
Oat Okara Fermentation: New Insights into the Microbiological and Metabolomic Characterization


 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Oat Okara Flour and Oat Okara Sourdough Preparation
2.2. Microbiological Analysis and pH Measurements
2.3. Molecular Characterization of Lactic Acid Bacteria at Strain Level
2.4. Genotypic Identifications of Lactic Acid Bacteria Isolates
2.5. Genotypic Identifications of Yeasts Isolates
2.6. Extraction of Metabolites and Untargeted Metabolomics Characterization
2.7. Statistical Analysis
3. Results and Discussion
3.1. Microbiological Evaluation of Oat Okara and pH Measurements
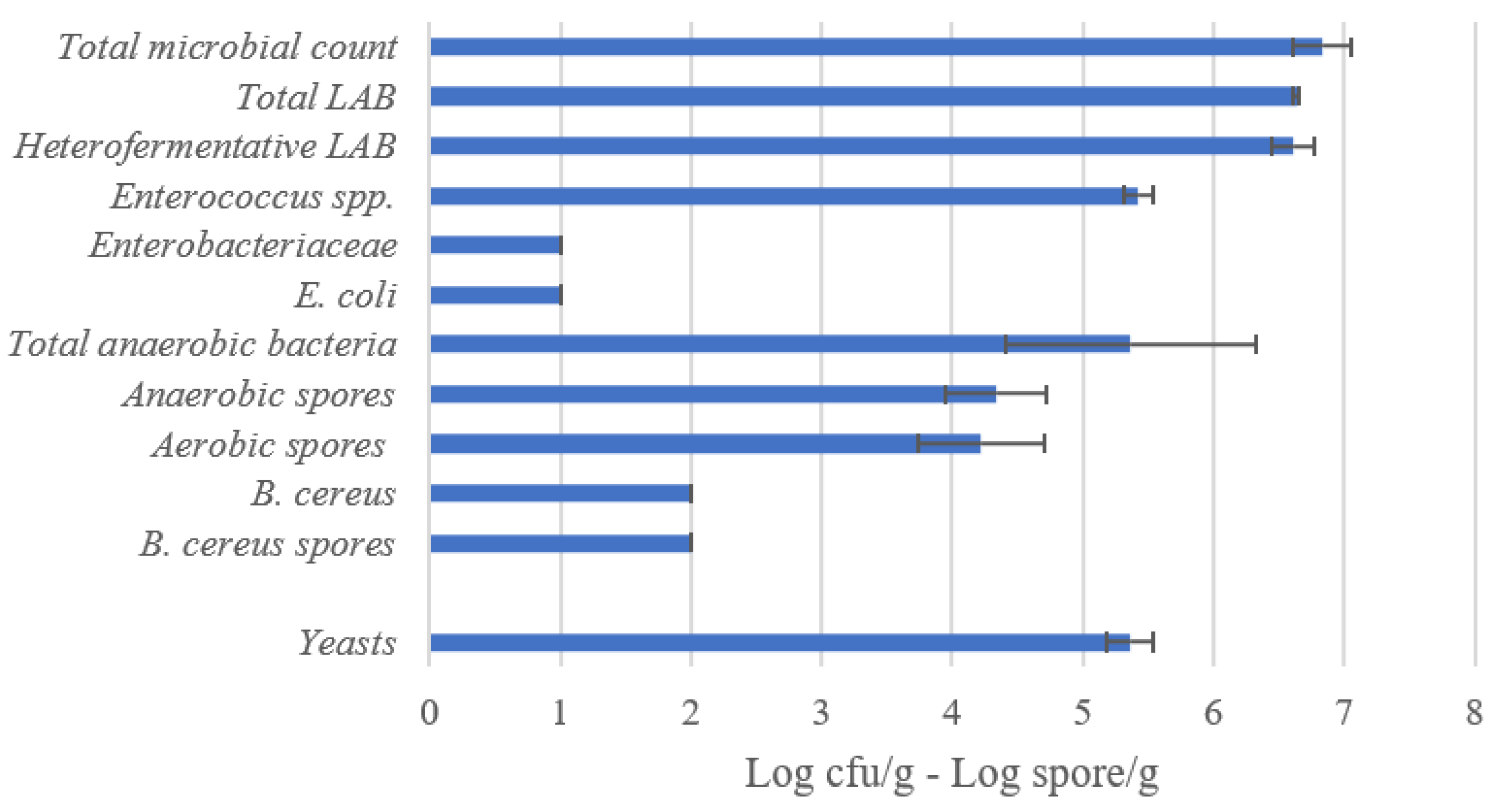
3.1.1. Oat Okara Flour
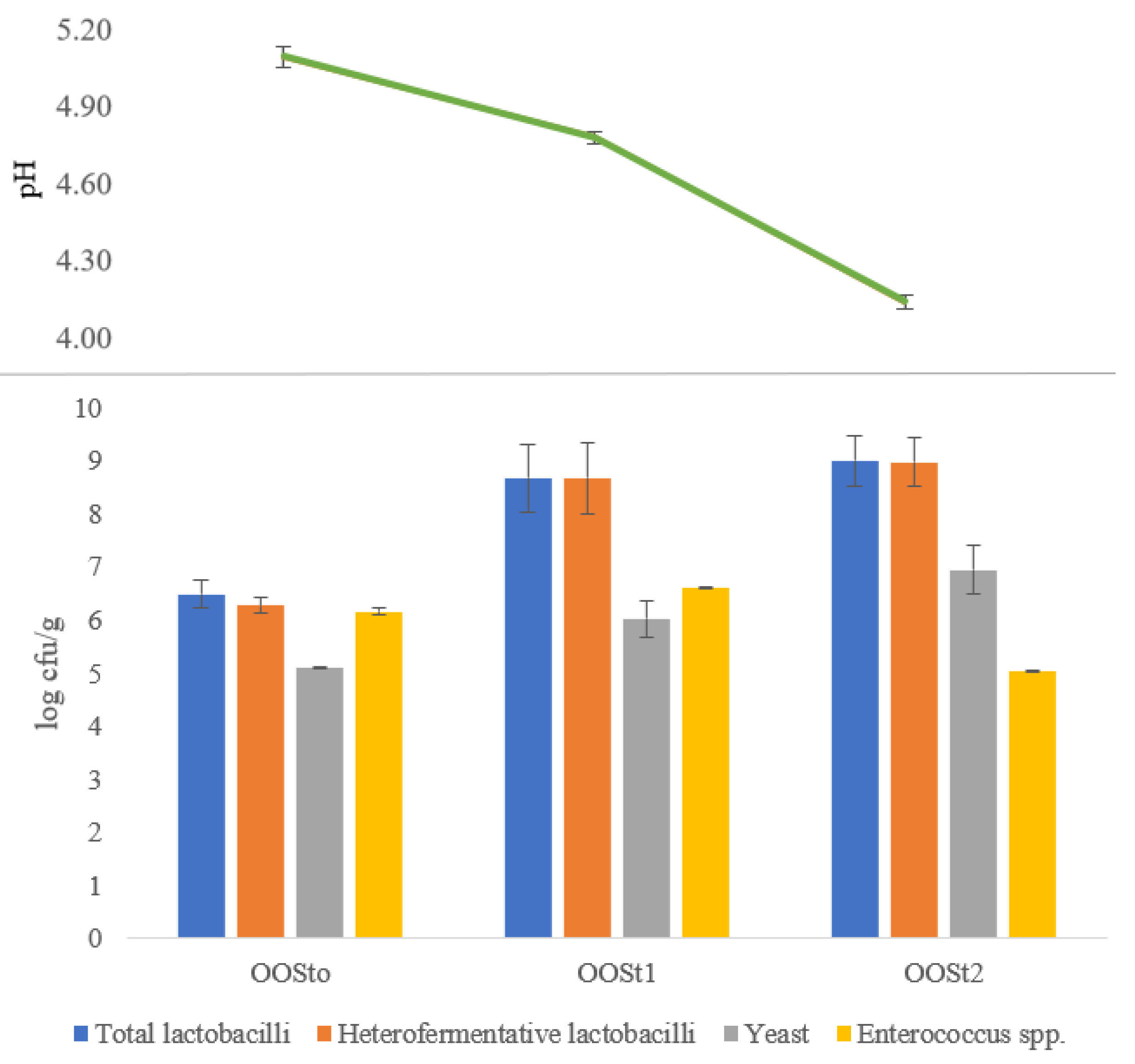
3.1.2. Microbiological Count of Oat Okara Sourdoughs
3.2. Molecular Identification and Characterization of the Isolates
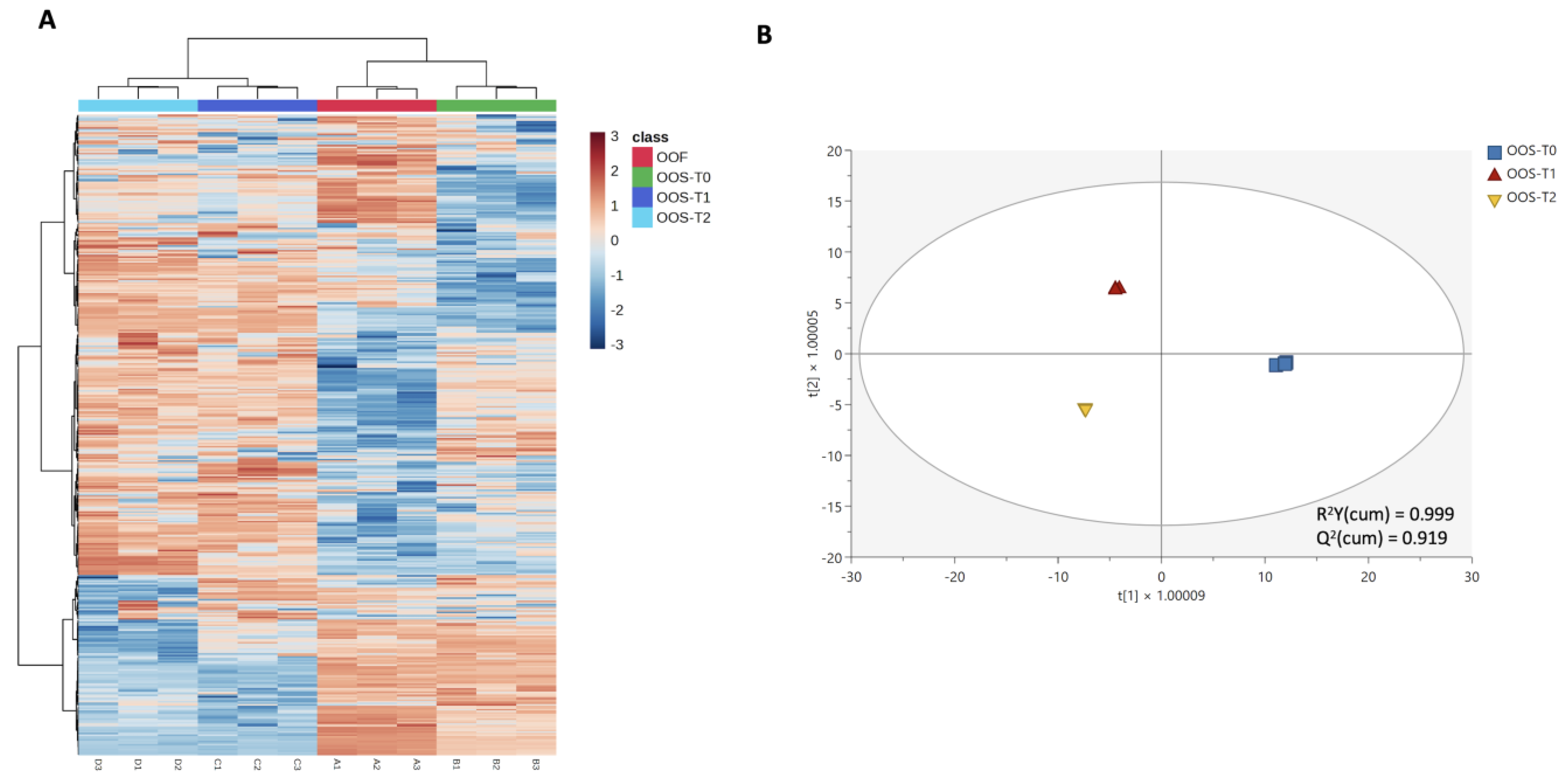
3.3. Untargeted Profiling by High-Resolution Mass Spectrometry (HRMS)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pasarín, V.; Viinikainen, T. Enabling a Legal Environment for the Prevention and Reduction of Food Loss and Waste; Legal Brief 9; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022; pp. 1–10. [Google Scholar]
