Expression of Long Non-Coding RNAs by Human Retinal Müller Glial Cells Infected with Clonal and Exotic Virulent Toxoplasma gondii

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Expression of Long Non-Coding RNA in Human Retinal Müller Cells
2.2. Time Course of T. gondii Growth in Human Retinal Müller Cells
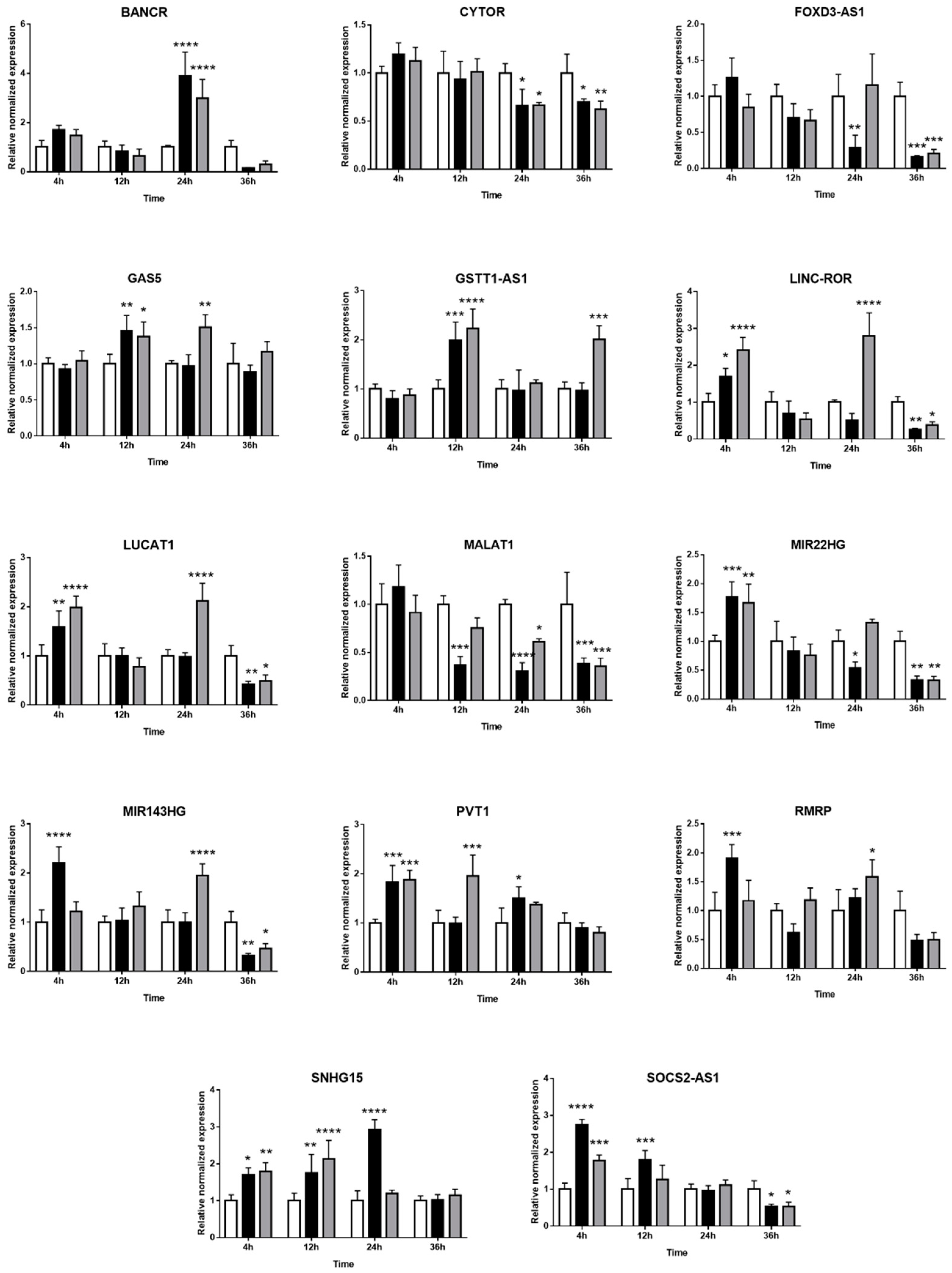
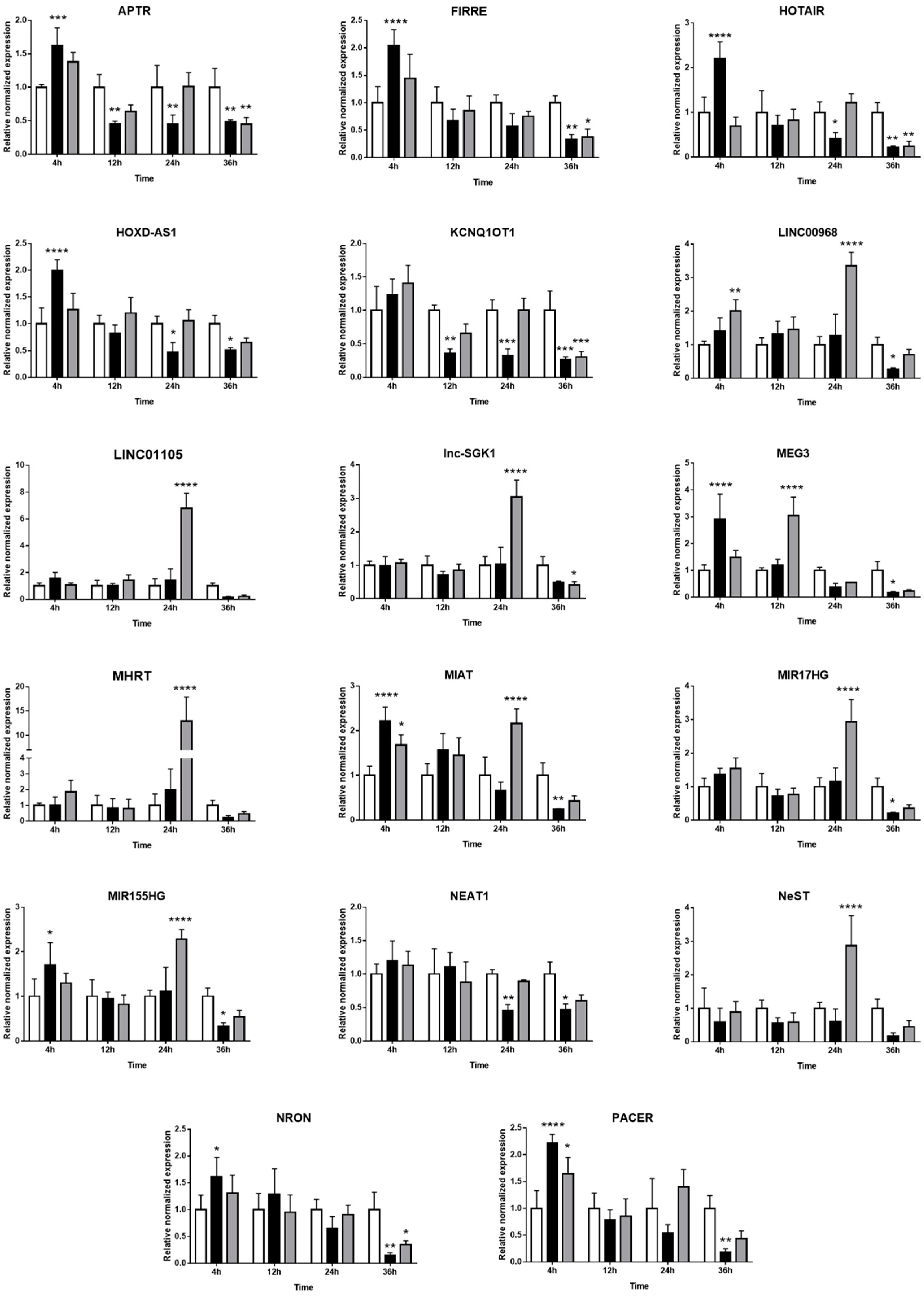
2.3. Expression of Long Non-Coding RNAs in Human Retinal Müller Cells during T. gondii Infection
3. Discussion
4. Materials and Methods
4.1. Human Subjects
4.2. Cell Culture
4.3. Parasite Culture
4.4. Infection of MIO-M1 Cells
4.5. RNA Isolation and Reverse Transcription
4.6. Polymerase Chain Reaction
4.7. Quantitative Real-Time Polymerase Chain Reaction
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Furtado, J.M.; Winthrop, K.L.; Butler, N.J.; Smith, J.R. Ocular toxoplasmosis I: Parasitology, epidemiology and public health. Clin. Exp. Ophthalmol. 2013, 41, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Butler, N.J.; Furtado, J.M.; Winthrop, K.L.; Smith, J.R. Ocular toxoplasmosis II: Clinical features, pathology and management. Clin. Exp. Ophthalmol. 2013, 41, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, M.C.; Aleixo, A.L.; Benchimol, E.I.; Leandro, A.C.; das Neves, L.B.; Vicente, R.T.; Bonecini-Almeida Mda, G.; Amendoeira, M.R. The IFN-gamma +874T/A gene polymorphism is associated with retinochoroiditis toxoplasmosis susceptibility. Mem. Inst. Oswaldo Cruz 2009, 104, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, C.A.; Moreira, P.R.; Bessa, T.F.; Costa, G.C.; Dutra, W.O.; Campos, W.R.; Orefice, F.; Young, L.H.; Teixeira, A.L. Interleukin-6 gene polymorphism (−174 G/C) is associated with toxoplasmic retinochoroiditis. Acta Ophthalmol. 2013, 91, e311–e314. [Google Scholar] [CrossRef] [PubMed]
- De-la-Torre, A.; Sauer, A.; Pfaff, A.W.; Bourcier, T.; Brunet, J.; Speeg-Schatz, C.; Ballonzoli, L.; Villard, O.; Ajzenberg, D.; Sundar, N.; et al. Severe South American ocular toxoplasmosis is associated with decreased Ifn-gamma/Il-17a and increased Il-6/Il-13 intraocular levels. PLoS Negl. Trop. Dis. 2013, 7, e2541. [Google Scholar] [CrossRef] [PubMed]
