
Human Papillomavirus-Encoded microRNAs as Regulators of Human Gene Expression in Anal Squamous Cell Carcinoma: A Meta-Transcriptomics Study

 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
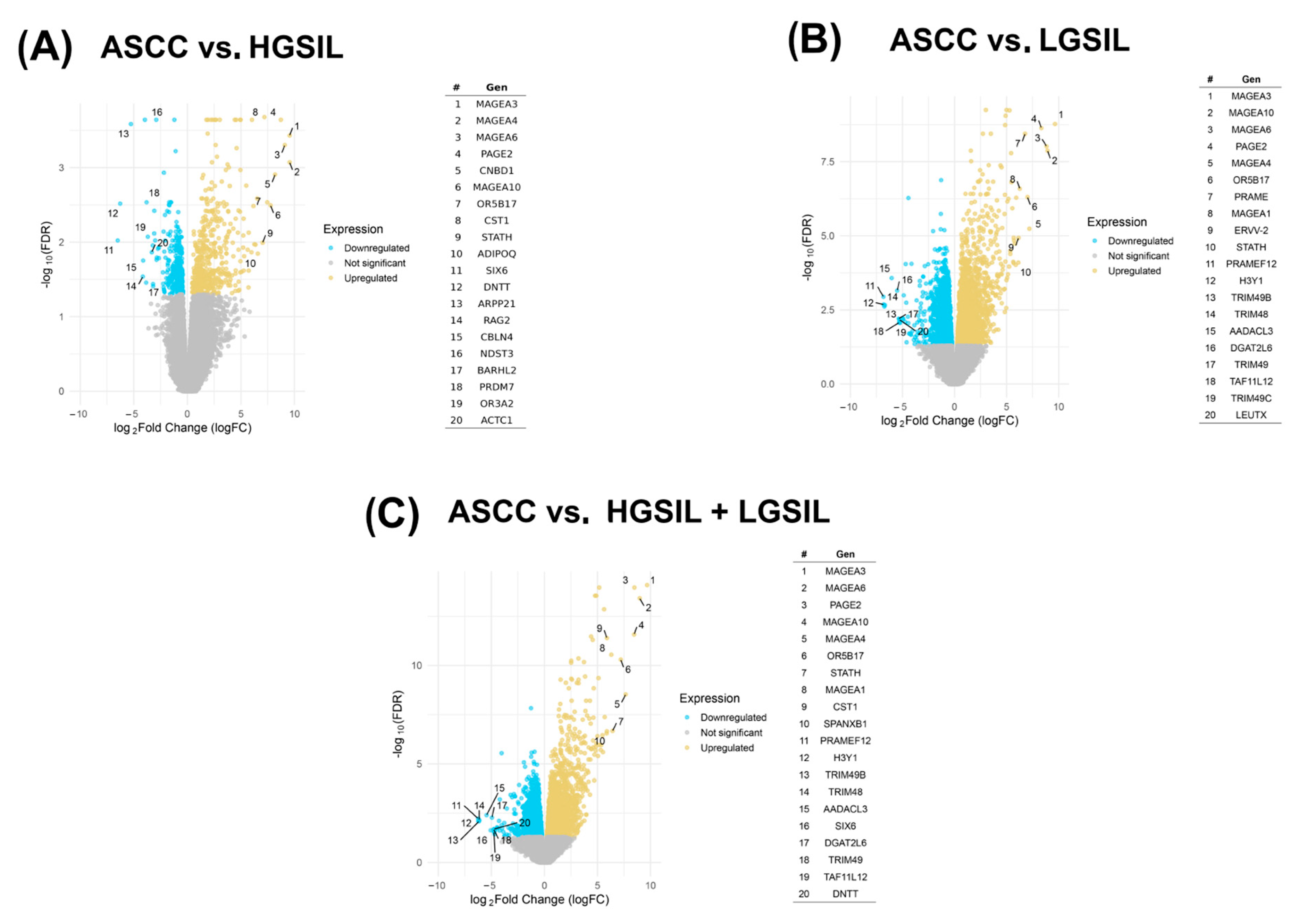
2. Results
2.1. Differential Microbial Abundance Across Precursor Lesions and ASCC
2.2. Detection and Profiling of Viral miRNAs
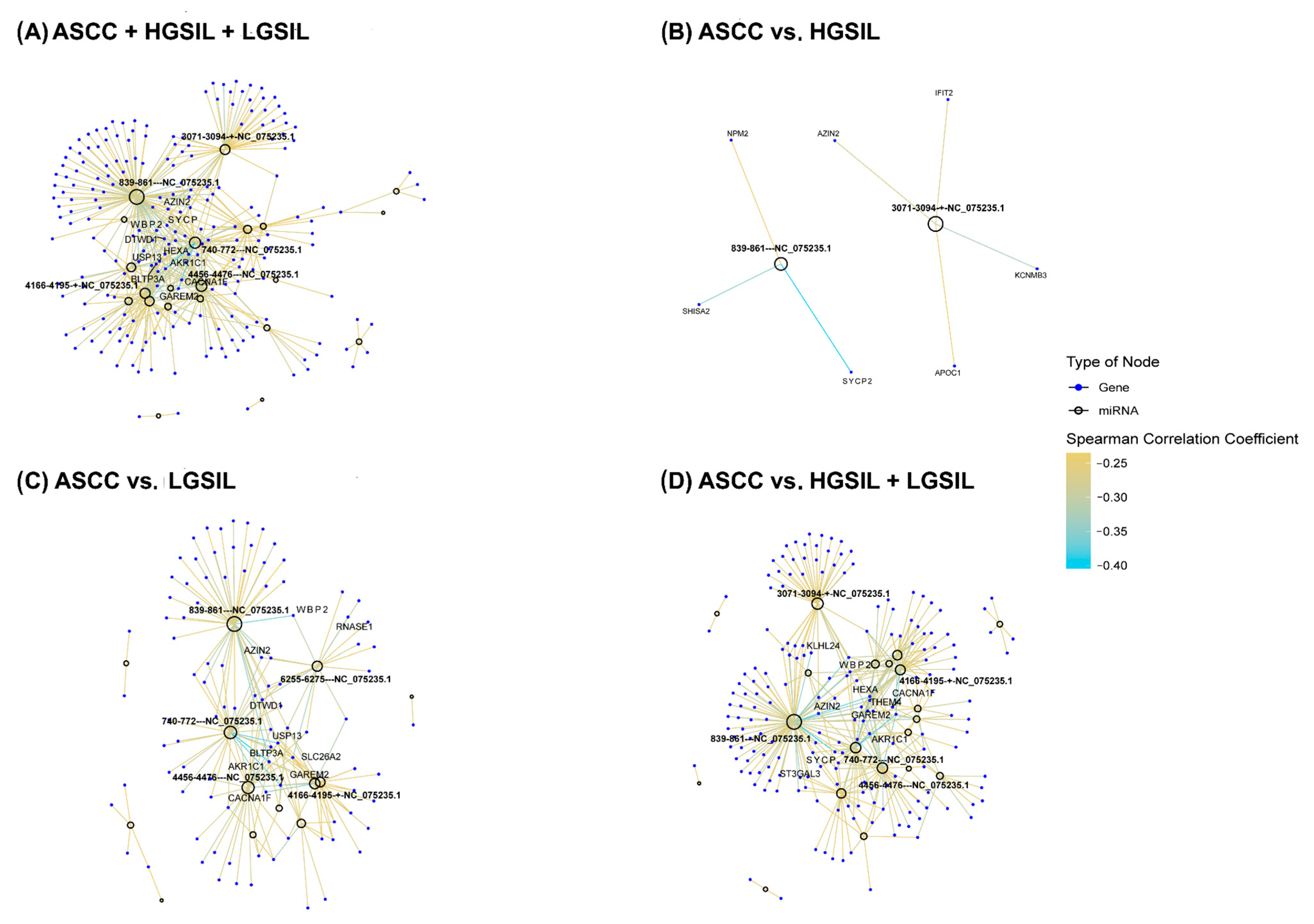
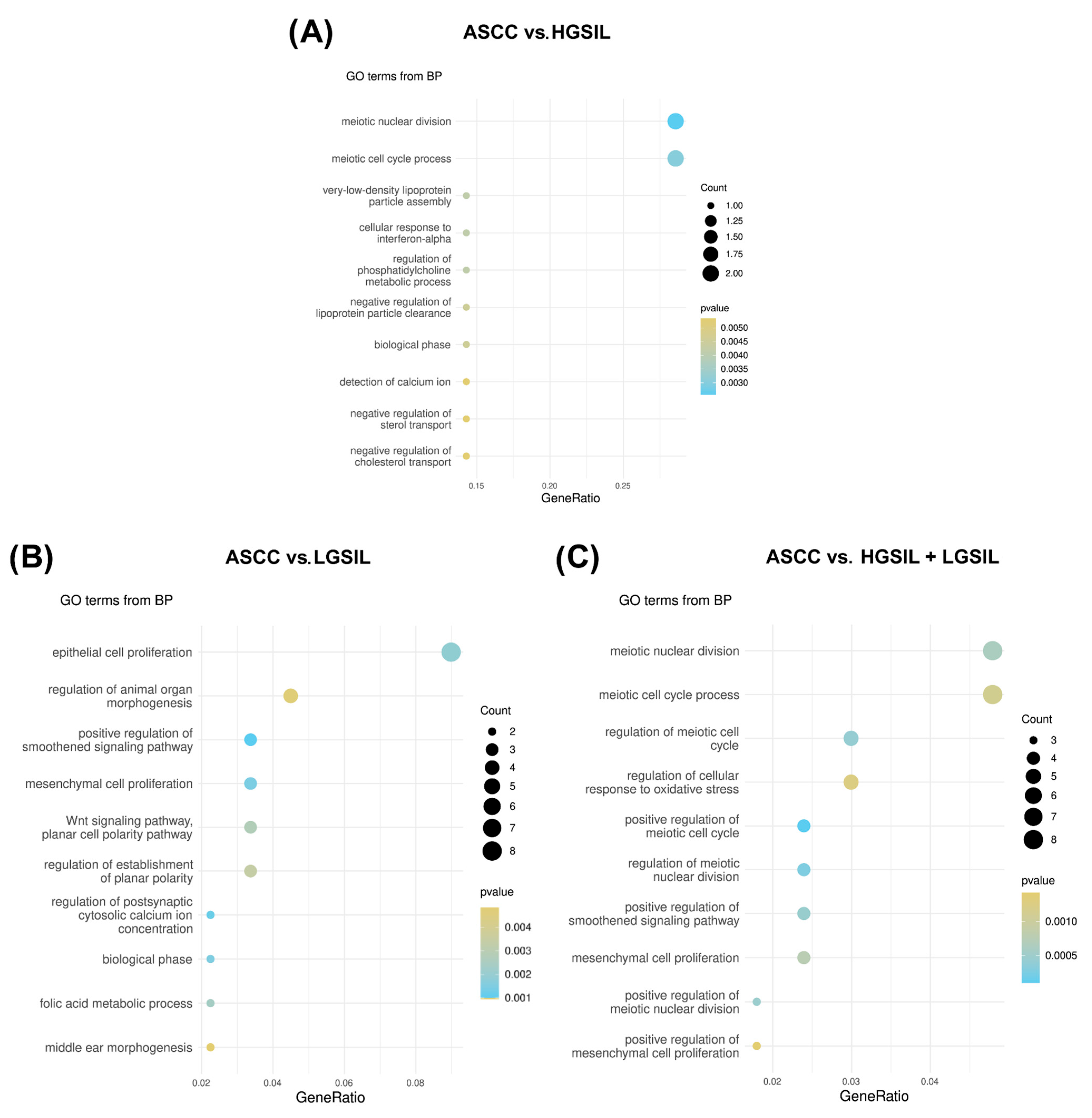
2.3. Prediction and Correlation of Viral miRNA Targets with Host Gene Expression
3. Discussion
4. Material and Methods
4.1. Data Acquisition and Comparative Design
4.2. Processing, Mapping, and Analysis of Human Transcriptomics Data
4.3. Analysis Workflow for Viral miRNA Identification
4.4. Prediction of miRNA-Regulated Target Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| Alpha-PV | Alphapapillomavirus |
| ASCC | Anal squamous cell carcinoma |
| HGSIL | High-grade squamous intraepithelial lesion |
| HPV | Human papillomavirus |
| LGSIL | Low-grade squamous intraepithelial lesion |
| miRNAs | MicroRNAs |
| ncRNAs | Non-coding RNAs |
| SIL | Squamous intraepithelial lesion |
| UTR | Untranslated region |
References
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.F.; Gregory, T.R. The case for junk DNA. PLoS Genet. 2014, 10, e1004351. [Google Scholar] [CrossRef]
- The expanding world of noncoding RNA biology. Nat. Cell Biol. 2022, 24, 1447.
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Nemeth, K.; Bayraktar, R.; Ferracin, M.; Calin, G.A. Non-coding RNAs in disease: From mechanisms to therapeutics. Nat. Rev. Genet. 2024, 25, 211–232. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Gao, F.; Wang, F.; Chen, Y.; Deng, B.; Yang, F.; Cao, H.; Chen, J.; Chen, H.; Qi, F.; Kapranov, P. The human genome encodes a multitude of novel miRNAs. Nucleic Acids Res. 2025, 53, gkaf070. [Google Scholar] [CrossRef]
- Drula, R.; Pardini, B.; Fu, X.; De Los Santos, M.C.; Jurj, A.; Pang, L.; El-Daly, S.M.; Fabris, L.; Knutsen, E.; Dragomir, M.P.; et al. 17beta-estradiol promotes extracellular vesicle release and selective miRNA loading in ERalpha-positive breast cancer. Proc. Natl. Acad. Sci. USA 2023, 120, e2122053120. [Google Scholar] [CrossRef]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.M.; Kruger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An unconventional role for miRNA: Let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat. Neurosci. 2012, 15, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, J.; Xiong, J.; Hu, J.Q.; Yang, L.Y.; Sun, Y.X.; Wei, Y.; Zhao, Y.; Li, X.; Zheng, Q.H.; et al. Potential cardiac-derived exosomal miRNAs involved in cardiac healing and remodeling after myocardial ischemia-reperfusion injury. Sci. Rep. 2024, 14, 24275. [Google Scholar] [CrossRef] [PubMed]
