; Belloch, C. Screening of Debaryomyces hansenii strains for flavor production under a reduced concentration of nitrifying preservatives used in meat products. J. Agric. Food Chem. 2017, 65, 3900–3909. [Google Scholar] [CrossRef]
- Ramos-Moreno, L.; Ruiz-Castilla, F.J.; Bravo, C.; Martínez, E. Inoculation with a terroir selected Debaryomyces hansenii strain changes physico-chemical characteristics of Iberian cured pork loin. Meat Sci. 2019, 157, 107875. [Google Scholar] [CrossRef]
- Martín, A.; Córdoba, J.; Benito, M.; Aranda, E.; Asensio, M. Effect of Penicillium chrysogenum and Debaryomyces hansenii on the volatile compounds during controlled ripening of pork loins. Int. J. Food Microbiol. 2003, 84, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Elias, M.; Fraqueza, M.J. The Use of Starter Cultures in Traditional Meat Products. J. Food Quality 2017, 2017, 9546026. [Google Scholar] [CrossRef]
- Meftah, S.; Abid, S.; Dias, T.; Rodrigues, P. Effect of dry-sausage starter culture and endogenous yeasts on Aspergillus westerdijkiae and Penicillium nordicum growth and OTA production. LWT-Food Sci. Technol. 2018, 87, 250–258. [Google Scholar] [CrossRef]
- Ramos, J.; Melero, Y.; Ramos-Moreno, L.; Michán, C.; Cabezas, L. Debaryomyces hansenii strains from Valle de los pedroches Iberian dry meat products: Solation, identification, characterization, and selection for starter cultures. J. Microbiol. Biotechnol. 2017, 27, 1576–1585. [Google Scholar] [CrossRef]
- Xue, F.; Jiang, Y.S.; Zhang, W.J.; Zhao, T.T. Effect of microbial culture starters on free amino acid in rabbit sausage. Adv. Mater. Res. 2013, 781–784, 1801–1805. [Google Scholar] [CrossRef]
- Andrade, M.J.; Córdoba, J.J.; Casado, E.M.; Córdoba, M.G.; Rodríguez, M. Effect of selected strains of Debaryomyces hansenii on the volatile compound production of dry fermented sausage “salchichón”. Meat Sci. 2010, 85, 256–264. [Google Scholar] [CrossRef] [PubMed]
- García-Béjar, B.; Sánchez-Carabias, D.; Arévalo-Villena, M.; Briones, A. Autochthonous yeast from pork and game meat fermented sausages for application in meat protection and aroma developing. Animals 2020, 10, 2340. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Corral, S.; Cano-García, L.; Salvador, A.; Belloch, C. Yeast strains as potential aroma enhancers in dry fermented sausages. Int. J. Food Sci. Technol. 2015, 212, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Selgas, M.D.; García, M.L. Yeasts. In Handbook of Fermented Meat and Poultry; Toldrá, F., Ed.; Wiley Blackwell: Hoboken, NJ, USA, 2015; pp. 139–146. [Google Scholar]