- Howe, D.K.; Sibley, L.D. Toxoplasma gondii comprises three clonal lineages: Correlation of parasite genotype with human disease. J. Infect. Dis. 1995, 172, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Dubey, J.P.; Su, C.; Ajioka, J.W.; Rosenthal, B.M.; Sibley, L.D. Genetic analyses of atypical Toxoplasma gondii strains reveal a fourth clonal lineage in North America. Int. J. Parasitol. 2011, 41, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Shwab, E.K.; Zhu, X.Q.; Majumdar, D.; Pena, H.F.; Gennari, S.M.; Dubey, J.P.; Su, C. Geographical patterns of Toxoplasma gondii genetic diversity revealed by multilocus PCR-RFLP genotyping. Parasitology 2014, 141, 453–461. [Google Scholar] [CrossRef]
- Minot, S.; Melo, M.B.; Li, F.; Lu, D.; Niedelman, W.; Levine, S.S.; Saeij, J.P. Admixture and recombination among Toxoplasma gondii lineages explain global genome diversity. Proc. Natl. Acad. Sci. USA 2012, 109, 13458–13463. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Khan, A.; Zhou, P.; Majumdar, D.; Ajzenberg, D.; Darde, M.L.; Zhu, X.Q.; Ajioka, J.W.; Rosenthal, B.M.; Dubey, J.P.; et al. Globally diverse Toxoplasma gondii isolates comprise six major clades originating from a small number of distinct ancestral lineages. Proc. Natl. Acad. Sci. USA 2012, 109, 5844–5849. [Google Scholar] [CrossRef]
- Glasner, P.D.; Silveira, C.; Kruszon-Moran, D.; Martins, M.C.; Burnier Junior, M.; Silveira, S.; Camargo, M.E.; Nussenblatt, R.B.; Kaslow, R.A.; Belfort Junior, R. An unusually high prevalence of ocular toxoplasmosis in southern Brazil. Am. J. Ophthalmol. 1992, 114, 136–144. [Google Scholar] [CrossRef]
- De-la-Torre, A.; Gonzalez, G.; Diaz-Ramirez, J.; Gomez-Marin, J.E. Screening by ophthalmoscopy for Toxoplasma retinochoroiditis in Colombia. Am. J. Ophthalmol. 2007, 143, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, A.W.; de-la-Torre, A.; Rochet, E.; Brunet, J.; Sabou, M.; Sauer, A.; Bourcier, T.; Gomez-Marin, J.E.; Candolfi, E. New clinical and experimental insights into Old World and neotropical ocular toxoplasmosis. Int. J. Parasitol. 2014, 44, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Bringmann, A. New functions of Muller cells. Glia 2013, 61, 651–678. [Google Scholar] [CrossRef] [PubMed]
- Shamsuddin, N.; Kumar, A. TLR2 mediates the innate response of retinal Muller glia to Staphylococcus aureus. J. Immunol. 2011, 186, 7089–7097. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Pandey, R.K.; Miller, L.J.; Singh, P.K.; Kanwar, M. Muller glia in retinal innate immunity: A perspective on their roles in endophthalmitis. Crit. Rev. Immunol. 2013, 33, 119–135. [Google Scholar] [CrossRef]
- Kumar, A.; Shamsuddin, N. Retinal Muller glia initiate innate response to infectious stimuli via toll-like receptor signaling. PLoS ONE 2012, 7, e29830. [Google Scholar]