- Starhof, C.; Hejl, A.M.; Heegaard, N.H.H.; Carlsen, A.L.; Burton, M.; Lilje, B.; Winge, K. The biomarker potential of cell-free microRNA from cerebrospinal fluid in Parkinsonian Syndromes. Mov. Disord. 2019, 34, 246–254. [Google Scholar] [CrossRef]
- Negi, V.; Paul, D.; Das, S.; Bajpai, P.; Singh, S.; Mukhopadhyay, A.; Agrawal, A.; Ghosh, B. Altered expression and editing of miRNA-100 regulates iTreg differentiation. Nucleic Acids Res. 2015, 43, 8057–8065. [Google Scholar] [CrossRef] [PubMed]
- Alisi, L.; Giovannetti, F.; Armentano, M.; Lucchino, L.; Lambiase, A.; Bruscolini, A. Challenging corneal diseases and microRNA expression: Focus on rare diseases and new therapeutic frontiers. Surv. Ophthalmol. 2025, 70, 121–131. [Google Scholar] [CrossRef]
- Fehlmann, T.; Lehallier, B.; Schaum, N.; Hahn, O.; Kahraman, M.; Li, Y.; Grammes, N.; Geffers, L.; Backes, C.; Balling, R.; et al. Common diseases alter the physiological age-related blood microRNA profile. Nat. Commun. 2020, 11, 5958. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y.; et al. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Li, X.; Padi, S.K.; Zhang, Q.; Tang, M.S.; Guo, B. Downregulation of miR-205 and miR-31 confers resistance to chemotherapy-induced apoptosis in prostate cancer cells. Cell Death Dis. 2010, 1, e105. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Chi, Y.; Chen, Y.; Huang, S.; Ye, X.; Niu, J.; Wang, W.; Pfeffer, L.M.; Shao, Z.M.; Wu, Z.H.; et al. MiRNA-621 sensitizes breast cancer to chemotherapy by suppressing FBXO11 and enhancing p53 activity. Oncogene 2016, 35, 448–458. [Google Scholar] [CrossRef]
- Niu, J.; Xue, A.; Chi, Y.; Xue, J.; Wang, W.; Zhao, Z.; Fan, M.; Yang, C.H.; Shao, Z.M.; Pfeffer, L.M.; et al. Induction of miRNA-181a by genotoxic treatments promotes chemotherapeutic resistance and metastasis in breast cancer. Oncogene 2016, 35, 1302–1313. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Priya, S.; Burns, M.B.; Ward, T.; Mars, R.A.T.; Adamowicz, B.; Lock, E.F.; Kashyap, P.C.; Knights, D.; Blekhman, R. Identification of shared and disease-specific host gene-microbiome associations across human diseases using multi-omic integration. Nat. Microbiol. 2022, 7, 780–795. [Google Scholar] [CrossRef]
- Zhou, X.; Li, X.; Wu, M. miRNAs reshape immunity and inflammatory responses in bacterial infection. Signal Transduct. Target. Ther. 2018, 3, 14. [Google Scholar] [CrossRef]
- Liu, X.; Xiong, W.; Ye, M.; Lu, T.; Yuan, K.; Chang, S.; Han, Y.; Wang, Y.; Lu, L.; Bao, Y. Non-coding RNAs expression in SARS-CoV-2 infection: Pathogenesis, clinical significance, and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liang, Z.; Wang, C.; Shen, Z.; Sun, S.; Gong, C.; Hu, X. Viral Circular RNAs and Their Possible Roles in Virus-Host Interaction. Front. Immunol. 2022, 13, 939768. [Google Scholar] [CrossRef] [PubMed]
