1. Introduction
Single cell migration is an almost ubiquitous phenomenon in eukaryotic biology that serves many important physiological roles including embryonic development, immune response, and wound healing [
1]. Cells use a variety of biophysical mechanisms to migrate that vary depending on their external environment [
2]. For example, mesenchymal migration employed by cells such as keratocytes and fibroblasts on a flat 2D substrate is characterized by actin polymerization at the leading edge and substrate adhesion [
3,
4,
5]. In contrast, some cells in 3D use an amoeboid mode of motility, where cells have a round morphology and lack mature adhesions and actin stress fibers [
6,
7]. Amoeboid migration plays important roles in developmental biology and immune system function [
8,
9]. Additionally, certain tumor cells can transition between mesenchymal and amoeboid migration modes and thereby increase cancer invasiveness [
1]. Some cells have even been shown to switch migration modes after biochemical or mechanical stimulation [
10,
11,
12].
The focus of this manuscript is on a relatively novel migratory mechanism of rounded cells that do not rely on cell-surface adhesion for efficient migration in complex 3D environments [
13,
14]. We refer to this form of locomotion as adhesion-independent amoeboid movement. This migration mode is relevant for the migration of leukocytes through the endothelial barrier out of blood circulation to the location of damaged tissue during wound healing [
8,
15], and the in vivo migration of various cells in developing embryos [
16], metastatic cancer [
1,
17], and immune cells migrating through tissue while patrolling for pathogens [
18].
Amoeboid cells move much faster and are more autonomous from their extracellular environment than mesenchymal cells in the sense that they adapt to their environment instead of remodeling it [
13]. Locomotion of amoeboid cells does not depend on adhesive ligands, and they are able to migrate efficiently in a non-frictional manner even in suspensions or artificial materials [
15,
19]. In the adhesion-independent amoeboid mechanism, cells find the path of least resistance through the extracellular matrix by picking larger pores in the matrix over small ones [
20], actively deforming their cell body and/or transiently dilating the pore in order to pass it through [
21,
22,
23]. Charras and Paluch hypothesized that a cell exerts forces perpendicularly to the substrate such that it can squeeze itself forward using membrane protrusions called
blebs [
24]. This phenomenon has been termed ‘chimneying’ [
25] (authors observed cells migrating between two glass coverslips), in reference to a technique used by mountain climbers. Exactly how cellular forces are transmitted to the substrate in order to produce migration remains an open question.
In this work, we consider amoeboid cell migration in the case where the leading edge protrusion is generated by a bleb, a spherical membrane protrusion characterized by a delamination of the actomyosin cortex from the cell plasma membrane [
26]. When the actin cortex is separated from the membrane, tension from actomyosin contractility is no longer transmitted to the membrane in the delamination region, and pressure is locally reduced [
27]. Cytoplasmic content then streams from the cell body into this region, expanding the round membrane protrusion. Bleb initation can occur by either a localized loss in membrane-cortex adhesion proteins or local-rupture of the cortex [
24,
28]. Actin, myosin, and associated proteins eventually reform under the naked membrane to drive bleb retraction and complete the life-cycle of a bleb. Blebbing cell migration has been observed in a number of cell types, such as amoebas, zebrafish germ layer progenitor cells, and Walker 256 carcinosarcoma cells [
16,
29,
30].
Several computational models have been developed to investigate the relative importance of key factors in confined migration, such as geometry of the environment, actomyosin contractility, role of nucleus, and type of leading edge protrusion [
31,
32,
33]. The aim of such computational modeling is to complement the experimental findings and propose mechanisms for generating internal forces and transmitting these forces to the surface in order to produce traction. A detailed hybrid agent-based/finite-element model of cancer cell motility was presented in [
32]. The authors consider a number of migratory mechanisms including pressure-driven (amoeboid) and actin-rich (mesenchymal) protrusions on flat surfaces, channels, and discontinuous 3D-like environments with varying levels of cell-surface adhesion. The authors found that in the absence of any cell-surface adhesions only cells exhibiting the amoeboid mechanism could migrate efficiently in a discontinuous environment. A similar type of model was proposed in [
33] to quantify conditions for motility modes for a cell migrating through an elastic extracellular matrix. One limitation of both models is that intracellular fluid flow is not incorporated (i.e., intracellular pressure is treated as constant). The model in [
31] does include intra-and extracellular fluid mechanics, but results focus on the role of nuclear stiffness during amoeboid cell migration. The theory of active gels has also been used to model confined cell migration [
34,
35]. For example, in [
35], the authors show that motion can occur in confinement when the cell cytoplasm is modeled as a polymerizing viscoelastic material.
A natural question to ask is whether a confined environment is even necessary for cell migration in a fluid environment. Several groups have developed models to determine conditions when a cell can “swim” in low Reynolds number fluid. The Scallop Theorem states that time-symmetric motion cannot achieve net displacement in Stokes flow [
36]. In order to swim in fluid, many prokaryotes (and sperm, paramecia and some eukaryotes) use a beating flagella or cilia to migrate. Such flagellated organisms achieve self-propulsion through periodic flagellar bending waves [
37,
38]. Experiments have shown that amoebae and neutrophils are also able to swim [
39], and several models of amoeboid cell swimming have been developed (reviewed in [
40]). A model for bleb-based swimming modeled the cell as two spheres submerged in fluid that can expand or contract radially that are connected by an extensible arm in [
41]. An amoeboid cell representative of
Dictyostelium discoideum immersed in fluid was shown to swim through shape changes and membrane tension gradients in [
42,
43].
In [
44], the authors developed a model similar to ours in that it includes a blebbing cell immersed in Stokes fluid with the model equations solved for using the method of regularized Stokeslets [
45]. The authors showed that in their model the cell was able to swim because membrane deformations during bleb expansion differed from those during bleb retraction. Results showed that migration speed was optimal at an intermediate confinement level. This model was then used to predict that the optimal gap size increases with weakening adhesion between the cell membrane and actin cortex, which was experimentally verified in [
46].