- Knight, B.C.; Kissane, S.; Falciani, F.; Salmon, M.; Stanford, M.R.; Wallace, G.R. Expression analysis of immune response genes of Muller cells infected with Toxoplasma gondii. J. Neuroimmunol. 2006, 179, 126–131. [Google Scholar] [CrossRef]
- Furtado, J.M.; Ashander, L.M.; Mohs, K.; Chipps, T.J.; Appukuttan, B.; Smith, J.R. Toxoplasma gondii migration within and infection of human retina. PLoS ONE 2013, 8, e54358. [Google Scholar] [CrossRef]
- Lahmar, I.; Guinard, M.; Sauer, A.; Marcellin, L.; Abdelrahman, T.; Roux, M.; Mousli, M.; Moussa, A.; Babba, H.; Pfaff, A.W.; et al. Murine neonatal infection provides an efficient model for congenital ocular toxoplasmosis. Exp. Parasitol. 2010, 124, 190–196. [Google Scholar] [CrossRef]
- McMenamin, P.G.; Dutton, G.N.; Hay, J.; Cameron, S. The ultrastructural pathology of congenital murine toxoplasmic retinochoroiditis. Part I: The localization and morphology of Toxoplasma cysts in the retina. Exp. Eye Res. 1986, 43, 529–543. [Google Scholar] [CrossRef]
- Pavesio, C.E.; Chiappino, M.L.; Gormley, P.; Setzer, P.Y.; Nichols, B.A. Acquired retinochoroiditis in hamsters inoculated with ME 49 strain Toxoplasma. Invest. Ophthalmol. Vis. Sci. 1995, 36, 2166–2175. [Google Scholar] [PubMed]
- Song, H.B.; Jung, B.K.; Kim, J.H.; Lee, Y.H.; Choi, M.H.; Kim, J.H. Investigation of tissue cysts in the retina in a mouse model of ocular toxoplasmosis: Distribution and interaction with glial cells. Parasitol. Res. 2018, 117, 2597–2605. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Kraus, W.L. From discovery to function: The expanding roles of long noncoding RNAs in physiology and disease. Endocr. Rev. 2015, 36, 25–64. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Kim, H.W.; Nam, J.W. The small peptide world in long noncoding RNAs. Brief Bioinform. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dhir, A.; Dhir, S.; Proudfoot, N.J.; Jopling, C.L. Microprocessor mediates transcriptional termination of long noncoding RNA transcripts hosting microRNAs. Nat. Struct. Mol. Biol. 2015, 22, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Hadjicharalambous, M.R.; Lindsay, M.A. Long non-coding RNAs and the innate immune response. Noncoding RNA 2019, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Agliano, F.; Rathinam, V.A.; Medvedev, A.E.; Vanaja, S.K.; Vella, A.T. Long noncoding RNAs in host-pathogen interactions. Trends Immunol. 2019, 40, 492–510. [Google Scholar] [CrossRef]