- Giza, D.E.; Fuentes-Mattei, E.; Bullock, M.D.; Tudor, S.; Goblirsch, M.J.; Fabbri, M.; Lupu, F.; Yeung, S.J.; Vasilescu, C.; Calin, G.A. Cellular and viral microRNAs in sepsis: Mechanisms of action and clinical applications. Cell Death Differ. 2016, 23, 1906–1918. [Google Scholar] [CrossRef]
- Abbas, M.N.; Kausar, S.; Asma, B.; Ran, W.; Li, J.; Lin, Z.; Li, T.; Cui, H. MicroRNAs reshape the immunity of insects in response to bacterial infection. Front. Immunol. 2023, 14, 1176966. [Google Scholar] [CrossRef]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- Thibault, P.A.; Huys, A.; Amador-Canizares, Y.; Gailius, J.E.; Pinel, D.E.; Wilson, J.A. Regulation of Hepatitis C Virus Genome Replication by Xrn1 and MicroRNA-122 Binding to Individual Sites in the 5’ Untranslated Region. J. Virol. 2015, 89, 6294–6311. [Google Scholar] [CrossRef]
- Luna, J.M.; Scheel, T.K.; Danino, T.; Shaw, K.S.; Mele, A.; Fak, J.J.; Nishiuchi, E.; Takacs, C.N.; Catanese, M.T.; de Jong, Y.P.; et al. Hepatitis C virus RNA functionally sequesters miR-122. Cell 2015, 160, 1099–1110. [Google Scholar] [CrossRef]
- Martin-Perez, D.; Vargiu, P.; Montes-Moreno, S.; Leon, E.A.; Rodriguez-Pinilla, S.M.; Lisio, L.D.; Martinez, N.; Rodriguez, R.; Mollejo, M.; Castellvi, J.; et al. Epstein-Barr virus microRNAs repress BCL6 expression in diffuse large B-cell lymphoma. Leukemia 2012, 26, 180–183. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Diggins, N.L.; Hancock, M.H. Viral miRNA regulation of host gene expression. Semin. Cell Dev. Biol. 2023, 146, 2–19. [Google Scholar] [CrossRef]
- Morrison, K.; Manzano, M.; Chung, K.; Schipma, M.J.; Bartom, E.T.; Gottwein, E. The Oncogenic Kaposi’s Sarcoma-Associated Herpesvirus Encodes a Mimic of the Tumor-Suppressive miR-15/16 miRNA Family. Cell Rep. 2019, 29, 2961–2969.e6. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, J.; Xiang, X.; Deng, J.; Gao, D.; Sheng, X. Viral long non-coding RNA regulates virus life-cycle and pathogenicity. Mol. Biol. Rep. 2022, 49, 6693–6700. [Google Scholar] [CrossRef]
- Borah, S.; Darricarrere, N.; Darnell, A.; Myoung, J.; Steitz, J.A. A viral nuclear noncoding RNA binds re-localized poly(A) binding protein and is required for late KSHV gene expression. PLoS Pathog. 2011, 7, e1002300. [Google Scholar] [CrossRef]
- Rossetto, C.C.; Pari, G.S. Kaposi’s sarcoma-associated herpesvirus noncoding polyadenylated nuclear RNA interacts with virus- and host cell-encoded proteins and suppresses expression of genes involved in immune modulation. J. Virol. 2011, 85, 13290–13297. [Google Scholar] [CrossRef]
- Kang, S.H.; Aknadibossian, V.; Kharel, L.; Mudiyanselage, S.D.D.; Wang, Y.; Folimonova, S.Y. The Intriguing Conundrum of a Nonconserved Multifunctional Protein of Citrus Tristeza Virus That Interacts with a Viral Long Non-Coding RNA. Viruses 2021, 13, 2129. [Google Scholar] [CrossRef] [PubMed]