Here, we present a dynamic computational model of adhesion-independent cell migration using cycles of bleb expansion and retraction. Our model is formulated using the method of regularized Stokeslets [
45] to handle the fluid-structure interaction. Our model differs from previous work in that we consider intra and extra-cellular fluid flows, the actomyosin cortex is modeled as a poro-viscoelastic material, and various channel geometries are considered. Our results show that cyclic pressure gradients from blebbing together with cortical actin dynamics result in cell shape changes such as expansion and contraction of the cell body. However, the shape change pattern is nearly reversible and does not result in sustained net locomotion, even in confined environments. In exploring design principles for locomotion, the channel width is varied and an asymmetrical wall geometry is considered. Neither one of these endeavours improved cell movement. However, introducing actin turnover did produce sustained net locomotion in both suspensions and confined environments. Our results show that confinement enhances locomotion speed for most channel geometries. Speficially, if the wall has large crevices, a cell can become stuck and bleb vertically within a channel gap. Simulation results also show that migration speed increases with actin turnover.
The paper is organized as follows. In
Section 2, we describe the model of the cell, channel, and bleb life cycle. We describe the computational algorithm to solve and implement the model equations. In
Section 3, the model is first simulated using a poroelastic cortex using different bleb sizes as well as channel geometries. Next, we consider blebbing with cortical actin turnover as modeled by the Maxwell viscoelastic constitutive law in unconfined and various confined geometries. The effect of actin turnover on cell migration speed is also quantified. A discussion of results and conclusion remarks are provided in
Section 4.
2. Materials and Methods
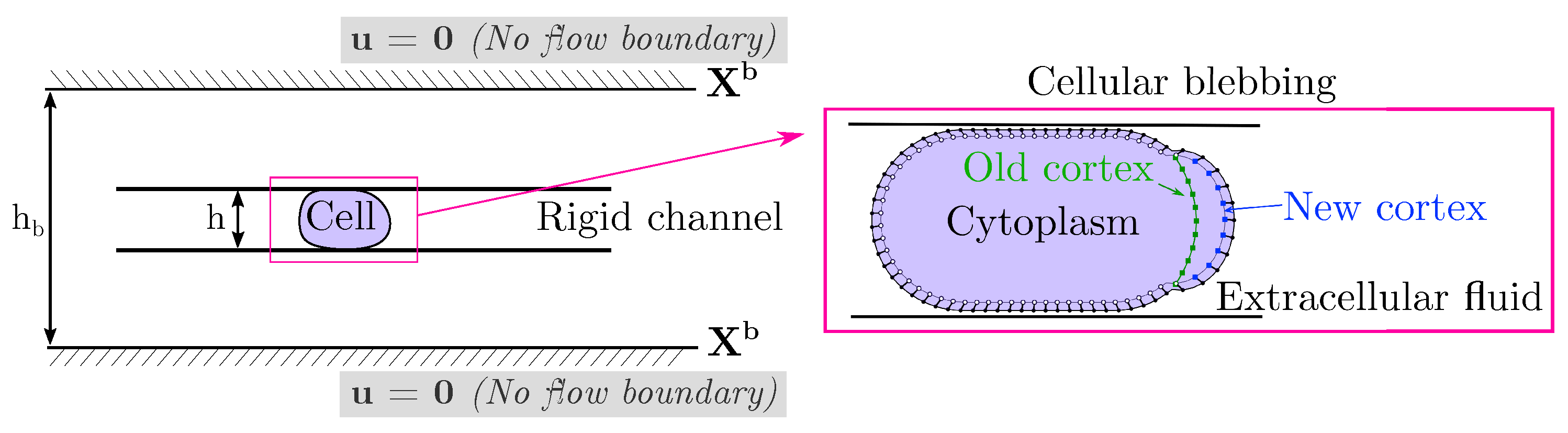
We build a computational model of a cell placed in a microfluidic rigid channel undergoing cycles of bleb expansion and contraction driven by intracellular fluid flows as illustrated in
Figure 1. The model is two-dimensional in that it captures the motion of the cell in the horizontal direction of motion as well as the channel height. We assume that the flow in the third dimension across the channel is negligible. The motion in the horizontal direction is due to extracellular fluid flows induced by cell shape changes. Our model has three sub-cellular components: an elastic plasma membrane, a contractile actomyosin cortex, and the cell cytoplasm. The cell cytoplasm is assumed to be a viscous fluid enclosed in the thin actin cortex and plasma membrane. The cortex is modeled as a thin 1D porous (visco)elastic material and its position is denoted by
where
t is time and
s is the local parametric coordinate on the structure. The plasma membrane is described as an incompressible 1D elastic outer layer with position
. The cortex is bound to the plasma membrane via membrane-anchoring proteins modeled here as elastic links [
47]. When a bleb is initiated, membrane-cortex attachment links are removed and the cell front expands due to the emergent fluid pressure gradient. As the cortex reforms at the bleb site, membrane-cortex attachment links reform and the cell front retracts. Since the focus is on adhesion-independent migration, there are no physical links between the cell and the extracellular environment.
2.1. Equations of Motion
Movement in viscous fluid at zero Reynolds number is governed by Stokes equations due to the small length scales at the cellular level [
36]. In our model, the external forces applied to the fluid are due to the deformations of the plasma membrane, the membrane-cortex attachment links, the viscous drag with cortex, as well as a repulsive steric interaction with the top and bottom channel walls:
where
is the fluid velocity,
p is the fluid pressure, and
is the fluid viscosity. Expressions for these cellular forces are provided below. We use the convention of lower case letters indicating fluid quantities while upper case letters indicate forces and positions of structures.
We consider two rheological descriptions for the membrane and cortex contours. Each contour experiences forces due to either elasticity or viscoelasticity. Let
denotes the elastic force density on the membrane and cortex
where
denotes the unit tangent vector to the closed curve
or
. The tension
T is given by
which describes a linearly elastic spring with stiffness
k and resting tension
. The plasma membrane and actomyosin cortex have their own characteristic stiffness and resting tension (see
Table 1). When the membrane and cortex are modeled as viscoelastic structures, we use a Maxwell model to capture stress relaxation of the actomyosin cortex due to actin filament rearrangement within the cortex [
48]. Following the approach in [
49], a viscoelastic structure is modeled as a purely elastic spring (see Equations (
3) and (
4)) whose reference configuration
relaxes to the current configuration
over time with the derived expression
Here,
is the deformation gradient tensor (
) and
is the strain relaxation timescale. In the limit of small strain, the authors in [
49] show that the update equation for the reference configuration in Equation (
5) together with the elastic force in Equation (
3) agrees with the Maxwell model for viscoelasticity.