- Scialabba, N.E. Food Wastage Footprint Full-Cost Accounting: Final Report; Food and Agriculture Organization of the United Na-tions: Rome, Italy, 2014; pp. 1–98, e-ISBN 978-92-5-108513-4. [Google Scholar]
- Vong, W.C.; Yang, K.L.C.A.; Liu, S.-Q. Okara (soybean residue) biotransformation by yeast Yarrowia lipolytica. Int. J. Food Microbiol. 2016, 235, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Čechp, M.; Hercp, P.; Ivanišováp, E.; Kolesárováp, A.; Urminskáp, D.; Grygorievap, O. Okara-by-product from soy processing: Characteristic, properties, benefits, and potential perspectives for industry. Int. J. Exp. Res. Rev. 2022, 28, 66–83. [Google Scholar] [CrossRef]
- Swallah, M.S.; Fan, H.; Wang, S.; Yu, H.; Piao, C. Prebiotic impacts of soybean residue (Okara) on eubiosis/dysbiosis condition of the gut and the possible effects on liver and kidney functions. Molecules 2021, 26, 326. [Google Scholar] [CrossRef] [PubMed]
- Mok, W.K.; Tan, Y.X.; Chen, W.N. Evaluating the potential of Bacillus subtilis fermented okara as a functional food ingredient through in vitro digestion and fermentation. Food Biotechnol. 2021, 35, 136–157. [Google Scholar] [CrossRef]
- Gupta, S.; Lee, J.J.L.; Chen, W.N. Analysis of Improved Nutritional Composition of Potential Functional Food (Okara) after Probiotic Solid-State Fermentation. J. Agric. Food Chem. 2018, 66, 5373–5381. [Google Scholar] [CrossRef]
