
Three-Dimensional-Bioprinted Embedded-Based Cerebral Organoids: An Alternative Approach for Mini-Brain In Vitro Modeling Beyond Conventional Generation Methods

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results and Discussion
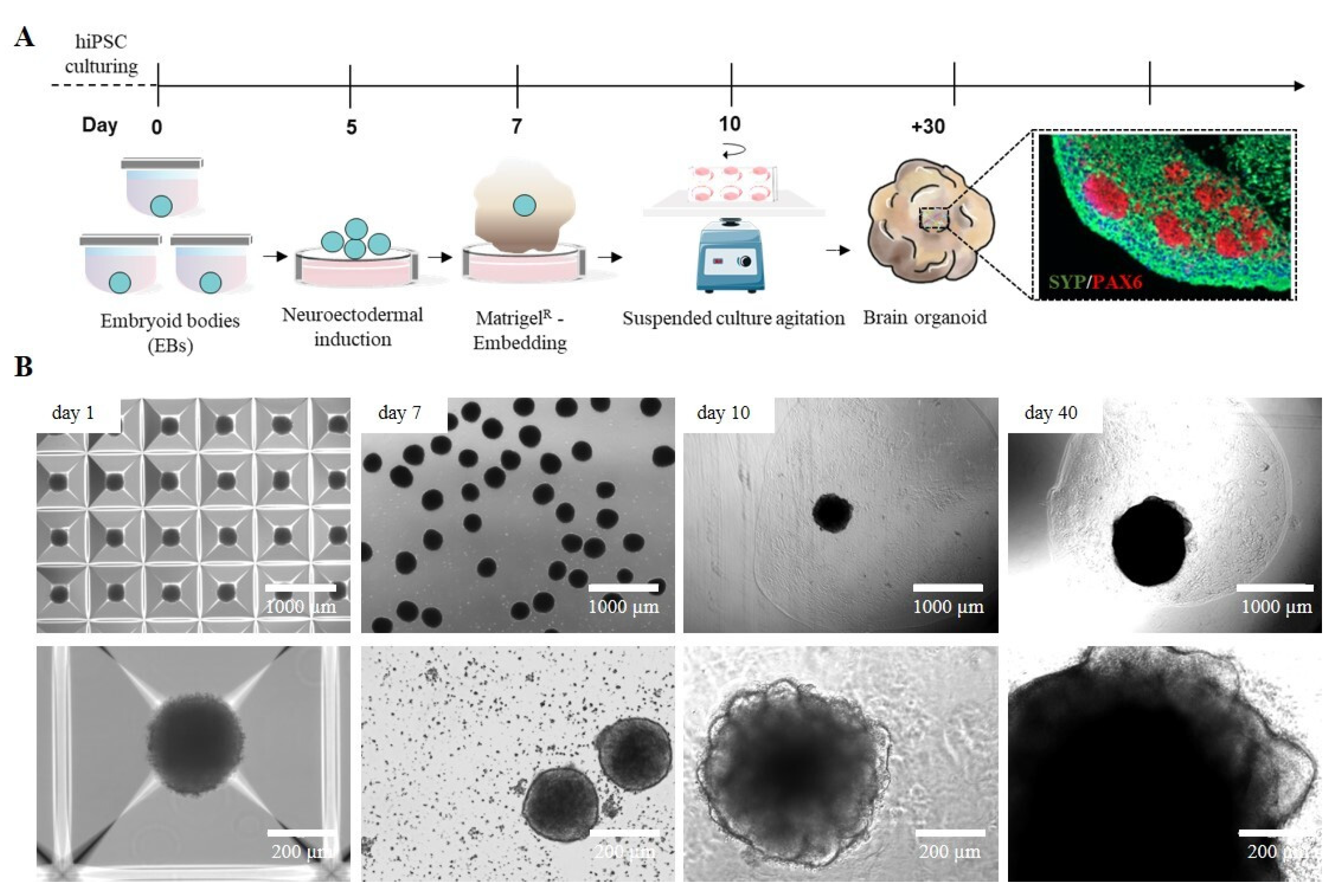
2.1. iPSC-Derived Cerebral Organoid Generation by