- Marquitz, A.R.; Mathur, A.; Edwards, R.H.; Raab-Traub, N. Host Gene Expression Is Regulated by Two Types of Noncoding RNAs Transcribed from the Epstein-Barr Virus BamHI A Rightward Transcript Region. J. Virol. 2015, 89, 11256–11268. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Tripathy, M.K.; Pasquereau, S.; Al Moussawi, F.; Abbas, W.; Coquard, L.; Khan, K.A.; Russo, L.; Algros, M.P.; Valmary-Degano, S.; et al. The Human Cytomegalovirus Strain DB Activates Oncogenic Pathways in Mammary Epithelial Cells. EBioMedicine 2018, 30, 167–183. [Google Scholar] [CrossRef]
- Gorbea, C.; Mosbruger, T.; Cazalla, D. A viral Sm-class RNA base-pairs with mRNAs and recruits microRNAs to inhibit apoptosis. Nature 2017, 550, 275–279. [Google Scholar] [CrossRef]
- Marima, R.; Hull, R.; Mbeje, M.; Molefi, T.; Mathabe, K.; Elbagory, A.M.; Demetriou, D.; Dlamini, Z. Role of Precision Oncology in Type II Endometrial and Prostate Cancers in the African Population: Global Cancer Genomics Disparities. Int. J. Mol. Sci. 2022, 23, 628. [Google Scholar] [CrossRef] [PubMed]
- Hoots, B.E.; Palefsky, J.M.; Pimenta, J.M.; Smith, J.S. Human papillomavirus type distribution in anal cancer and anal intraepithelial lesions. Int. J. Cancer 2009, 124, 2375–2383. [Google Scholar] [CrossRef]
- Islami, F.; Ferlay, J.; Lortet-Tieulent, J.; Bray, F.; Jemal, A. International trends in anal cancer incidence rates. Int. J. Epidemiol. 2017, 46, 924–938. [Google Scholar] [CrossRef]
- Clifford, G.M.; Georges, D.; Shiels, M.S.; Engels, E.A.; Albuquerque, A.; Poynten, I.M.; de Pokomandy, A.; Easson, A.M.; Stier, E.A. A meta-analysis of anal cancer incidence by risk group: Toward a unified anal cancer risk scale. Int. J. Cancer 2021, 148, 38–47. [Google Scholar] [CrossRef]
- Daling, J.R.; Madeleine, M.M.; Johnson, L.G.; Schwartz, S.M.; Shera, K.A.; Wurscher, M.A.; Carter, J.J.; Porter, P.L.; Galloway, D.A.; McDougall, J.K. Human papillomavirus, smoking, and sexual practices in the etiology of anal cancer. Cancer 2004, 101, 270–280. [Google Scholar] [CrossRef]
- Pineda, C.E.; Welton, M.L. Management of anal squamous intraepithelial lesions. Clin. Colon Rectal Surg. 2009, 22, 94–101. [Google Scholar] [CrossRef]
- Cornall, A.M.; Roberts, J.M.; Garland, S.M.; Hillman, R.J.; Grulich, A.E.; Tabrizi, S.N. Anal and perianal squamous carcinomas and high-grade intraepithelial lesions exclusively associated with “low-risk” HPV genotypes 6 and 11. Int. J. Cancer 2013, 133, 2253–2258. [Google Scholar] [CrossRef]
- Liu, M.Z.; Hung, Y.P.; Huang, E.C.; Howitt, B.E.; Nucci, M.R.; Crum, C.P. HPV 6-associated HSIL/Squamous Carcinoma in the Anogenital Tract. Int. J. Gynecol. Pathol. 2019, 38, 493–497. [Google Scholar] [CrossRef]
- Vyas, M.; Gonzalez, R.S. Anal intraepithelial neoplasia: A review of terminology, differential diagnoses, and patient management. Hum. Pathol. 2023, 132, 56–64. [Google Scholar] [CrossRef]