- Li, B.; Qiao, M.; Lu, F. Composition, Nutrition, and Utilization of Okara (Soybean Residue). Food Rev. Int. 2012, 28, 231–252. [Google Scholar] [CrossRef]
- Fuentes, A.P.C.; Bengoa, A.; Gagliarini, N.; Abraham, A.; Pla, M.F.d.E.; Flores, S.K. Physicochemical and functional characterisation of a food ingredient based on okara containing probiotics. Food Bioprod. Process. 2022, 135, 74–86. [Google Scholar] [CrossRef]
- Colletti, A.; Attrovio, A.; Boffa, L.; Mantegna, S.; Cravotto, G. Valorisation of by-products from soybean (Glycine max (L.) Merr.) processing. Molecules 2020, 25, 2129. [Google Scholar] [CrossRef]
- Paz, G.M.; King, J.M.; Prinyawiwatkul, W. High Protein Rice Flour in the Development of Gluten-Free Bread. J. Culin. Sci. Technol. 2020, 19, 315–330. [Google Scholar] [CrossRef]
- Holopainen-Mantila, U.; Vanhatalo, S.; Lehtinen, P.; Sozer, N. Oats as a source of nutritious alternative protein. J. Cereal Sci. 2024, 116, 103862. [Google Scholar] [CrossRef]
- Mel, R.; Malalgoda, M. Oat protein as a novel protein ingredient: Structure, functionality, and factors impacting utilization. Cereal Chem. 2021, 99, 21–36. [Google Scholar] [CrossRef]