Membrane-cortex attachments are modeled as elastic springs that connect the plasma membrane to the underlying actin cortex with a force density given by
The drag force on the cell cortex is balanced by (visco)-elastic forces within the cortex and elastic forces from cortex attachment to the plasma membrane:
The cortical drag is defined as where denotes the viscous drag coefficient and is the cortex velocity.
The cell interacts with the channel walls through a repulsive force due to contact with the surface:
Note that the steric force is only nonzero when the membrane location exceeds the top or bottom channel walls. Here, is the vertical distance from the plasma membrane to the channel walls, is a unit vector in the outward normal direction, and is the stiffness of the steric interaction.
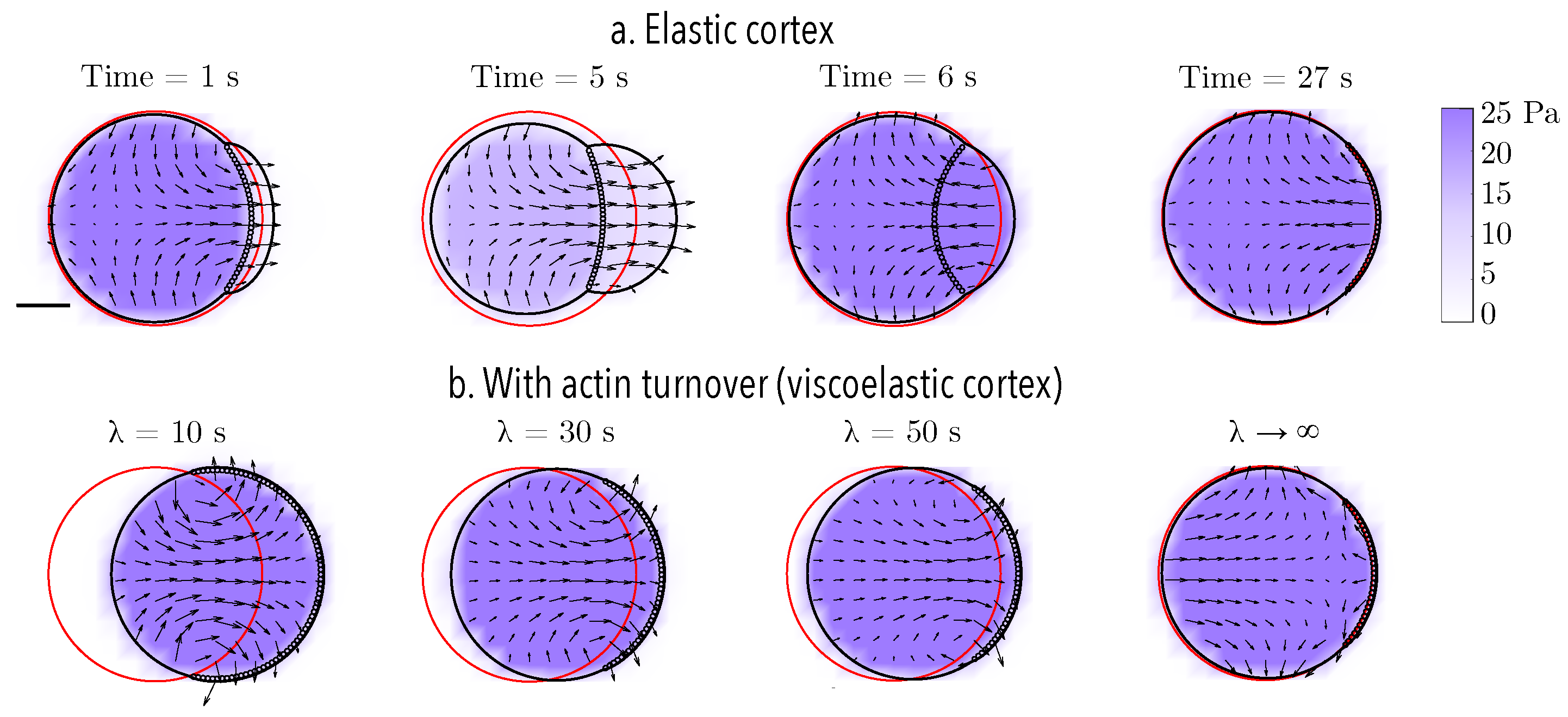
Next, we describe the model for a cellular bleb. In order to account for reformation of the actin cortex within the bleb, we include an additional numerical contour to represent the new cortex in the bleb.
Figure 1 (right) shows the location of the old and new cortex points and the location of membrane-cortex attachment links during blebbing. Cortical elasticity is multiplied by local density
on the new and old cortex. A bleb is initiated by setting the density of the new cortex to zero in a small region at the front of the cell. Adhesive links between the old cortex and the membrane are removed but are maintained in the new cortex. Since the elasticity is zero when a bleb is initialized, the new cortex points stay close to the membrane in the growing bleb. We specify a time for bleb expansion of 5 s to allow a large bleb to form at the cell front [
50]. After 5 s, a bleb moves to the retraction phase.
During bleb retraction, the density of the new (
) and old cortex (
) are updated over time according to the following equations:
The rate of cortex reformation at the bleb site is . Note that densities of the new and old cortex are assumed to be spatially uniform in their respective locations. In the bleb retraction phase, increases over time while monotonically decreases. Once the density on the old cortex reaches a critical value of 0.2 during bleb expansion, the bleb switches from retraction to expansion by resetting the density on the old cortex to a value of 1 and the density of the new cortex to 0. The critical value of 0.2 was chosen because the location of the cortex at this value was found to be very close to its location before the bleb was initiated.