- Belloch, C.; Perea-Sanz, L.; Gamero, A.; Flores, M. Selection of Debaryomyces hansenii isolates as starters in meat products based on phenotypic virulence factors, tolerance to abiotic stress conditions, and aroma generation. J. Appl. Microbiol. 2022, 133, 200–211. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 4: Suitability of taxonomic units notified to EFSA until March 2016. EFSA J. 2016, 14, e04522. [Google Scholar] [CrossRef]
- Asefa, D.T. Yeast diversity and dynamics in the production processes of Norwegian dry-cured meat products. Int. J. Food Microbiol. 2009, 133, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Cocolin, L.; Urso, R.; Rantsiou, K.; Cantoni, C.; Comi, G. Dynamics and characterization of yeasts during natural fermentation of Italian sausages. FEMS Yeast Res. 2006, 6, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Ochangco, H.S.; Gamero, A.; Smith, I.M.; Christensen, J.E.; Jespersen, L.; Arneborg, N. In vitro investigation of Debaryomyces hansenii strains for potential probiotic properties. World J. Microbiol. Biotechnol. 2016, 32, 141. [Google Scholar] [CrossRef]
- Selgas, M.D.; Ros, J.; García, M.L. Effect of selected yeast strains on the sensory properties of dry fermented sausages. Eur. Food Res. Technol. 2003, 217, 475–480. [Google Scholar] [CrossRef]
- Hu, Y.Y.; Li, Y.; Zhu, J.; Kong, B.; Liu, Q.; Chen, Q. Improving the taste profile of reduced-salt dry sausage by inoculating different lactic acid bacteria. Food Res. Int. 2021, 145, 110391. [Google Scholar] [CrossRef]
- Corral, S.; Belloch, C.; López-Díez, J.J.; Salvador, A.; Flores, M. Yeast inoculation as a strategy to improve the physico-chemical and sensory properties of reduced salt fermented sausages produced with entire male fat. Meat Sci. 2017, 123, 1–7. [Google Scholar] [CrossRef]
- Durá, M.A.; Flores, M.; Toldrá, F. Effect of growth phase and dry-cured sausage processing conditions on Debaryomyces spp. generation of volatile compounds from branched-chain amino acids. Food Chem. 2004, 86, 391–399. [Google Scholar] [CrossRef]
- Rhee, K.S.; Anderson, L.M.; Sams, A.R. Lipid oxidation potential of beef, chicken and pork. J. Food Sci. 1996, 61, 8–12. [Google Scholar] [CrossRef]
- Engberg, R.M.; Lauridsen, C.; Jensen, S.K.; Jakobsen, K. Inclusion of oxidized vegetable oil in broiler diets. Its influence on nutrient balance and on the antioxidative status of broilers. Poultry Sci. 1996, 75, 1003–1011. [Google Scholar] [CrossRef]
- Chen, L.S.; Ma, Y.; Chen, L.J.; Zhao, C.H.; Maubois, J.L.; Jiang, T.M.; Li, H.M.; He, S.H. Antioxidant activity of two yeasts and their attenuation effect on 4-nitroquinoline 1-oxide induced in vitrolipid peroxidation. Int. J. Food Sci. Technol. 2010, 45, 555–561. [Google Scholar] [CrossRef]
- Mancini, R.A.; Hunt, M.C. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef]