- Shewry, P.R.; Beaudoin, F.; Jenkins, J.; Griffiths-Jones, S.; Mills, E.N.C. Plant protein families and their relationships to food allergy. Biochem. Soc. Trans. 2002, 30, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Helstad, A.; Marefati, A.; Ahlström, C.; Rayner, M.; Purhagen, J.; Östbring, K. High-Pressure Pasteurization of Oat Okara. Foods 2023, 12, 4070. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, L.; Zheng, X.; Huang, Q.; Farag, M.A.; Zhu, R.; Zhao, C. Emerging applications of metabolomics in food science and future trends. Food Chem. X 2022, 16, 100500. [Google Scholar] [CrossRef]
- Ferri, M.; Serrazanetti, D.I.; Tassoni, A.; Baldissarri, M.; Gianotti, A. Improving the functional and sensorial profile of cereal-based fermented foods by selecting Lactobacillus plantarum strains via a metabolomics approach. Food Res. Int. 2016, 89, 1095–1105. [Google Scholar] [CrossRef]
- Balestra, F.; Laghi, L.; Saa, D.T.; Gianotti, A.; Rocculi, P.; Pinnavaia, G. Physico-chemical and metabolomic characterization of KAMUT® Khorasan and durum wheat fermented dough. Food Chem. 2015, 187, 451–459. [Google Scholar] [CrossRef]
- ISO 15214:1998 (En); Microbiology of Food Animal Feeding Stuffs Horizontal Method for the Enumeration of Mesophilic Lactic Acid Bacteria Colony-Count Technique at 30 Degrees, C. International Organisation for Standardization (ISO): Geneva, Switzerland, 1998. Available online: https://www.iso.org/standard/26853.html (accessed on 12 July 2024).