Once a bleb cycle is completed (i.e., after the density on the old cortex reaches a value of 0.2), the old cortex is reset to coincide with membrane points at the start of the new cycle of bleb expansion. The process ensures a re-calibration at the beginning of a new cycle.
Table 1.
Model parameters.
Table 1.
Model parameters.
| Symbol | Quantity | Value | Source |
|---|
| Initial cell radius | 10 m | [28] |
| Membrane surface tension | 40 pN/m | [28] |
| Membrane stiffness coefficient | 4 pN/m | |
| Cortical tension | 400 pN/m | [28] |
| Cortical stiffness coefficient | 100 pN/m | [28] |
| Membrane-cortex attachment stiffness | 267 pN/m | [51] |
| Constant for cell–surface repulsive interaction | 5 kPa | |
| Rate of cortex reformation | 1 s | |
| | Bleb expansion time | 5 s | |
| Cytosolic viscosity | Pa-s | [26,28,52] |
| Cortical drag coefficient | 10 pN-s/m | [51] |
| Strain relaxation time scale | 30 s | |
Given a configuration of the membrane and cortex structures, forces at every location on the structures are computed as described above, and then the pressure and velocity of the fluid, along with velocity of the membrane and cortex are obtained by solving Equations (
1), (
2) and (
7). The position of each structure is updated according to their own respective velocities:
The viscoelastic response of the membrane and cortex structures require each an additional equation for the stress relaxation of the structures, namely Equation (
5).
2.2. Numerical Method
Given an initial configuration of the plasma membrane and cortex, the one-dimensional contours are discretized into a finite number of nodes. At every node on the membrane and cortex, forces are computed according to constitutive laws provided in the previous section. After forces are numerically computed, we use the method of regularized Stokeslets [
45] to solve for the fluid velocity and pressure in Equations (
1) and (
2). In free space, the fluid velocity at the membrane and cortex structures is
where
denotes the regularized Stokeslet matrix with entries
where
and the regularization parameter
which maps smeared cellular forces to fluid velocities at the cellular structures. Here, we use the 2D blob function
from [
45] to spread or regularize a point force density over a small ball around a point
. Values for numerical parameters such as
, the grid spacing of the discretized membrane contour, are listed in
Table 2. Once the fluid velocity is known, the position of the immersed cellular structures are update using the forward Euler time integrator scheme applied to Equations (
11) and (
12).
Although the force balance in Equations (
1) and (
2) ensure zero sum of forces, the introduction of cell-surface interaction can lead to a force imbalance. In 3D, the first term in the Stokeslet decays such as
, where
r is the distance from a point force, whereas in 2D, this term decays like
. Therefore, even a numerically negligible error in sum of forces can result in
as
. To ensure the forces sum to zero, we enforce a no flow boundary condition far away from the physical domain (see
Figure 1 (right)). Although we could have imposed the boundary condition directly on the channel walls as in [
44], our approach allows us to avoid resolving thin fluid boundary layers between the cell and channel wall from satisfying a no-slip boundary condition. Our approach satisfies a no-penetration boundary condition
on the channel wall and was previously used in [
49] to simulate cell deformation in a microfluidic channel with an imposed background flow. To obtain the velocity on membrane and cortex nodes, the linear system in Equation (
13) must then be modified to
The notation
denotes the regularized Stokeslet velocity matrix mapping smeared forces at locations
to velocities at locations
. For example,
describes the effect of cellular forces at
to evaluate velocities of the outer boundary channel
. Note that the method in Equation (
16) ensures that there is no fluid flow at the boundary location, i.e.,
. Alternatively, one can rewrite Equation (
16) for the fluid flow at cellular nodes
as
Pressure is computed as follows,
where
represents the regularized Stokeslet pressure matrix which maps regularized forces at locations
to pressure at locations
[
45]. Thus, pressure along the cellular locations
is given by:
To compute the pressure at arbitrary locations
, which include the cell and external boundary wall forces,
In the limit of small relaxation, our model for viscoelasticity describes a fluid rather than solid; the method does not guarantee to preserve the mesh spacing as the material deforms. Thus, for large deformations, in the case of moving, deforming structure, a re-meshing algorithm maintains resolution of the discretized structures. Here, the protocol is to re-mesh when a bleb cycle is completed (i.e., the density on the old cortex reaches a value of 0.2). In order to re-space the nodes on the cortex and membrane structures uniformly and preserve their strain, a periodic spline function is used to construct differentiable functions from the position of the discrete nodes. The integral of these differentiable functions yields the arclength of the closed curve,
, as a function of current Lagrangian coordinate for the deformed configuration. The inverse map from the Lagrangian coordinate to the corresponding arclength is computed using another periodic spline function,
. Lastly, we define a new equally-spaced arclength function and compute the new parameteric coordinate on the arclength by evaluating the previously formed function. Similarly, a periodic spline function is formed for the tension,
, and it is evaluated at the new parametric coordinate locations,
. A similar re-meshing algorithm was implemented and tested in [
31].