- Chen, Y.G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef]
- Fernandes, J.C.R.; Acuna, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long non-coding RNAs in the regulation of gene expression: Physiology and disease. Noncoding RNA 2019, 5, 17. [Google Scholar] [CrossRef]
- Mumtaz, P.T.; Bhat, S.A.; Ahmad, S.M.; Dar, M.A.; Ahmed, R.; Urwat, U.; Ayaz, A.; Shrivastava, D.; Shah, R.A.; Ganai, N.A. LncRNAs and immunity: Watchdogs for host pathogen interactions. Biol. Proced. Online 2017, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Mathy, N.W.; Chen, X.M. Long non-coding RNAs (lncRNAs) and their transcriptional control of inflammatory responses. J. Biol. Chem. 2017, 292, 12375–12382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, X. Long noncoding RNAs in innate immunity. Cell. Mol. Immunol. 2016, 13, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. The NeST long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-gamma locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, C.; Ma, X.; Geng, G.; Liu, B.; Zhang, Y.; Zhang, S.; Zhong, F.; Liu, C.; Yin, Y.; et al. Long noncoding RNA NRON contributes to HIV-1 latency by specifically inducing tat protein degradation. Nat. Commun. 2016, 7, 11730. [Google Scholar] [CrossRef] [PubMed]
- Lahmar, I.; Pfaff, A.W.; Marcellin, L.; Sauer, A.; Moussa, A.; Babba, H.; Candolfi, E. Muller cell activation and photoreceptor depletion in a mice model of congenital ocular toxoplasmosis. Exp. Parasitol. 2014, 144, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Huang, L.; Wei, Q.; Zhang, Y.; Zhang, S.; Zhang, W.; Cai, L.; Liang, S. Microarray analysis of long non-coding RNA expression profiles uncovers a Toxoplasma-induced negative regulation of host immune signaling. Parasit. Vectors 2018, 11, 174. [Google Scholar] [CrossRef]
- Menard, K.L.; Haskins, B.E.; Colombo, A.P.; Denkers, E.Y. Toxoplasma gondii manipulates expression of host long noncoding RNA during intracellular infection. Sci. Rep. 2018, 8, 15017. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Suzuki, Y.; Orellana, M.A.; Schreiber, R.D.; Remington, J.S. Interferon-gamma: The major mediator of resistance against Toxoplasma gondii. Science 1988, 240, 516–518. [Google Scholar] [CrossRef]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-gamma at the crossroads of tumor immune surveillance or evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef]