- Johansson, M.L.; Quednau, M.; Molin, G.; Ahrné, S. Randomly amplified polymorphic DNA (RAPD) for rapid typing of Lactobacillus plantarum strains. Lett. Appl. Microbiol. 1995, 21, 155–159. [Google Scholar] [CrossRef]
- Di Cello, F.; Fani, R. A molecular strategy for the study of natural bacterial communities by PCR-based techniques. Minerva Biotecnol. 1996, 8, 126–134. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR—Protocols and Applications—A Laboratory Manual; Academic Press: Cambridge, MA, USA, 1990; Chapter 38; pp. 315–322. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Evol. Microbiol. 1999, 49, 329–337. [Google Scholar] [CrossRef]
- Minervini, F.; Di Cagno, R.; Lattanzi, A.; De Angelis, M.; Antonielli, L.; Cardinali, G.; Cappelle, S.; Gobbetti, M. Lactic acid bacterium and yeast microbiotas of 19 sourdoughs used for traditional/typical Italian breads: Interactions between ingredients and microbial species diversity. Appl. Environ. Microbiol. 2012, 78, 1251–1264. [Google Scholar] [CrossRef]
- Iurlina, M.O.; Saiz, A.I.; Fuselli, S.R.; Fritz, R. Prevalence of Bacillus spp. in different food products collected in Argentina. LWT 2006, 39, 105–110. [Google Scholar] [CrossRef]
- Fangio, M.F.; Roura, S.I.; Fritz, R. Isolation and identification of Bacillus spp. and related genera from different starchy foods. J. Food Sci. 2010, 75, M218–M221. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.P.M.; Stradiotto, G.C.; Freire, L.; Alvarenga, V.O.; Crucello, A.; Morassi, L.L.; Silva, F.P.; Sant’ana, A.S. Occurrence and enumeration of rope-producing spore forming bacteria in flour and their spoilage potential in different bread formulations. LWT 2020, 133, 110108. [Google Scholar] [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough Microbiome Comparison and Benefits. Microorganisms 2021, 9, 1355. [Google Scholar] [CrossRef]
- Boreczek, J.; Litwinek, D.; Żylińska-Urban, J.; Izak, D.; Buksa, K.; Gawor, J.; Gromadka, R.; Bardowski, J.K.; Kowalczyk, M. Bacterial community dynamics in spontaneous sourdoughs made from wheat, spelt, and rye wholemeal flour. Microbiologyopen 2020, 9, e1009. [Google Scholar] [CrossRef]
- Hüttner, E.K.; Bello, F.D.; Arendt, E.K. Identification of lactic acid bacteria isolated from oat sourdoughs and investigation into their potential for the improvement of oat bread quality. Eur. Food Res. Technol. 2010, 230, 849–857. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L.; López, C.C.; Felis, G.E.; Mastrangelo, M.; Suzzi, G. A taxonomic survey of lactic acid bacteria isolated from wheat (Triticum durum) kernels and non-conventional flours. Syst. Appl. Microbiol. 2007, 30, 561–571. [Google Scholar] [CrossRef]
- Alfonzo, A.; Miceli, C.; Nasca, A.; Franciosi, E.; Ventimiglia, G.; Di Gerlando, R.; Tuohy, K.; Francesca, N.; Moschetti, G.; Settanni, L. Monitoring of wheat lactic acid bacteria from the field until the first step of dough fermentation. Food Microbiol. 2017, 62, 256–269. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L.; Valmorri, S.; Mastrangelo, M.; Suzzi, G. Identification of subdominant sourdough lactic acid bacteria and their evolution during laboratory-scale fermentations. Food Microbiol. 2007, 24, 592–600. [Google Scholar] [CrossRef]
- García-Béjar, B.; Fernández-Pacheco, P.; Carreño-Domínguez, J.; Briones, A.; Arévalo-Villena, M. Identification and biotechnological characterisation of yeast microbiota involved in spontaneous fermented wholegrain sourdoughs. J. Sci. Food Agric. 2023, 103, 7683–7693. [Google Scholar] [CrossRef]
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty years of knowledge on sourdough fermentation: A systematic review. Trends Food Sci. Technol. 2020, 108, 71–83. [Google Scholar] [CrossRef]