- Faustmana, C.; Specht, S.M.; Malkusa, L.A.; Kinsmana, D.M. Pigment oxidation in ground veal: Influence of lipid oxidation, iron and zinc. Meat Sci. 1992, 31, 351–362. [Google Scholar] [CrossRef]
- Bruce, H.L.; Aalhus, J.L. Advances in the understanding and measurement of meat texture. In New Aspects of Meat Quality from Genes to Ethics; Peter, P.P., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 129–166. [Google Scholar]
- Dashdorj, D.; Amna, T.; Hwang, I. Influence of specific taste-active components on meat flavor as affected by intrinsic and extrinsic factors: An overview. Eur. Food Res. Technol. 2015, 241, 157–171. [Google Scholar] [CrossRef]
- Ramalingam, V.; Song, Z.; Hwang, I. The potential role of secondary metabolites in modulating the flavor and taste of the meat. Food Res Int. 2019, 122, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Bajpai, V.K.; Kim, H.R.; Kang, S.C. Antibacterial activity of bioconverted eicosapentaenoic (EPA) and docosahexaenoic acid (DHA) against foodborne pathogenic bacteria. Int. J. Food Microbiol. 2007, 113, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Cano-García, L.; Rivera-Jiménez, S.; Belloch, C.; Flores, M. Generation of aroma compounds in a fermented sausage meat model system by Debaryomyces hansenii strains. Food Chem. 2014, 151, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Calkins, C.R.; Hodgen, J.M. A fresh look at meat flavor. Meat Sci. 2007, 77, 63–80. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Parameters | Group | ||
|---|---|---|---|
| Control | D. hansenii Y61 | D. hansenii Y67 | |
| pH | 5.96 ± 0.05 a | 4.76 ± 0.08 b | 4.92 ± 0.01 b |
| aw | 0.81 ± 0.002 a | 0.76 ± 0.001 b | 0.74 ± 0.001 b |
| TBARS (mg/kg) | 0.88 ± 0.031 a | 0.52 ± 0.020 c | 0.55 ± 0.052 b |
| Brightness (L*) | 35.44 ± 0.13 b | 39.78 ± 0.62 b | 41.14 ± 0.01 a |
| Redness (a*) | 5.38 ± 0.04 b | 13.88 ± 0.9 a | 13.01 ± 0.01 a |
| Yellowness (b*) | 12.25 ± 1.14 c | 8.63 ± 0.06 a | 9.23 ± 0.02 b |
| Yeasts (cfu/g) | 9.1 × 102 c | 3.79 × 104 b | 4.27 × 104 a |
| Total bacteria (cfu/g) | 6.24 × 104 a | 4.71 × 104 c | 5.92 × 104 b |
| Enterobacteriaceae (cfu/g) | 2.00 c | 1.00 b | 0 a |
| Parameters | Group | ||
|---|---|---|---|
| Control | D. hansenii Y61 | D. hansenii Y67 | |
| Hardness/(N·cm−2) | 1493.0 ± 21.22 c | 7577.7 ± 37.56 a | 4245.0 ± 31.58 b |
| Springiness/cm | 0.76 ± 0.01 a | 0.70 ± 0.05 a | 0.71 ± 0.01 a |
| Cohesiveness | 0.59 ± 0.02 c | 0.72 ± 0.05 a | 0.67 ± 0.02 b |
| Gumminess/(N·S) | 1080.6 ± 37.66 c | 4684.21 ± 380.77 a | 2557.26 ± 105.32 b |
| Chewiness/(N·cm−1) | 825.10 ± 24.38 c | 3313.25 ± 489.24 a | 1805.78 ± 108.73 b |
| FAA | Group | ||
|---|---|---|---|
| Control | D. hansenii Y61 | D. hansenii Y67 | |
| Asp | 64.1 ± 0.15 b | 60.38 ± 0.01 c | 79.79 ± 0.02 a |
| Glu | 487.07 ± 0.43 c | 662.1 ± 0.50 b | 714.0 ± 0.37 a |