- Spurlock, C.F., 3rd; Crooke, P.S., 3rd; Aune, T.M. Biogenesis and transcriptional regulation of long noncoding RNAs in the human immune system. J. Immunol. 2016, 197, 4509–4517. [Google Scholar] [CrossRef] [PubMed]
- Kuo, G.; Wu, C.Y.; Yang, H.Y. MiR-17-92 cluster and immunity. J. Formos. Med. Assoc. 2019, 118, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Jiang, Q.; Jiang, L.; Wu, J.; Zhang, Q.; Wang, J.; Feng, H.; Zang, P. Lnc-SGK1 induced by Helicobacter pylori infection and high salt diet promote Th2 and Th17 differentiation in human gastric cancer by SGK1/Jun B signaling. Oncotarget 2016, 7, 20549–20560. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Q.; Hu, S.Y.; Wang, Z.Y.; Lin, J.; Jian, S.; Dong, Y.C.; Wu, X.F.; Dai, L.; Cao, L.J. Long non-coding RNA MEG3 inhibits microRNA-125a-5p expression and induces immune imbalance of Treg/Th17 in immune thrombocytopenic purpura. Biomed. Pharm. 2016, 83, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.Y.; Wu, Y.; Lin, M.J.; Bian, T.; Xiao, Y.L.; Qin, C. LncRNA-MEG3 functions as a competing endogenous RNA to regulate Treg/Th17 balance in patients with asthma by targeting microRNA-17/ RORgammat. Biomed. Pharm. 2019, 111, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Lahmar, I.; Abou-Bacar, A.; Abdelrahman, T.; Guinard, M.; Babba, H.; Ben Yahia, S.; Kairallah, M.; Speeg-Schatz, C.; Bourcier, T.; Sauer, A.; et al. Cytokine profiles in toxoplasmic and viral uveitis. J. Infect. Dis. 2009, 199, 1239–1249. [Google Scholar] [CrossRef]
- Sauer, A.; Pfaff, A.W.; Villard, O.; Creuzot-Garcher, C.; Dalle, F.; Chiquet, C.; Pelloux, H.; Speeg-Schatz, C.; Gaucher, D.; Prevost, G.; et al. Interleukin 17A as an effective target for anti-inflammatory and antiparasitic treatment of toxoplasmic uveitis. J. Infect. Dis. 2012, 206, 1319–1329. [Google Scholar] [CrossRef]
- Sherrill, J.D.; Kiran, K.C.; Blanchard, C.; Stucke, E.M.; Kemme, K.A.; Collins, M.H.; Abonia, J.P.; Putnam, P.E.; Mukkada, V.A.; Kaul, A.; et al. Analysis and expansion of the eosinophilic esophagitis transcriptome by RNA sequencing. Genes Immun. 2014, 15, 361–369. [Google Scholar] [CrossRef]