Manual Matrigel® Embedding
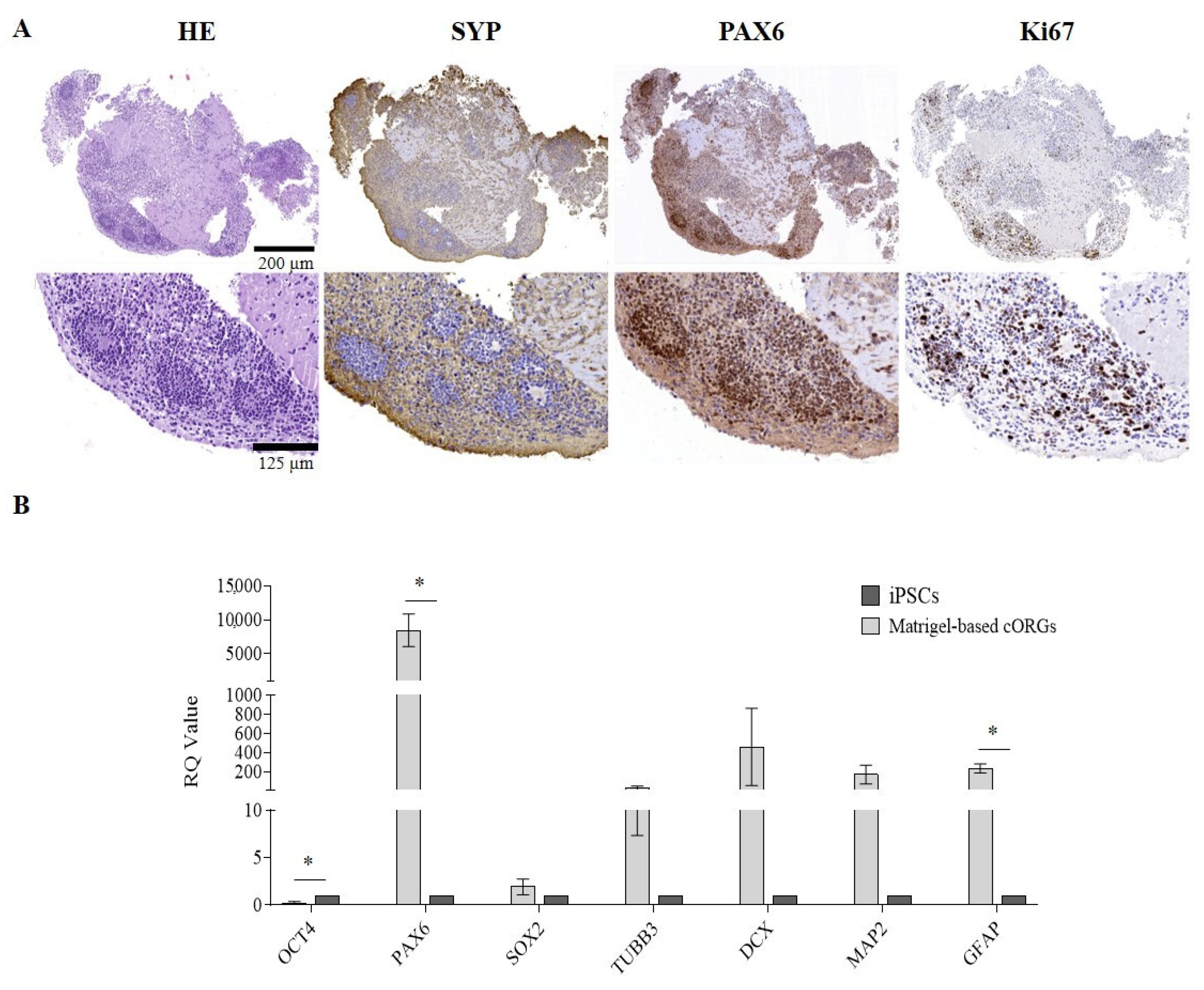
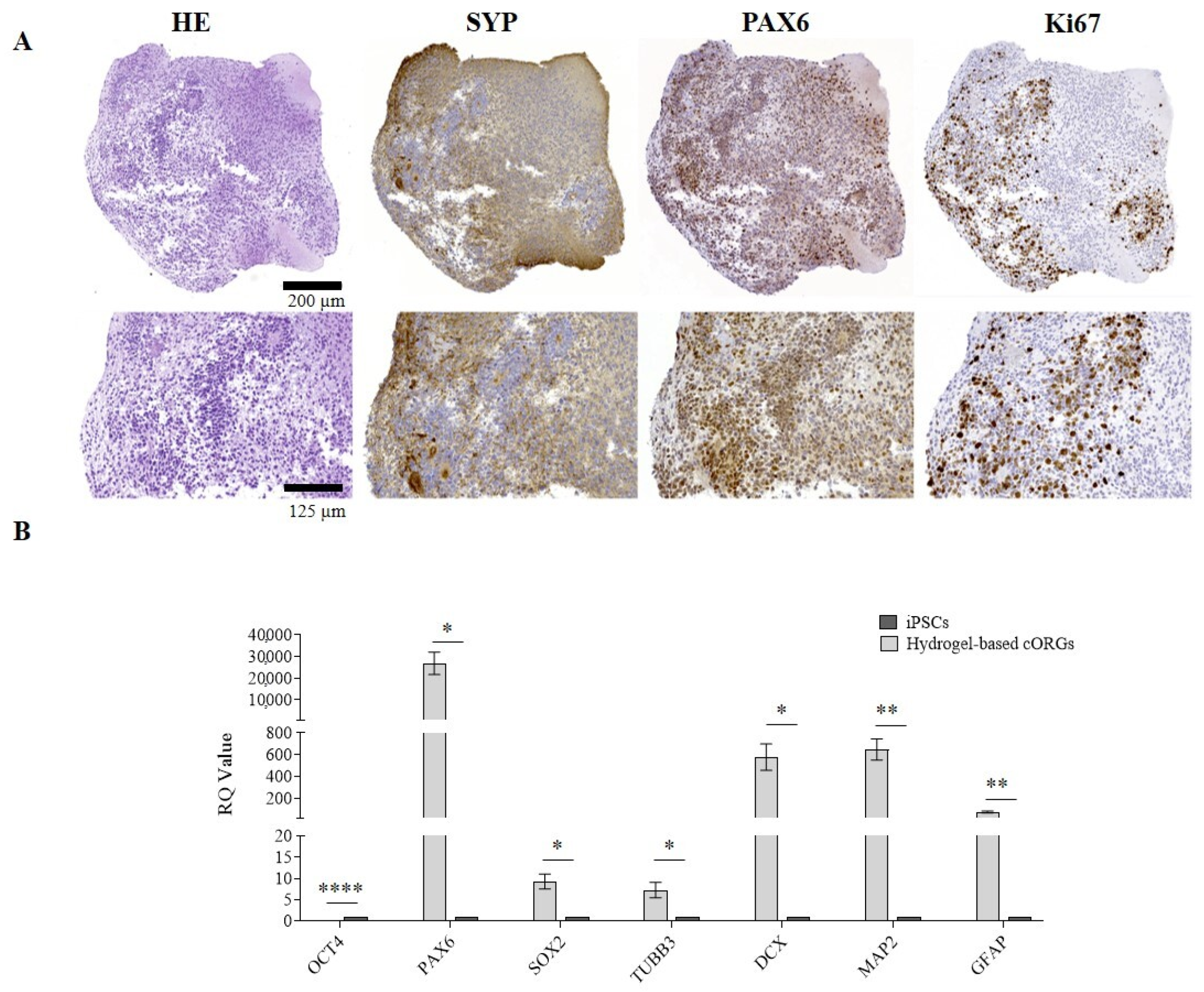
2.2. Manually Matrigel®-Embedded Cerebral Organoid Characterization