| Cys | 4.4 ± 0.02 b | 13.33 ± 0.05 a | nd c |
| Ser | 160.95 ± 0.01 c | 193.46 ± 0.05 b | 199.86 ± 0.03 a |
| Gly | 220.07 ± 0.33 b | 268.86 ± 0.37 a | 265.35 ± 0.40 a |
| His | 533.29 ± 0.15 b | 268.86 ± 0.26 c | 695.3 ± 0.30 a |
| Arg | 3948.71 ± 0.42 b | 4388.21 ± 0.56 a | 4418.5 ± 0.31 a |
| Thr * | 131.78 ± 0.25 c | 151.85 ± 0.23 b | 155.66 ± 0.21 a |
| Ala | 502.6 ± 0.05 c | 624.61 ± 0.02 a | 606.01 ± 0.05 b |
| Pro | 551.64 ± 0.04 c | 600.28 ± 0.01 b | 621.19 ± 0.05 a |
| Tyr | 79.0 ± 0.02 b | 111.95 ± 0.01 a | 113.4 ± 0.02 a |
| Val * | 169.59 ± 0.15 b | 222.74 ± 0.21 a | 219.66 ± 0.11 a |
| Met | 128.48 ± 0.39 a | 81.54 ± 0.22 b | 83.2 ± 0.35 b |
| Lle * | 128.48 ± 0.23 b | 165.69 ± 0.17 a | 163.81 ± 0.21 a |
| Leu * | 202.91 ± 0.11 b | 251.57 ± 0.15 a | 249.21 ± 0.24 a |
| Phe * | 103.47 ± 0.10 b | 126.19 ± 0.03 a | 123.81 ± 0.2 a |
| Lys * | 429.63 ± 0.20 c | 593.44 ± 0.20 a | 535.42 ± 0.26 b |
| ∑FAA | 7846.17 ± 3.05 c | 8785.06 ± 3.14 b | 9244.18 ± 3.13 a |
| ∑EAA | 1165.86 ± 1.04 c | 1511.48 ± 0.99 a | 1447.57 ± 1.23 b |
| FFA | Group | ||
|---|---|---|---|
| Control | D. hansenii Y61 | D. hansenii Y67 | |
| C10:0 | 34.30 ± 0.15 a | 5.92 ± 0.24 b | 0 c |
| C12:0 | 27.41 ± 0.45 b | 65.07 ± 0.48 a | 13.85 ± 2.70 c |
| C14:0 | 204.57 ± 1.80 a | 269.98 ± 1.68 b | 90.68 ± 10.03 c |
| C16:0 | 274.85 ± 0.15 a | 230.228 ± 3.12 b | 197.65 ± 2.43 c |
| C18:0 | 68.14 ± 0.45 a | 23.668 ± 0.48 c | 36.92 ± 2.70 b |
| C20:0 | 17.92 ± 4.80 c | 103.64 ± 0.24 b | 214.76 ± 5.40 a |
| ∑SFA | 627.19 ± 7.80 a | 698.48 ± 8.40 b | 553.86 ± 18.90 c |
| C14:1 n-5 | 18.68 ± 0.30 a | 15.85 ± 1.20 c | 19.28 ± 0.81 b |
| C16:1 n-3 | 45.63 ± 1.80 a | 27.21 ± 28.08 b | 30.14 ± 2.97 c |
| C18:1 n-9 | 497.80 ± 19.05 c | 802.11 ± 36.24 b | 1184.81 ± 40.50 a |
| C20:1 n-9 | 94.63 ± 11.55 c | 170.12 ± 32.16 b | 212.58 ± 4.32 a |
| ∑MUFA | 656.74 ± 32.7 b | 1015.31 ± 97.68 b | 1446.81 ± 48.60 a |
| C18:2 n-6 | 91.41 ± 0.75 c | 143.86 ± 28.32 b | 176.74 ± 35.37 a |
| C20:2 n-6 | 33.23 ± 0.30 c | 75.95 ± 5.04 b | 109.96 ± 3.24 a |
| C20:5 n-3 (EPA) | 72.12 ± 0.15 c | 123.75 ± 2.88 b | 165.88 ± 13.50 a |
| C22:2 n-6 | 43.33 ± 0.15 b | 85.42 ± 1.20 a | 52.94 ± 0.54 c |
| C22:6 n-3 (DHA) | 22.51 ± 0.45 c | 170.12 ± 0.24 b | 196.02 ± 2.70 a |
| ∑PUFA | 247.29 ± 1.80 c | 652.34 ± 40.08 a | 714.30 ± 55.89 b |
| Total | 1531.22 ± 84.60 c | 2366.13 ± 285.36 b | 2714.97 ± 250.60 a |
| Number | Volatile Compound | Group | ||
|---|---|---|---|---|
| Control | D. hansenii Y61 | D. hansenii Y67 | ||
| Aldehydes | ||||
| 1 | Hexanal | 11.4 ± 0.01 a | 2.65 ± 0.12 b | 1.17 ± 0.02 c |
| 2 | 2-Heptenal | nd b | nd b | 1.11 ± 0.15 a |
| 3 | 2-Octenal | nd b | nd b | 1.15 ± 0.02 a |
| Total | 11.4 ± 0.01 a | 2.65 ± 0.12 c | 3.43 ± 0.19 b | |
| Ketones | ||||