- Madahian, S.; Judelson, R.; Zhu, X.; Meng, X.; Dresser, K.; Hutchinson, L.; Bledsoe, J.R. CD56 expression in basaloid anal squamous cell carcinoma—A potential diagnostic pitfall. Ann. Diagn. Pathol. 2021, 53, 151758. [Google Scholar] [CrossRef]
- Zhu, X.; Jamshed, S.; Zou, J.; Azar, A.; Meng, X.; Bathini, V.; Dresser, K.; Strock, C.; Yalamarti, B.; Yang, M.; et al. Molecular and immunophenotypic characterization of anal squamous cell carcinoma reveals distinct clinicopathologic groups associated with HPV and TP53 mutation status. Mod. Pathol. 2021, 34, 1017–1030. [Google Scholar] [CrossRef]
- Sun, G.; Dong, X.; Tang, X.; Qu, H.; Zhang, H.; Zhao, E. The prognostic value of HPV combined p16 status in patients with anal squamous cell carcinoma: A meta-analysis. Oncotarget 2018, 9, 8081–8088. [Google Scholar] [CrossRef]
- Silverberg, M.J.; Lau, B.; Justice, A.C.; Engels, E.; Gill, M.J.; Goedert, J.J.; Kirk, G.D.; D’Souza, G.; Bosch, R.J.; Brooks, J.T.; et al. Risk of anal cancer in HIV-infected and HIV-uninfected individuals in North America. Clin. Infect. Dis. 2012, 54, 1026–1034. [Google Scholar] [CrossRef]
- Sunesen, K.G.; Norgaard, M.; Thorlacius-Ussing, O.; Laurberg, S. Immunosuppressive disorders and risk of anal squamous cell carcinoma: A nationwide cohort study in Denmark, 1978-2005. Int. J. Cancer 2010, 127, 675–684. [Google Scholar] [CrossRef]
- Ron, R.; Cabello, A.; Gosalbes, M.J.; Sanchez-Conde, M.; Talavera-Rodriguez, A.; Zamora, J.; Monge-Maillo, B.; Jimenez, D.; Martinez-Sanz, J.; Lopez, Y.; et al. Exploiting the Microbiota for the Diagnosis of Anal Precancerous Lesions in Men Who Have Sex With Men. J. Infect. Dis. 2021, 224, 1247–1256. [Google Scholar] [CrossRef]
- Serrano-Villar, S.; Tincati, C.; Raju, S.C.; Saenz, J.S.; Moreno, E.; Bargiela, R.; Cabello-Ubeda, A.; Sendagorta, E.; Kurz, A.; Perez Molina, J.A.; et al. Microbiome-derived cobalamin and succinyl-CoA as biomarkers for improved screening of anal cancer. Nat. Med. 2023, 29, 1738–1749. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Liu, Y.; He, Y.; Duan, X.; Li, Q.; Huang, Y.; Xu, G.; Lu, Q. HPV16-miRNAs exert oncogenic effects through enhancers in human cervical cancer. Cancer Cell Int. 2024, 24, 172. [Google Scholar] [CrossRef]
- Lacunza, E.; Fink, V.; Salas, M.E.; Gun, A.M.; Basiletti, J.A.; Picconi, M.A.; Golubicki, M.; Robbio, J.; Kujaruk, M.; Iseas, S.; et al. Transcriptome and microbiome-immune changes across preinvasive and invasive anal cancer lesions. JCI Insight 2024, 9, e180907. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; McVeigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Zhang, Y.; Qiu, K.; Ren, J.; Zhao, Y.; Cheng, P. Roles of human papillomavirus in cancers: Oncogenic mechanisms and clinical use. Signal Transduct. Target. Ther. 2025, 10, 44. [Google Scholar] [CrossRef]