To validate our numerical methods, we simulated 30 s of bleb expansion and retraction with no walls using a membrane and cortex discretization consisting of
and 268 points. We simulated the model with an elastic (
) and viscoelastic cortex (
). In the
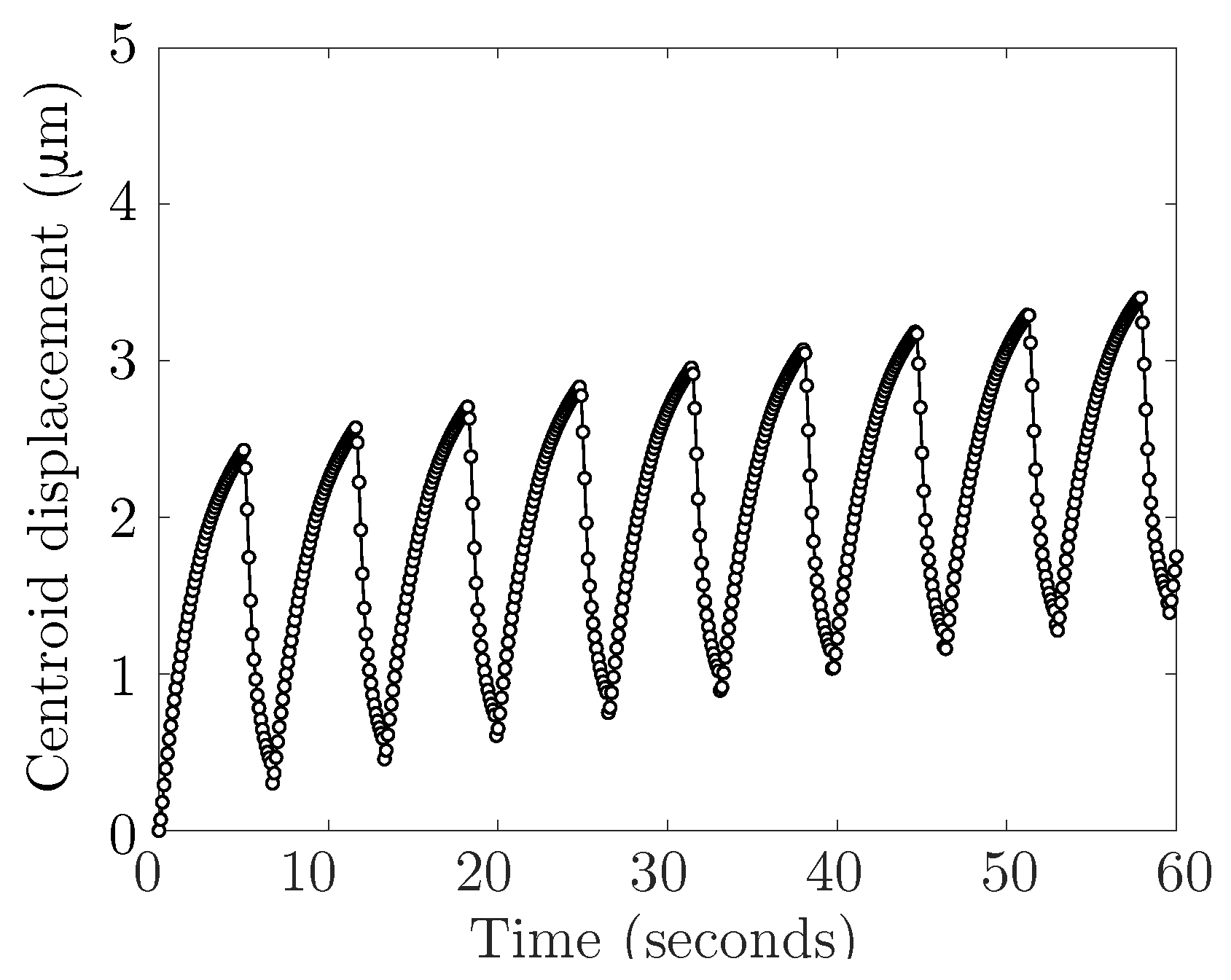
Section 3, we include graphs of the horizontal displacement of the cell centroid over time, and compute cell speed by a linear least-square fit over the first four cycles of bleb expansion and retraction (26.5 s). The graphs for horizontal centroid displacement were indistinguishable for both grid refinements. For the simulation with an elastic cortex, the value of cell speed is
m/min (
), and 0.5447 μm/min (
). The relative percentage error (relative to
) was less than 2%. For the viscoelastic model, cell speed is
μm/min for
and 2.3445 μm/min for
. The relative percentage error (relative to
) is less than 3%. We conclude that simulation data are insensitive to the number of grid points, and use
for the remainder of the paper.
4. Discussion and Conclusions
Model simulations of a blebbing cell with a poroelastic cortex immersed in viscous fluid (and no channel) show that membrane shape changes are reciprocal during bleb expansion and retraction. As a result, a cell cannot swim in zero Reynolds number flow [
36]. Even when a geometric asymmetry is introduced with a curved top channel wall, the cell cannot efficiently migrate. In this case, some motion is possible if a bleb wedges the cell into a gap, but sustained motion does not appear possible under our simplified model assumptions. Our results are contradictory to those in [
44], where swimming between two straight walls with an elastic cortex was observed. One notable difference between our model and [
44] is the force balance on the cortex (Equation (
7)). The cortex model from [
44] was studied in [
53], where the authors found an imbalance in cortical forces led to large pressure relief compared to the model from [
51] that forms the basis for this work. In [
32], the authors found that bleb-based migration without substrate adhesion was possible in a discontinuous environment. We did not consider this type of environment here, but migration in our model may be possible when the cortex is treated as an elastic material and both walls are replaced with spaced point sources.