- Galli, V.; Venturi, M.; Coda, R.; Maina, N.H.; Granchi, L. Isolation and characterization of indigenous Weissella confusa for in situ bacterial exopolysaccharides (EPS) production in chickpea sourdough. Food Res. Int. 2020, 138, 109785. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi-Kouchesfahani, M.; Hamidi-Esfahani, Z.; Azizi, M.H. Isolation and identification of lactic acid bacteria with phytase activity from sourdough. Food Sci. Nutr. 2019, 7, 3700–3708. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.-H.; Park, J.; Sung, J.M.; Choi, E.-J.; Choi, Y.-S.; Park, J.-D. Characterization of Lactic Acid Bacteria and Yeast from Grains as Starter Cultures for Gluten-Free Sourdough. Foods 2023, 12, 4367. [Google Scholar] [CrossRef]
- Maidana, S.D.; Ficoseco, C.A.; Bassi, D.; Cocconcelli, P.S.; Puglisi, E.; Savoy, G.; Vignolo, G.; Fontana, C. Biodiversity and technological-functional potential of lactic acid bacteria isolated from spontaneously fermented chia sourdough. Int. J. Food Microbiol. 2020, 316, 108425. [Google Scholar] [CrossRef]
- Divisekera, D.M.W.D.; Samarasekera, J.K.R.R.; Hettiarachchi, C.; Gooneratne, J.; Choudhary, M.I.; Gopalakrishnan, S.; Wahab, A.-T. Lactic acid bacteria isolated from fermented flour of finger millet, its probiotic attributes and bioactive properties. Ann. Microbiol. 2018, 69, 79–92. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Manini, F.; Brasca, M.; Plumed-Ferrer, C.; Morandi, S.; Erba, D.; Casiraghi, M.C. Study of the chemical changes and evolution of microbiota during sourdough like fermentation of wheat bran. Cereal Chem. 2014, 91, 342–349. [Google Scholar] [CrossRef]
- Hajinia, F.; Sadeghi, A.; Mahoonak, A.S. The use of antifungal oat-sourdough lactic acid bacteria to improve safety and technological functionalities of the supplemented wheat bread. J. Food Saf. 2020, 41, e12873. [Google Scholar] [CrossRef]
- Abedfar, A.; Hosseininezhad, M.; Sadeghi, A.; Raeisi, M.; Feizy, J. Investigation on “spontaneous fermentation” and the productivity of microbial exopolysaccharides by Lactobacillus plantarum and Pediococcus pentosaceus isolated from wheat bran sourdough. LWT 2018, 96, 686–693. [Google Scholar] [CrossRef]
- Iosca, G.; De Vero, L.; Di Rocco, G.; Perrone, G.; Gullo, M.; Pulvirenti, A. Anti-Spoilage Activity and Exopolysaccharides Production by Selected Lactic Acid Bacteria. Foods 2022, 11, 1914. [Google Scholar] [CrossRef] [PubMed]
- Alfonzo, A.; Ventimiglia, G.; Corona, O.; Di Gerlando, R.; Gaglio, R.; Francesca, N.; Moschetti, G.; Settanni, L. Diversity and technological potential of lactic acid bacteria of wheat flours. Food Microbiol. 2013, 36, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Lancetti, R.; Sciarini, L.; Pérez, G.T.; Salvucci, E. Technological Performance and Selection of Lactic Acid Bacteria Isolated from Argentinian Grains as Starters for Wheat Sourdough. Curr. Microbiol. 2020, 78, 255–264. [Google Scholar] [CrossRef]
- Salvucci, E.; LeBlanc, J.G.; Pérez, G. Technological properties of Lactic acid bacteria isolated from raw cereal material. LWT 2016, 70, 185–191. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Gioulatos, S.; Tsakalidou, E.; Kalantzopoulos, G. Interactions between Saccharomyces cerevisiae and lactic acid bacteria in sourdough. Process. Biochem. 2006, 41, 2429–2433. [Google Scholar] [CrossRef]
- Syrokou, M.K.; Themeli, C.; Paramithiotis, S.; Mataragas, M.; Bosnea, L.; Argyri, A.A.; Chorianopoulos, N.G.; Skandamis, P.N.; Drosinos, E.H. Microbial ecology of greek wheat sourdoughs, identified by a culture-dependent and a culture-independent approach. Foods 2020, 9, 1603. [Google Scholar] [CrossRef]
- EL Boujamaai, M.; Mannani, N.; Aloui, A.; Errachidi, F.; Ben Salah-Abbès, J.; Riba, A.; Abbès, S.; Rocha, J.M.; Bartkiene, E.; Brabet, C.; et al. Biodiversity and biotechnological properties of lactic acid bacteria isolated from traditional Moroccan sourdoughs. World J. Microbiol. Biotechnol. 2023, 39, 1–11. [Google Scholar] [CrossRef]