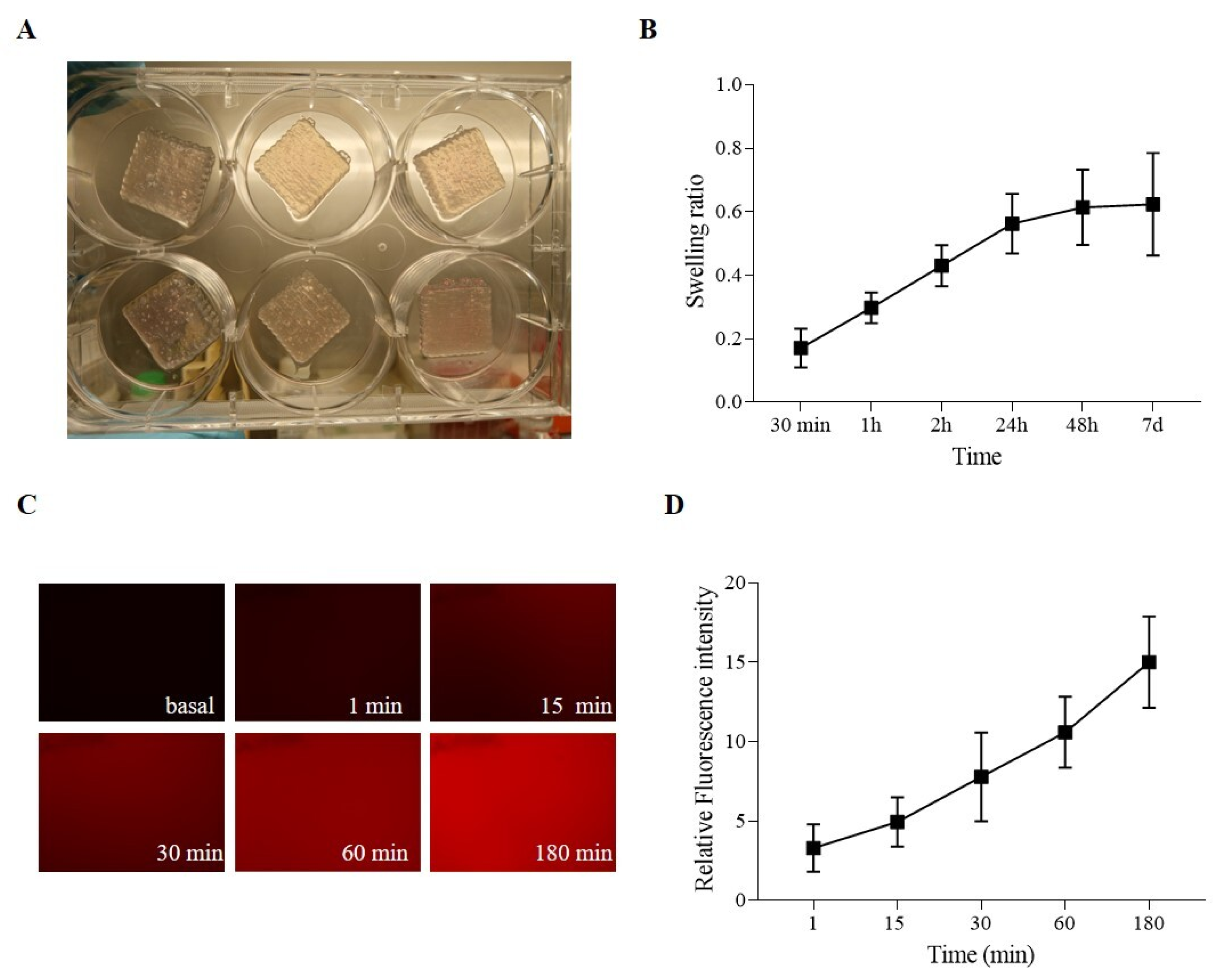
2.3. Hydrogel-Based Ink Stability and Permeability Evaluation
2.4. Hydrogel-Based Ink Printing Process Characterization