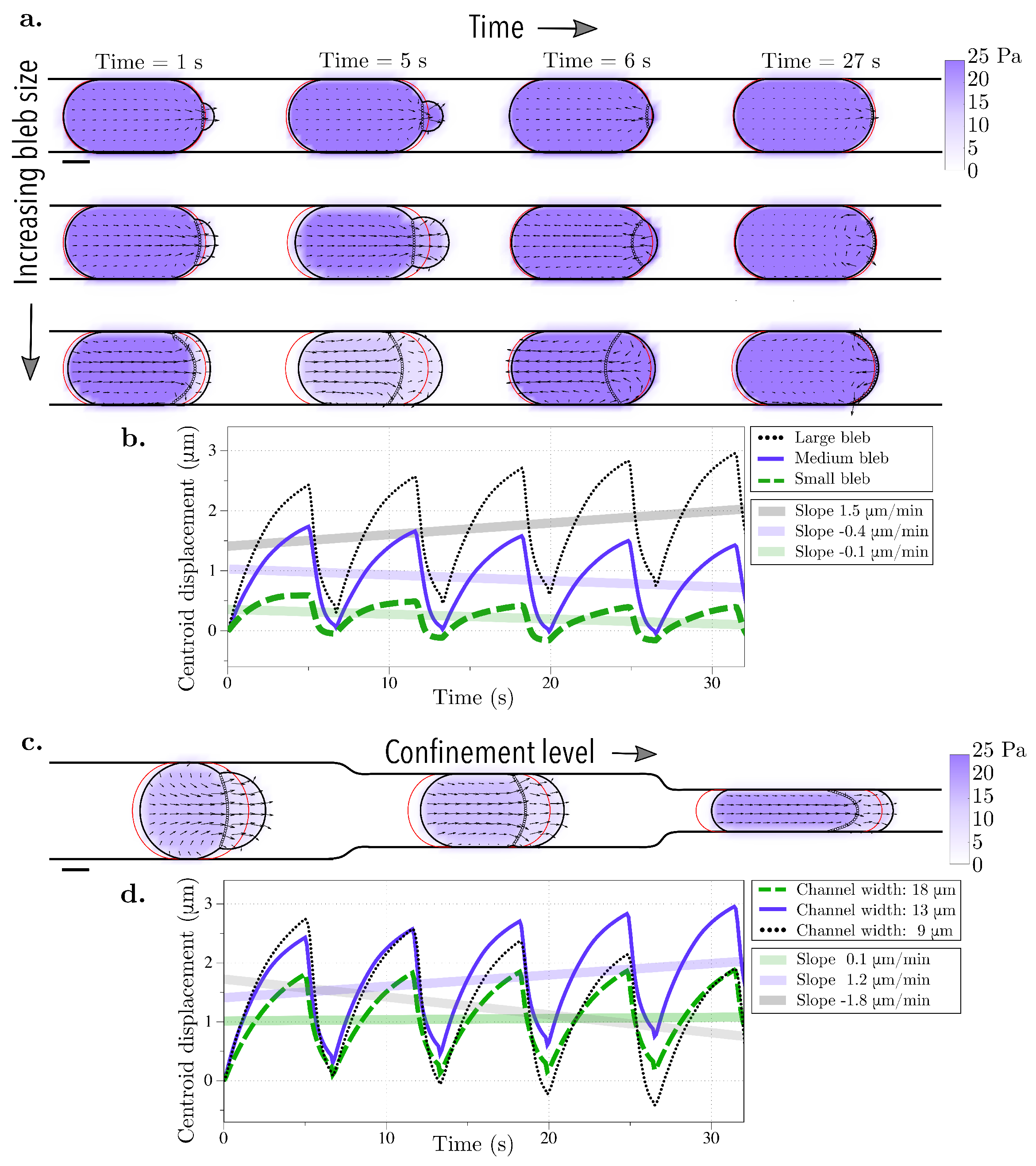
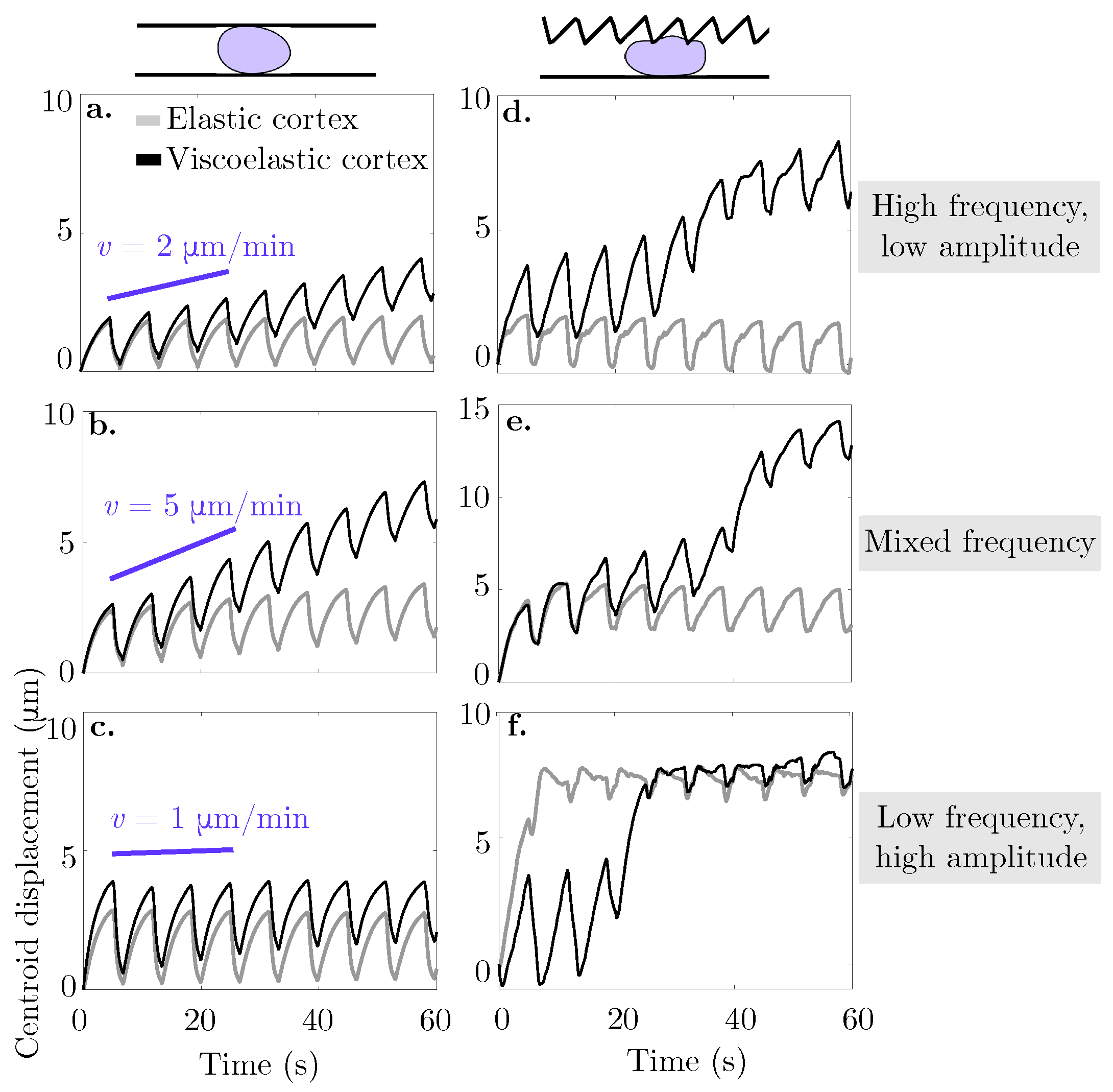
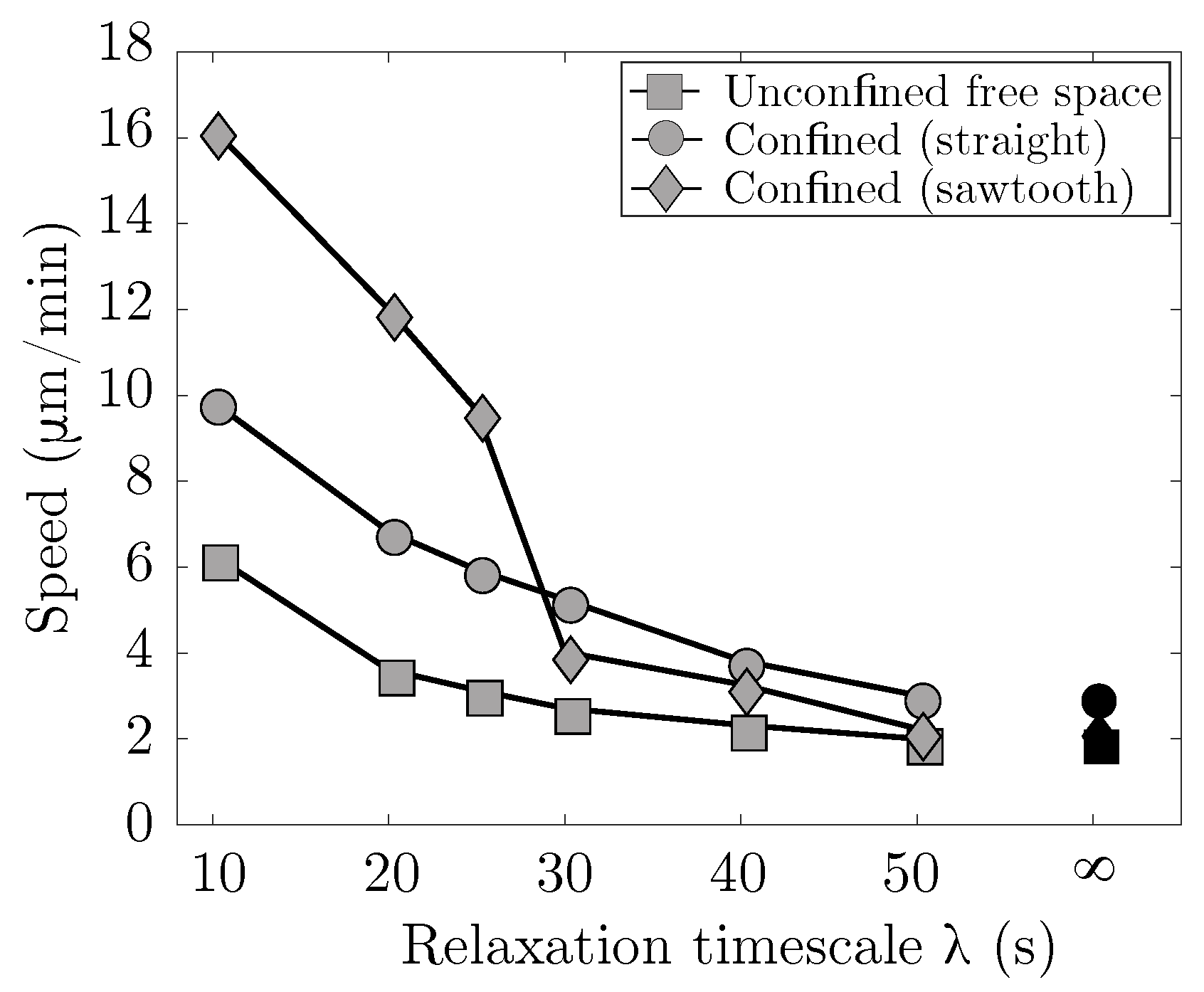
Our results show that the combination of blebbing with cortical actin turnover results in swimming in the absence of confinement. In the context of viscoelasticity, the strain on the cortex evolves in time thus, it creates an asymmetry in the resistance of the material during the bleb expansion and retraction phases. In the case of straight channel walls, we found the fastest migration speed at an intermediate gap size of 13
m (
Figure 6). In [
44], the authors attribute similar results to an increase in intracellular pressure that causes a bleb to form at the rear of the cell. In our model, we only allow blebs to form at the front of the cell and specify that a bleb retracts after 5 s, regardless of the geometry. If we had altered this rule to take membrane speed or diffusion of actin into the bleb, it is possible that the cell speed would be affected.
Our values for cell speed are consistent with those from experimental data in [
46], where confined cell speeds were reported to be 2–6
m/min. In addition, in [
46], the experiments showed a biphasic relationship between confinement and cell speed in the case of horizontal channel walls, with larger speeds at medium confinement levels. We observe this biphasic relationship in our simulations (see
Figure 6). Since our results show actin turnover is necessary for confined migration, confined motility experiments where actin polymerization is perturbed by drugs, such as jasplakinolide, may result in decreased or inhibited migration after treatment.