- Cardinali, F.; Garofalo, C.; Reale, A.; Boscaino, F.; Osimani, A.; Milanović, V.; Taccari, M.; Aquilanti, L. Liquid sourdough from stone-ground soft wheat (Triticum aestivum) flour: Development and exploitation in the breadmaking process. Food Res. Int. 2022, 161, 111796. [Google Scholar] [CrossRef]
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.-M.; Weckx, S. Microbial ecology of sourdough fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef]
- Kahve, H.I. In Vitro Evaluation of the Technological and Probiotic Potential of Pichia kudriavzevii Strains Isolated from Traditional Fermented Foods. Curr. Microbiol. 2023, 80, 1–9. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, H.; Xi, J.; Jin, Y.; Chen, Y.; Guo, L.; Jin, Z.; Xu, X. Improving bread aroma using low-temperature sourdough fermentation. Food Biosci. 2020, 37, 100704. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, Y.; Zhao, Q.; Xu, X.; Xu, D. The contributions of Pichia kudriavzevii EP1 to sourdough: Acid utilization, oligosaccharides and peptides production and aroma enhancement. Food Biosci. 2024, 59, 104225. [Google Scholar] [CrossRef]
- Bourbon-Melo, N.; Palma, M.; Rocha, M.P.; Ferreira, A.; Bronze, M.R.; Elias, H.; Sá-Correia, I. Use of Hanseniaspora guilliermondii and Hanseniaspora opuntiae to enhance the aromatic profile of beer in mixed-culture fermentation with Saccharomyces cerevisiae. Food Microbiol. 2020, 95, 103678. [Google Scholar] [CrossRef] [PubMed]
- Papalexandratou, Z.; Lefeber, T.; Bahrim, B.; Lee, O.S.; Daniel, H.-M.; De Vuyst, L. Hanseniaspora opuntiae, Saccharomyces cerevisiae, Lactobacillus fermentum, and Acetobacter pasteurianus predominate during well-performed Malaysian cocoa bean box fermentations, underlining the importance of these microbial species for a successful cocoa bean fermentation process. Food Microbiol. 2013, 35, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Sevgili, A.; Can, C.; Ceyhan, D.I.; Erkmen, O. Molecular identification of LAB and yeasts from traditional sourdoughs and their impacts on the sourdough bread quality characteristics. Curr. Res. Food Sci. 2023, 6, 100479. [Google Scholar] [CrossRef] [PubMed]
- Hitayezu, R.; Baakdah, M.M.; Kinnin, J.; Henderson, K.; Tsopmo, A. Antioxidant activity, avenanthramide and phenolic acid contents of oat milling fractions. J. Cereal Sci. 2015, 63, 35–40. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Djorgbenoo, R.; Djorgbenoo, R.; Hu, J.; Hu, J.; Hu, C.; Hu, C.; Sang, S.; Sang, S. Fermented Oats as a Novel Functional Food. Nutrients 2023, 15, 3521. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Cai, Y.-Z.; Zhang, Y. Natural phenolic compounds from medicinal herbs and dietary plants: Potential use for cancer prevention. Nutr. Cancer 2009, 62, 1–20. [Google Scholar] [CrossRef]
- Boudaoud, S.; Aouf, C.; Devillers, H.; Sicard, D.; Segond, D. Sourdough yeast-bacteria interactions can change ferulic acid metabolism during fermentation. Food Microbiol. 2021, 98, 103790. [Google Scholar] [CrossRef]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Hakim, B.N.A.; Xuan, N.J.; Oslan, S.N.H. A Comprehensive Review of Bioactive Compounds from Lactic Acid Bacteria: Potential Functions as Functional Food in Dietetics and the Food Industry. Foods 2023, 12, 2850. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhou, C.; Zhang, T.; Jiang, B.; Mu, W. Short communication: 3-Phenyllactic acid production in milk by Pediococcus pentosaceus SK25 during laboratory fermentation process. J. Dairy Sci. 2015, 98, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef]