- Li, D.Y.; Chen, W.J.; Shang, J.; Chen, G.; Li, S.K. Regulatory interactions between long noncoding RNA LINC00968 and miR-9-3p in non-small cell lung cancer: A bioinformatic analysis based on miRNA microarray, GEO and TCGA. Oncol. Lett. 2018, 15, 9487–9497. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Xu, Y.J.; Hu, H.B. Long non-coding RNA LINC00968 acts as oncogene in NSCLC by activating the Wnt signaling pathway. J. Cell. Physiol. 2018, 233, 3397–3406. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, Z.; Meng, X.; Yu, P. LncRNA LINC00968 accelerates the proliferation and fibrosis of diabetic nephropathy by epigenetically repressing p21 via recruiting EZH2. Biochem. Biophys. Res. Commun. 2018, 504, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.; Jia, L.; Duan, Y.; Li, Y.; Bao, L.; Sha, N. Long non-coding RNA BANCR promotes proliferation in malignant melanoma by regulating MAPK pathway activation. PLoS ONE 2014, 9, e100893. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhang, D.; Xu, B.; Wu, Z.; Liu, S.; Zhang, L.; Tian, Y.; Han, X.; Tian, D. Long non-coding RNA BANCR promotes proliferation and migration of lung carcinoma via MAPK pathways. Biomed. Pharm. 2015, 69, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Loewer, S.; Cabili, M.N.; Guttman, M.; Loh, Y.H.; Thomas, K.; Park, I.H.; Garber, M.; Curran, M.; Onder, T.; Agarwal, S.; et al. Large intergenic non-coding RNA-RoR modulates reprogramming of human induced pluripotent stem cells. Nat. Genet. 2010, 42, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhou, N.; Huang, J.; Liu, Q.; Fukuda, K.; Ma, D.; Lu, Z.; Bai, C.; Watabe, K.; Mo, Y.Y. The human long non-coding RNA-RoR is a p53 repressor in response to DNA damage. Cell Res. 2013, 23, 340–350. [Google Scholar] [CrossRef]
- Brunet, J.; Pfaff, A.W.; Abidi, A.; Unoki, M.; Nakamura, Y.; Guinard, M.; Klein, J.P.; Candolfi, E.; Mousli, M. Toxoplasma gondii exploits UHRF1 and induces host cell cycle arrest at G2 to enable its proliferation. Cell. Microbiol. 2008, 10, 908–920. [Google Scholar] [CrossRef]
- Molestina, R.E.; El-Guendy, N.; Sinai, A.P. Infection with Toxoplasma gondii results in dysregulation of the host cell cycle. Cell. Microbiol. 2008, 10, 1153–1165. [Google Scholar] [CrossRef]
- Angeloni, M.B.; Silva, N.M.; Castro, A.S.; Gomes, A.O.; Silva, D.A.; Mineo, J.R.; Ferro, E.A. Apoptosis and S phase of the cell cycle in BeWo trophoblastic and HeLa cells are differentially modulated by Toxoplasma gondii strain types. Placenta 2009, 30, 785–791. [Google Scholar] [CrossRef]
- Lavine, M.D.; Arrizabalaga, G. Induction of mitotic S-phase of host and neighboring cells by Toxoplasma gondii enhances parasite invasion. Mol. Biochem. Parasitol. 2009, 164, 95–99. [Google Scholar] [CrossRef]