| 4 | 2,2-dimethyl-Cyclobutanone | nd b | nd b | 1.1 ± 0.01 a |
| 5 | 3,4-Hexanedione | 2.12 ± 0.03 a | 0.29 ± 0.05 b | nd c |
| 6 | 3,5-Octadien-2-one | 1.22 ± 0.01 b | 1.14 ± 0.11 a | 0.47 ± 0.05 c |
| 7 | Pyrimidine-2,4-dione | nd b | nd b | 0.38 ± 0.01 a |
| 8 | 5,8-Quinolinedione | nd b | 0.44 ± 0.01 a | nd b |
| Total | 3.78 ± 0.04 ab | 2.16 ± 0.17 a | 1.95 ± 0.07 c | |
| Alcohols | ||||
| 9 | 5-methyl-2-Heptanol | 8.11 ± 0.10 a | nd b | nd b |
| 10 | 1,6-Octadien-3-ol | 1.73 ± 0.12 a | 0.6 ± 0.01 c | 0.93 ± 0.01 b |
| 11 | Ethanol | 4.07 ± 0.02 a | 2.1 ± 0.01 c | 2.46 ± 0.12 b |
| 12 | 1-Nonanol | nd b | 0.99 ± 0.03 a | nd b |
| Total | 13.91 ± 0.24 a | 3.69 ± 0.05 b | 3.39 ± 0.13 c | |
| Acids | ||||
| 13 | Formic acid | 26.78 ± 0.15 a | nd b | nd b |
| 14 | 2-methyl-Butanoic acid | 0.78 ± 0.01 a | nd b | nd b |
| 15 | Acetic acid | 1.1 ± 0.01 c | 21.61 ± 0.41 b | 24.67 ± 0.32 a |
| 16 | Benzoic acid | 0.45 ± 0.01 a | nd b | nd b |
| 17 | Propanoic acid | 0.67 ± 0.05 c | 4.25 ± 0.01 a | 3.27 ± 0.01 b |
| 18 | Propanedioic acid | nd c | 14.09 ± 0.1 a | 8.56 ± 0.10 b |
| 19 | 2-ethyl-Heptanoic acid | nd b | 0.37 ± 0.01 a | nd b |
| 20 | Lactic acid | 0.79 ± 0.30 c | 2.76 ± 0.12 b | 3.44 ± 0.10 a |
| 21 | Pentanoic acid | nd b | nd b | 0.31 ± 0.10 a |
| 22 | Mercaptoacetic acid | nd c | 0.45 ± 0.01 a | 0.36 ± 0.05 b |
| 23 | 2-Cyclopentene-1-carboxylic acid | nd b | nd b | 0.11 ± 0.03 a |
| 24 | 1,2-Cinnolinedicarboxylic acid | nd b | nd b | 0.7 ± 0.12 a |
| 25 | trans-Cinnamic acid | nd b | nd b | 0.48 ± 0.01 a |
| 26 | Methylphosphonic acid | nd b | nd b | 0.21 ± 0.07 a |
| 27 | 5-Aminovaleric acid | nd b | 0.76 ± 0.03 a | nd b |
| Total | 30.57 ± 0.23 c | 44.29 ± 0.78 a | 42.21 ± 0.91 b | |
| Ester compounds | ||||
| 28 | Hexanoic acid, ethyl ester | 15.42 ± 0.12 a | 18.19 ± 0.13 b | 11.53 ± 0.07 c |
| 29 | Octanoic acid, ethyl ester | nd c | 1.69 ± 0.04 a | 0.82 ± 0.06 b |
| 30 | Acetic acid, butyl ester | nd c | 0.58 ± 0.12 b | 1.41 ± 0.07 a |
| 31 | Decanoic acid, ethyl ester | nd b | nd b | 3.84 ± 0.30 a |
| 32 | Isobutyl ester | nd b | 0.4 ± 0.12 a | nd b |
| 33 | Butyrolactone | nd c | 3.22 ± 0.05 b | 1.96 ± 0.10 a |
| 34 | Ethyl 9-hexadecenoate | nd b | 0.88 ± 0.11 a | nd b |
| 35 | Isoamyl lactate | nd b | nd b | 0.18 ± 0.01 a |
| 36 | Hexadecanoic acid, ethyl ester | nd b | 0.24 ± 0.01 a | nd b |
| 37 | Methoxyacetic acid, butyl ester | nd c | 7.22 ± 0.03 b | 13.82 ± 0.12 a |
| 38 | Hexanoic acid, methyl ester | nd b | nd b | 2.63 ± 0.21 a |
| 39 | Octanoic acid, ethyl ester | nd c | 0.64 ± 0.03 b | 0.82 ± 0.01 a |
| 40 | Heptanoic acid, ethyl ester | nd b | nd b | 1.67 ± 0.12 a |
| 41 | Pentanoic acid, ethyl ester | 0.86 ± 0.01 b | 0.78 ± 0.21 c | 1.77 ± 0.05 a |
| 42 | Trimethylsilyl ester | 1.4 ± 0.03 a | 1.4 ± 0.01 a | 0.74 ± 0.02 b |