- Burd, E.M. Human papillomavirus and cervical cancer. Clin. Microbiol. Rev. 2003, 16, 1–17. [Google Scholar] [CrossRef]
- Grundhoff, A.; Sullivan, C.S. Virus-encoded microRNAs. Virology 2011, 411, 325–343. [Google Scholar] [CrossRef]
- Hull, R.; Marima, R.; Alaouna, M.; Demetriou, D.; Reis, R.M.; Molefi, T.; Dlamini, Z. Viral Encoded miRNAs in Tumorigenesis: Theranostic Opportunities in Precision Oncology. Microorganisms 2022, 10, 1448. [Google Scholar] [CrossRef]
- Quiroga-Garza, G.; Zhou, H.; Mody, D.R.; Schwartz, M.R.; Ge, Y. Unexpected high prevalence of HPV 90 infection in an underserved population: Is it really a low-risk genotype? Arch. Pathol. Lab. Med. 2013, 137, 1569–1573. [Google Scholar] [CrossRef]
- Liang, L.A.; Tanaka, L.F.; Radde, K.; Bussas, U.; Ikenberg, H.; Heideman, D.A.M.; Meijer, C.; Blettner, M.; Klug, S.J. Population-based age- and type-specific prevalence of human papillomavirus among non-vaccinated women aged 30 years and above in Germany. BMC Infect. Dis. 2024, 24, 1008. [Google Scholar] [CrossRef]
- Silva, L.L.D.; Teles, A.M.; Santos, J.M.O.; Souza de Andrade, M.; Medeiros, R.; Faustino-Rocha, A.I.; Oliveira, P.A.; Dos Santos, A.P.A.; Ferreira Lopes, F.; Braz, G.; et al. Malignancy Associated with Low-Risk HPV6 and HPV11: A Systematic Review and Implications for Cancer Prevention. Cancers 2023, 15, 4068. [Google Scholar] [CrossRef]
- Garbuglia, A.R.; Gentile, M.; Del Nonno, F.; Lorenzini, P.; Lapa, D.; Lupi, F.; Pinnetti, C.; Baiocchini, A.; Libertone, R.; Cicalini, S.; et al. An anal cancer screening program for MSM in Italy: Prevalence of multiple HPV types and vaccine-targeted infections. J. Clin. Virol. 2015, 72, 49–54. [Google Scholar] [CrossRef]
- West, A.M.; Rosenberg, S.C.; Ur, S.N.; Lehmer, M.K.; Ye, Q.; Hagemann, G.; Caballero, I.; Uson, I.; MacQueen, A.J.; Herzog, F.; et al. A conserved filamentous assembly underlies the structure of the meiotic chromosome axis. Elife 2019, 8, e40372. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, B.; Zhang, L.; Wang, X.; Zhu, X.; Yang, H.; Zhang, F.; Zhu, X.; Zhou, B.; Yao, S.; et al. Meiotic protein SYCP2 confers resistance to DNA-damaging agents through R-loop-mediated DNA repair. Nat. Commun. 2024, 15, 1568. [Google Scholar] [CrossRef]
- Zhou, J.; Stein, P.; Leu, N.A.; Chmatal, L.; Xue, J.; Ma, J.; Huang, X.; Lampson, M.A.; Schultz, R.M.; Wang, P.J. Accelerated reproductive aging in females lacking a novel centromere protein SYCP2L. Hum. Mol. Genet. 2015, 24, 6505–6514. [Google Scholar] [CrossRef]
- He, W.B.; Tan, C.; Zhang, Y.X.; Meng, L.L.; Gong, F.; Lu, G.X.; Lin, G.; Du, J.; Tan, Y.Q. Homozygous variants in SYCP2L cause premature ovarian insufficiency. J. Med. Genet. 2021, 58, 168–172. [Google Scholar] [CrossRef]