- Wang, G.; Gao, M. Influence of Toxoplasma gondii on in vitro proliferation and apoptosis of hepatoma carcinoma H7402 cell. Asian Pac. J. Trop. Med. 2016, 9, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Limb, G.A.; Salt, T.E.; Munro, P.M.; Moss, S.E.; Khaw, P.T. In vitro characterization of a spontaneously immortalized human Muller cell line (MIO-M1). Invest. Ophthalmol. Vis. Sci. 2002, 43, 864–869. [Google Scholar] [PubMed]
- Sibley, L.D.; Boothroyd, J.C. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature 1992, 359, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Fux, B.; Su, C.; Dubey, J.P.; Darde, M.L.; Ajioka, J.W.; Rosenthal, B.M.; Sibley, L.D. Recent transcontinental sweep of Toxoplasma gondii driven by a single monomorphic chromosome. Proc. Natl. Acad. Sci. USA 2007, 104, 14872–14877. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Behnke, M.S.; Dunay, I.R.; White, M.W.; Sibley, L.D. Phenotypic and gene expression changes among clonal type I strains of Toxoplasma gondii. Eukaryot. Cell 2009, 8, 1828–1836. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Helleman, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
- Appukuttan, B.; Ashander, L.M.; Ma, Y.; Smith, J.R. Selection of reference genes for studies of human retinal endothelial cell gene expression by reverse transcription-quantitative real-time polymerase chain reaction. Gene Rep. 2018, 10, 123–134. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rochet, E.; Appukuttan, B.; Ma, Y.; Ashander, L.M.; Smith, J.R. Expression of Long Non-Coding RNAs by Human Retinal Müller Glial Cells Infected with Clonal and Exotic Virulent Toxoplasma gondii. Non-Coding RNA 2019, 5, 48. https://doi.org/10.3390/ncrna5040048
Rochet E, Appukuttan B, Ma Y, Ashander LM, Smith JR. Expression of Long Non-Coding RNAs by Human Retinal Müller Glial Cells Infected with Clonal and Exotic Virulent Toxoplasma gondii. Non-Coding RNA. 2019; 5(4):48. https://doi.org/10.3390/ncrna5040048
Chicago/Turabian StyleRochet, Elise, Binoy Appukuttan, Yuefang Ma, Liam M. Ashander, and Justine R. Smith. 2019. "Expression of Long Non-Coding RNAs by Human Retinal Müller Glial Cells Infected with Clonal and Exotic Virulent Toxoplasma gondii" Non-Coding RNA 5, no. 4: 48. https://doi.org/10.3390/ncrna5040048
APA StyleRochet, E., Appukuttan, B., Ma, Y., Ashander, L. M., & Smith, J. R. (2019). Expression of Long Non-Coding RNAs by Human Retinal Müller Glial Cells Infected with Clonal and Exotic Virulent Toxoplasma gondii. Non-Coding RNA, 5(4), 48. https://doi.org/10.3390/ncrna5040048