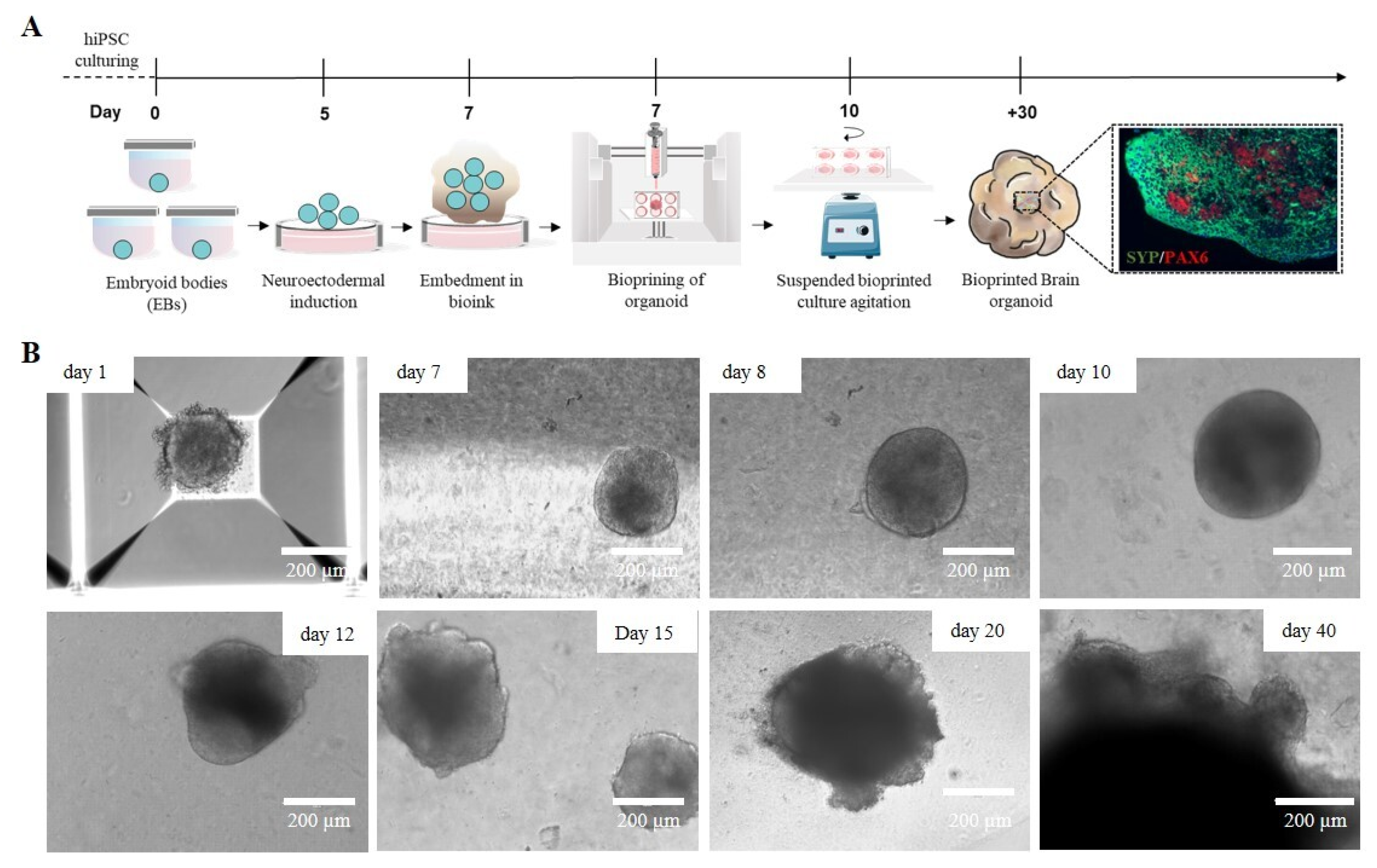
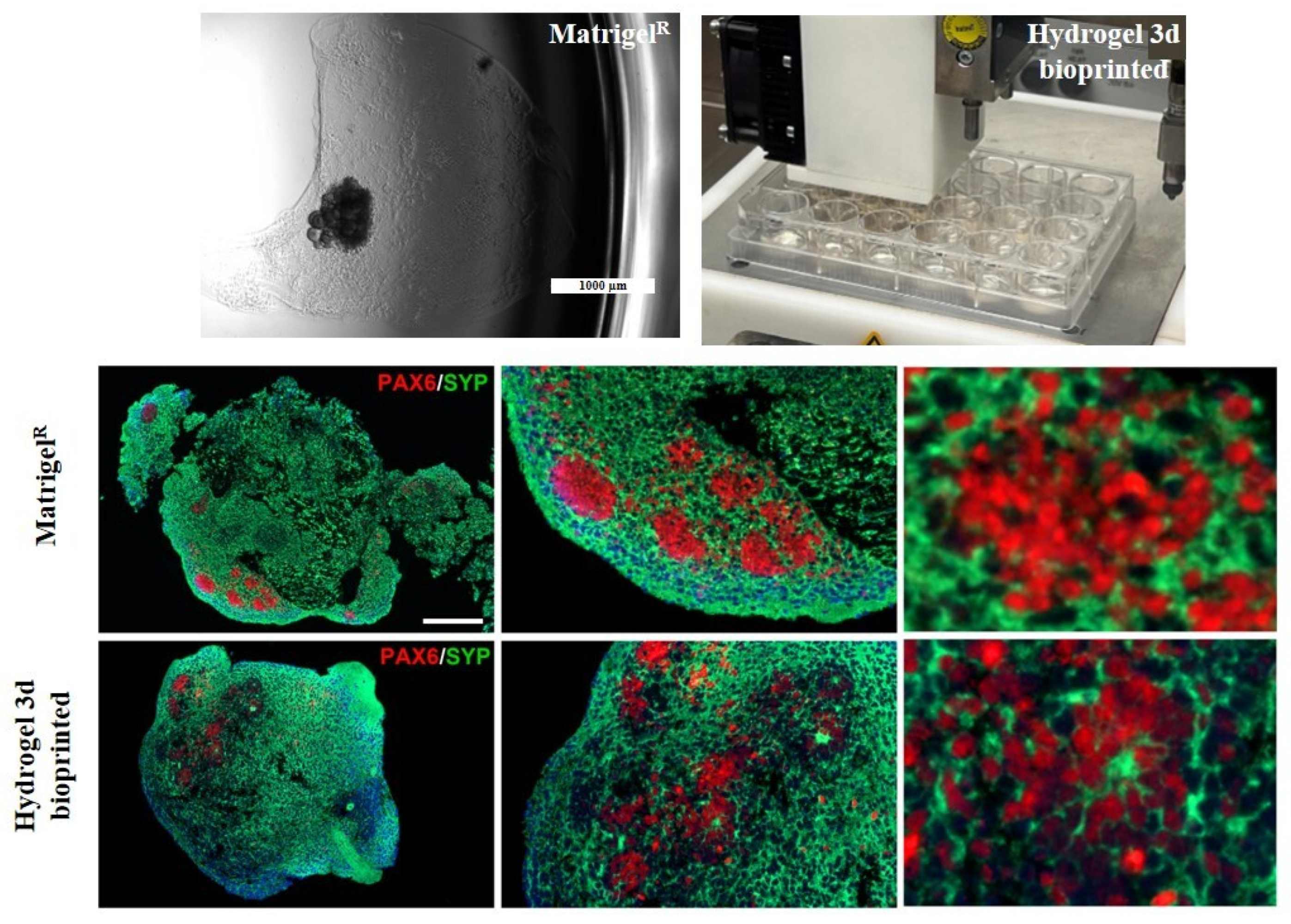
2.5. Bioprinted Hydrogel-Based Cerebral Organoid Generation