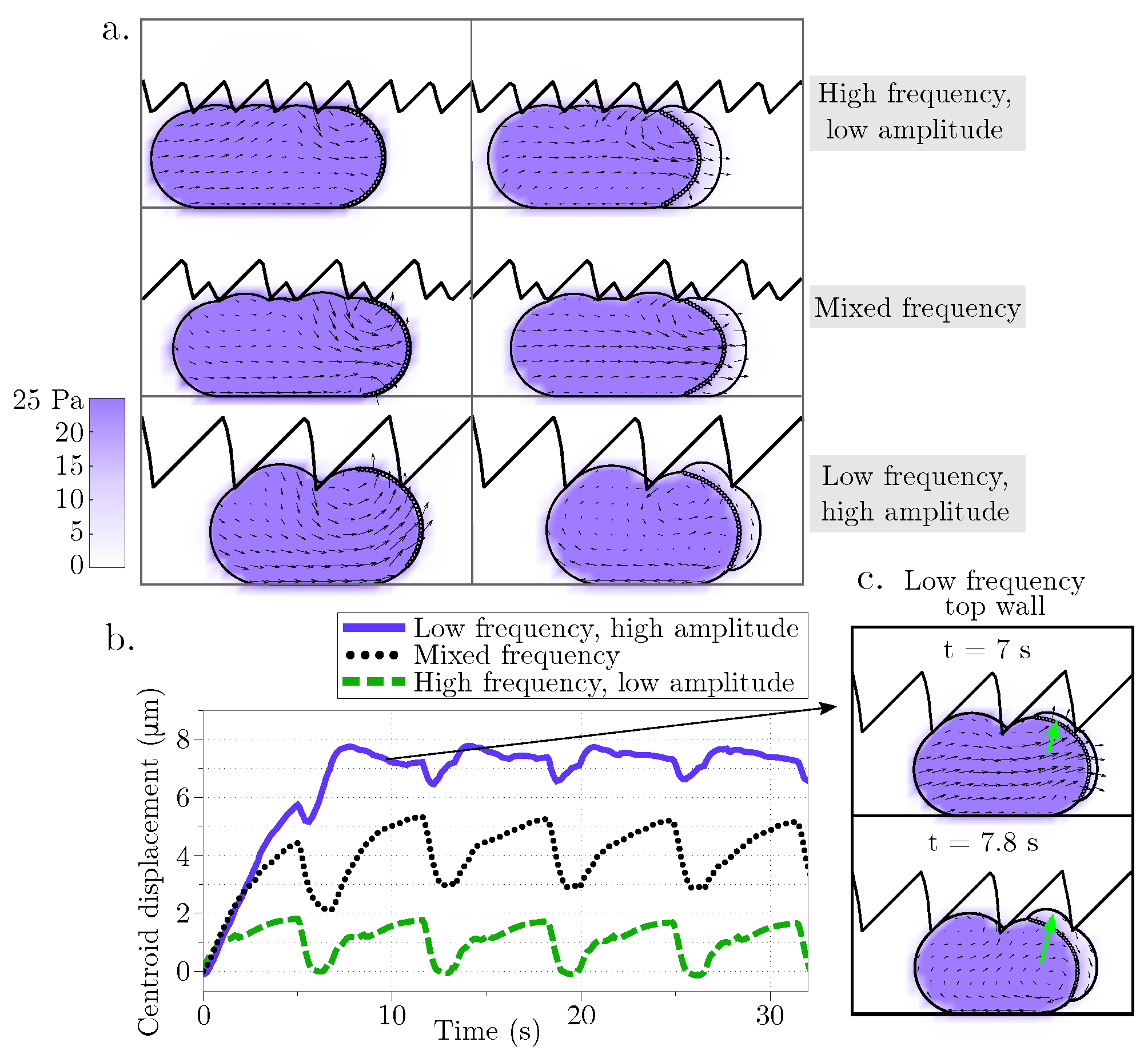
We found that migration speed increases as cortical turnover increases (relaxation timescale
decreases). Interestingly, simulation results show that when
is less than 30 s, cell migration speed is fastest in a geometry when one wall is described by a sawtooth function and the other wall is flat (
Figure 7). As
increases and approaches the case of an elastic cortex, the optimal channel geometry for migration is two flat channel walls. We also observe channel geometries where the cell migrates for a period of time before becoming lodged within a gap, even when the cortex is modeled as a viscoelastic material (see
Figure 6, bottom right). These results point to a non-trivial relationship between the rheology of the cell and its environment. The notion that surface flows can drive adhesion independent migration is not novel to our work [
29]; the authors in [
54] found that surface treadmilling controlled by active RhoA at the cell rear is sufficient to drive directional cellular motility on 2D surfaces and in liquid.
Several computational models have simulated confined or blebbing migration. In [
33], a model for confined migration (chimneying) assumed a mesenchymal mode of motility, driven by actin polymerization at the leading edge, but required proteolysis for migration. Our results on the importance of rearward flow of the cortical actin for 3D migration are in agreement with those from [
33], where a variety of motility mechanisms were studied. Our results differ from those in [
32], where simulations of a confined blebbing cell require substrate adhesion for migration with straight channel walls. One difference between the models is that Kelvin–Voigt viscoelastic elements were used to model the cell membrane and cortex in [
32], whereas we use a Maxwell model in this work.
Our model has several limitations that we hope to address in the future. In order to simulate a viscoelastic material, we re-mesh the membrane and cortex structures after each bleb cycle. The reference configuration can become under-resolved in areas, leading to errors in force computations. In particular, when cortical turnover is fast, the cortex begins to transition from a solid to a fluid, and our algorithm breaks down. In this case, it may be appropriate to simulate the cortex as a fluid. A future direction is to develop better numerical methods to re-mesh the deforming, moving structures and to simulate a thin viscous fluid film immersed in another fluid (such as in [
55]).
A cycle of adhesion-independent bleb-based amoeboid motility is thought to consist of protrusion (bleb), outward forces against the channel or extracellular matrix, and followed by a spatially localized rear contraction [
50]. We have conducted limited studies to explore the effect of introducing localized myosin-driven contraction, but the focus of this manuscript is on the locomotion due to the pressure gradients induced by blebbing. Rear contraction leads to cortical flows that exacerbate the numerical issues previously mentioned. We hope to comment in the future about the effect of tangential and bulk rear local contractions and the timing of contractions in relation to bleb expansion.
In the present work, the channel is modeled as a rigid structure. In several experiments of confined adhesion-independent migration, one channel wall is a glass coverslip, and the other channel wall is agarose gel [
29], which has some elastic properties. It has been suggested that pushing and deforming the channel walls aids the cell’s ability to migrate [
24]. One possible extension of this work is to explore how the material properties of the external environment affect cell migration without specific adhesion.
Finally, a limitation of our model is that it is 2D. The relative simplicity of a 2D model allows us to perform simulations over a range of parameter values. Although impressive 3D simulations of amoeboid cell motility were performed in [
56,
57], they do not include bleb-based motility. A long-term goal is to develop a computationally feasible 3D model that could be used for quantitative comparison to experimental data.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}