| 43 | Formic acid, hept-2-yl ester | nd b | 0.4 ± 0.06 a | ndb |
| 44 | 3,4-Hexanedione, 2,2,5-trimethyl | 4.12 ± 0.02 b | 4.35 ± 0.10 a | nd c |
| 45 | Pentafluoropropionate | nd b | 0.6 ± 0.01 a | nd b |
| 46 | 4-Methylcyclohexanol acetate | nd b | 0.36 ± 0.05 a | nd b |
| 47 | Fumaric acid, nonylmtetrahydrofurfuryl ester | 0.67 ± 0.03 b | 2.12 ± 0.03 a | nd c |
| Total | 22.47 ± 0.21 c | 42.05 ± 1.08 b | 41.19 ± 1.14 a | |
| Alkanes | ||||
| 48 | 1,4-dichloro-Cyclohexane | nd b | nd b | 0.18 ± 0.03 a |
| 49 | Cyclopentasiloxane | 3.4 ± 0.02 a | nd c | 1.68 ± 0.01 b |
| 50 | Dodecamethyl-pentasiloxane | 0.98 ± 0.13 a | 0.79 ± 0.05 b | 0.58 ± 0.10 c |
| 51 | Heptasiloxane | 3.76 ± 0.12 a | nd b | nd b |
| 52 | Difluorodimethyl-silane | 0.1 ± 0.03 b | nd c | 0.22 ± 0.01 a |
| 53 | 3-Methyl-oxiran | nd b | nd b | 1.36 ± 0.17 a |
| Total | 8.24 ± 0.30 a | 0.79 ± 0.05 c | 4.02 ± 0.37 b | |
| Sulphur compounds | ||||
| 54 | 3-Amino-2-phenazinol ditms | nd b | 0.48 ± 0.12 a | 0.23 ± 0.01 b |
| 55 | 2-Furanylmethyl | nd b | nd b | 1.05 ± 0.05 a |
| 56 | Tetrahydrofuran | nd b | nd b | 0.41 ± 0.10 a |
| 57 | 2-Methyl-tetrahydroquinoxaline | 0.35 ± 0.20 b | 0.14 ± 0.17 c | 0.68 ± 0.03 a |
| 58 | 2-pheny-1-Pyrroline | nd b | nd b | 0.42 ± 0.06 a |
| 59 | 4-phenyl-Quinazolin | 1.96 ± 0.05 a | nd b | ndb |
| 60 | Dimethyl ether | 7.21 ± 0.01 a | 2.75 ± 0.03 b | 0.75 ± 0.15 c |
| 61 | N-Morpholinomethyl-isopropyl-sulfide | 0.56 ± 0.05 a | nd c | 0.27 ± 0.10 b |
| Total | 10.08 ± 0.31 a | 3.37 ± 0.32 c | 6.60 ± 0.50 b | |
| Nitrogen compounds | ||||
| 62 | N-dimethylpropanamide | nd b | nd b | 0.38 ± 0.01 a |
| 63 | Formamide | 5.43 ± 0.20 a | nd b | nd b |
| 64 | L-Alanine-4-nitroanilide | nd b | nd b | 0.42 ± 0.24 a |
| Total | 5.43 ± 0.20 a | ndc | 0.80 ± 0.25 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Xiao, S.; Wang, J. Debaryomyces hansenii Strains from Traditional Chinese Dry-Cured Ham as Good Aroma Enhancers in Fermented Sausage. Fermentation 2024, 10, 152. https://doi.org/10.3390/fermentation10030152
Yang X, Xiao S, Wang J. Debaryomyces hansenii Strains from Traditional Chinese Dry-Cured Ham as Good Aroma Enhancers in Fermented Sausage. Fermentation. 2024; 10(3):152. https://doi.org/10.3390/fermentation10030152
Chicago/Turabian StyleYang, Xiaoying, Shan Xiao, and Jihui Wang. 2024. "Debaryomyces hansenii Strains from Traditional Chinese Dry-Cured Ham as Good Aroma Enhancers in Fermented Sausage" Fermentation 10, no. 3: 152. https://doi.org/10.3390/fermentation10030152
APA StyleYang, X., Xiao, S., & Wang, J. (2024). Debaryomyces hansenii Strains from Traditional Chinese Dry-Cured Ham as Good Aroma Enhancers in Fermented Sausage. Fermentation, 10(3), 152. https://doi.org/10.3390/fermentation10030152