- Ben, W.; Yang, Y.; Yuan, J.; Sun, J.; Huang, M.; Zhang, D.; Zheng, J. Human papillomavirus 16 E6 modulates the expression of host microRNAs in cervical cancer. Taiwan. J. Obstet. Gynecol. 2015, 54, 364–370. [Google Scholar] [CrossRef]
- Shakib, K.; Norman, J.T.; Fine, L.G.; Brown, L.R.; Godovac-Zimmermann, J. Proteomics profiling of nuclear proteins for kidney fibroblasts suggests hypoxia, meiosis, and cancer may meet in the nucleus. Proteomics 2005, 5, 2819–2838. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Pan, C.; Wang, W.; Yu, Y. HPV-driven heterogeneity in cervical cancer: Study on the role of epithelial cells and myofibroblasts in the tumor progression based on single-cell RNA sequencing analysis. PeerJ 2024, 12, e18158. [Google Scholar] [CrossRef]
- Nikmanesh, N.; Safarpour, A.R.; Niknam, R.; Nikmanesh, Y. Short Communication on HPV Infection, Pathogenesis of Cancer, Immune Cells Involved in Infection, Vaccination, and Suggested Treatments. J. Cancer Immunol. 2025, 7, 72–80. [Google Scholar] [CrossRef]
- Yeung, K.T.; Yang, J. Epithelial-mesenchymal transition in tumor metastasis. Mol. Oncol. 2017, 11, 28–39. [Google Scholar] [CrossRef]
- Han, Y.; Li, Q.; Ling, C.; Jin, M.; Li, D.; Zhong, J.; Wang, L. HPV-Induced MiR-21 Promotes Epithelial Mesenchymal Transformation and Tumor Progression in Cervical Cancer Cells through the TGFbeta R2/hTERC Pathway. Contrast Media Mol. Imaging 2022, 2022, 6297694. [Google Scholar] [CrossRef] [PubMed]
- Bello, J.O.; Nieva, L.O.; Paredes, A.C.; Gonzalez, A.M.; Zavaleta, L.R.; Lizano, M. Regulation of the Wnt/beta-Catenin Signaling Pathway by Human Papillomavirus E6 and E7 Oncoproteins. Viruses 2015, 7, 4734–4755. [Google Scholar] [CrossRef]
- Kolluri, A.; Ho, M. The Role of Glypican-3 in Regulating Wnt, YAP, and Hedgehog in Liver Cancer. Front. Oncol. 2019, 9, 708. [Google Scholar] [CrossRef]
- Sun, B.; Huang, Z.; Wang, B.; Yu, Y.; Lin, S.; Luo, L.; Wang, Y.; Huang, Z. Significance of Glypican-3 (GPC3) Expression in Hepatocellular Cancer Diagnosis. Med. Sci. Monit. 2017, 23, 850–855. [Google Scholar] [CrossRef]
- Wang, R.; Lin, X. GPC2 as a diagnostic and prognostic marker regulated progression of colorectal cancer. Arab. J. Gastroenterol. 2024, 25, 51–57. [Google Scholar] [CrossRef]
- Hosoya, N.; Miyagawa, K. Synaptonemal complex proteins modulate the level of genome integrity in cancers. Cancer Sci. 2021, 112, 989–996. [Google Scholar] [CrossRef] [PubMed]
- De Marco, F. Oxidative stress and HPV carcinogenesis. Viruses 2013, 5, 708–731. [Google Scholar] [CrossRef]
- Henderson, S.; Chakravarthy, A.; Su, X.; Boshoff, C.; Fenton, T.R. APOBEC-mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep. 2014, 7, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.J.; Westrich, J.A.; Doorslaer, K.V.; Pyeon, D. Roles of APOBEC3A and APOBEC3B in Human Papillomavirus Infection and Disease Progression. Viruses 2017, 9, 233. [Google Scholar] [CrossRef]