- Singh, R.; Pophaly, S.D.; Kaushik, J.K.; Tomar, S.K. Current status and emerging role of glutathione in food grade lactic acid bacteria. Microb. Cell Factories 2012, 11, 114. [Google Scholar] [CrossRef]
- Premi, L.; Rocchetti, G.; Rossetti, C.; Dallolio, M.; Lucini, L.; Rebecchi, A. Coagulase-negative staphylococci enhance the colour of fermented meat through a complex cross-talk between the arginase and nitric oxide synthase activities. LWT 2024, 202, 116333. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Class | OOF (mg/100 g DM) |
|---|---|
| Anthocyanins (Cyanidin Eq.) | 4.95 ± 0.23 |
| Flavan-3-ols (Catechin Eq.) | 3.22 ± 0.42 |
| Flavonols (Quercetin Eq.) | 2.70 ± 0.08 |
| Other flavonoids (Luteolin Eq.) | 6.00 ± 0.12 |
| Other phenolics (Oleuropein Eq.) | 16.56 ± 1.74 |
| Phenolic acids (Ferulic acid Eq.) | 29.05 ± 0.21 |
| Stilbenes (Resveratrol Eq.) | 1.28 ± 0.14 |
| Total phenolics | 63.76 ± 10.14 |
| Metabolites | Log2FC OOS-t1 vs. OOS-t0 | Log2FC OOS-t2 vs. OOS-t0 | VIP Score |
|---|---|---|---|
| 6-Phosphogluconic acid | 3.62 | 5.94 | 1.83 |
| Reduced Glutathione | −2.79 | −4.96 | 1.73 |
| Diacetyl | 1.82 | 4.87 | 1.70 |
| 5-Hydroxyindole-3-acetic acid | −1.18 | −3.54 | 1.42 |
| Citrulline | 1.18 | −1.82 | 1.68 |
| 2,3-Diaminopropionic acid | 0.89 | 1.63 | <1 |
| 3-phenyllactic acid | 1.14 | 1.62 | <1 |
| Lactoylglutathione | 0.45 | 1.54 | <1 |
| Indole-3-acetic acid | 1.38 | 1.42 | <1 |
| Lactic acid | 0.59 | 1.39 | <1 |
| NADH | 1.76 | 1.37 | <1 |
| trans-2-Hexenal | 1.26 | 1.30 | <1 |
| 2-Hydroxy-3-methylbutyric acid | 1.27 | 1.20 | <1 |
| 3-Indolepropionic acid | 2.35 | 1.14 | 1.00 |
| 4-Aminobutyrate | 1.87 | 1.02 | 1.03 |
| Ornithine | 0.92 | 0.77 | <1 |
| Butyric acid | 0.44 | 0.73 | <1 |
| Linolenic acid | 0.63 | 0.61 | <1 |
| 2-Hydroxyisobutyric acid | 0.62 | 0.56 | <1 |
| NAD+ | −0.38 | 0.50 | <1 |
| Oxidized Glutathione | 1.37 | −0.41 | 1.43 |
| 3-hydroxy-3-methylbutanoic acid | 0.63 | 0.36 | <1 |
| Succinic acid | 0.17 | 0.31 | <1 |
| 9-hydroxy-10,12-octadecadienoic acid | −2.59 | 0.25 | 1.50 |
| Citric acid | −2.54 | 0.22 | 1.64 |
| Propionic acid | −0.42 | −0.19 | <1 |
| p-Hydroxyphenyllactic acid | 0.14 | 0.19 | <1 |
| 8-4′-Dehydrodiferulic acid | −3.69 | −8.79 | 2.43 |
| p-Coumaroyl malic acid | 3.56 | 5.41 | 1.67 |
| 1,5-Diferuloylquinic acid | 2.47 | 4.56 | 1.65 |
| Sinapic acid | 1.69 | 4.54 | 1.65 |
| trans-Ferulic acid | 1.11 | −1.59 | 1.50 |
| p-Coumaric acid | 1.83 | 3.91 | 1.41 |
| Isoferulic Acid | 1.32 | 2.84 | 1.18 |
| Vanillic acid | −0.49 | −1.64 | 1.04 |
| 24-Methylcholestanol ferulate | −7.74 | −7.34 | 2.20 |
| Alanine | −0.54 | −1.43 | 1.01 |
| Tryptophan | 0.45 | −1.11 | 1.21 |
| Pyroglutamic acid | 0.61 | −0.95 | 1.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meanti, F.; Mussio, C.; Rocchetti, G.; Rebecchi, A.; Lucini, L.; Morelli, L. Oat Okara Fermentation: New Insights into the Microbiological and Metabolomic Characterization. Fermentation 2024, 10, 545. https://doi.org/10.3390/fermentation10110545
Meanti F, Mussio C, Rocchetti G, Rebecchi A, Lucini L, Morelli L. Oat Okara Fermentation: New Insights into the Microbiological and Metabolomic Characterization. Fermentation. 2024; 10(11):545. https://doi.org/10.3390/fermentation10110545
Chicago/Turabian StyleMeanti, Federica, Chiara Mussio, Gabriele Rocchetti, Annalisa Rebecchi, Luigi Lucini, and Lorenzo Morelli. 2024. "Oat Okara Fermentation: New Insights into the Microbiological and Metabolomic Characterization" Fermentation 10, no. 11: 545. https://doi.org/10.3390/fermentation10110545
APA StyleMeanti, F., Mussio, C., Rocchetti, G., Rebecchi, A., Lucini, L., & Morelli, L. (2024). Oat Okara Fermentation: New Insights into the Microbiological and Metabolomic Characterization. Fermentation, 10(11), 545. https://doi.org/10.3390/fermentation10110545