- Peronace, C.; Cione, E.; Abrego-Guandique, D.M.; Fazio, M.; Panduri, G.; Caroleo, M.C.; Cannataro, R.; Minchella, P. FAM19A4 and hsa-miR124-2 Double Methylation as Screening for ASC-H- and CIN1 HPV-Positive Women. Pathogens 2024, 13, 312. [Google Scholar] [CrossRef]
- Chen, H.; Cao, D.; Han, N.; Zhang, M.; Jiang, W.; Wang, X.; Zeng, Q.; Tang, H. Hepatitis B Virus-Encoded MicroRNA (HBV-miR-3) Inhibits FIH-1 Expression to Promote Tumor Angiogenesis in HBV-Related Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2023, 10, 2337–2353. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.L.; Terrón-Camero, L.C.; Castillo-González, J.; Fernández-Rengel, I.; Delgado, M.; Gonzalez-Rey, E.; Andrés-León, E. reanalyzerGSE: Tackling the everlasting lack of reproducibility and reanalyses in transcriptomics. bioRxiv 2023. bioRxiv:12.548663. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Nikolayeva, O.; Robinson, M.D. edgeR for differential RNA-seq and ChIP-seq analysis: An application to stem cell biology. Methods Mol. Biol. 2014, 1150, 45–79. [Google Scholar]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef]
- Cai, Z.; Fu, P.; Qiu, Y.; Wu, A.; Zhang, G.; Wang, Y.; Jiang, T.; Ge, X.Y.; Zhu, H.; Peng, Y. vsRNAfinder: A novel method for identifying high-confidence viral small RNAs from small RNA-Seq data. Brief. Bioinform. 2022, 23, bbac496. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef] [PubMed]
- Chipman, L.B.; Pasquinelli, A.E. miRNA Targeting: Growing beyond the Seed. Trends Genet. 2019, 35, 215–222. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García, D.J.; Pulpillo-Berrocal, M.A.; Ruiz, J.L.; Andrés-León, E.; Terrón-Camero, L.C. Human Papillomavirus-Encoded microRNAs as Regulators of Human Gene Expression in Anal Squamous Cell Carcinoma: A Meta-Transcriptomics Study. Non-Coding RNA 2025, 11, 43. https://doi.org/10.3390/ncrna11030043
García DJ, Pulpillo-Berrocal MA, Ruiz JL, Andrés-León E, Terrón-Camero LC. Human Papillomavirus-Encoded microRNAs as Regulators of Human Gene Expression in Anal Squamous Cell Carcinoma: A Meta-Transcriptomics Study. Non-Coding RNA. 2025; 11(3):43. https://doi.org/10.3390/ncrna11030043
Chicago/Turabian StyleGarcía, Daniel J., Marco A. Pulpillo-Berrocal, José L. Ruiz, Eduardo Andrés-León, and Laura C. Terrón-Camero. 2025. "Human Papillomavirus-Encoded microRNAs as Regulators of Human Gene Expression in Anal Squamous Cell Carcinoma: A Meta-Transcriptomics Study" Non-Coding RNA 11, no. 3: 43. https://doi.org/10.3390/ncrna11030043
APA StyleGarcía, D. J., Pulpillo-Berrocal, M. A., Ruiz, J. L., Andrés-León, E., & Terrón-Camero, L. C. (2025). Human Papillomavirus-Encoded microRNAs as Regulators of Human Gene Expression in Anal Squamous Cell Carcinoma: A Meta-Transcriptomics Study. Non-Coding RNA, 11(3), 43. https://doi.org/10.3390/ncrna11030043