2.6. Bioprinted Hydrogel-Based Cerebral Organoid Characterization
2.7. Comparison Between Matrigel®-Based and Bioprinted Hydrogel-Based Cerebral Organoids
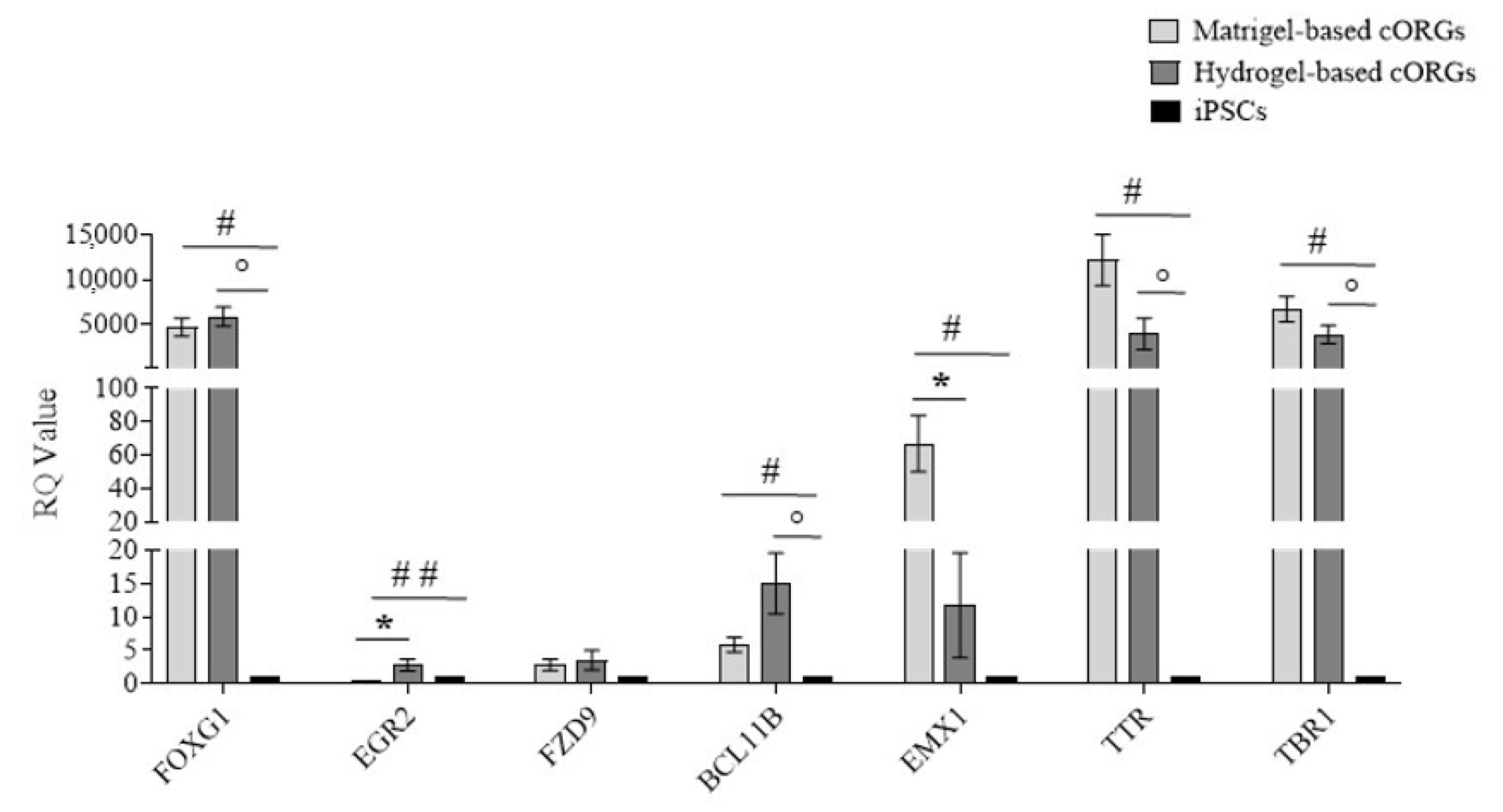
2.8. Early Brain Regionalization Evaluation in Matrigel®-Based and Bioprinted Hydrogel-Based Cerebral Organoids
3. Conclusions
4. Materials and Methods
4.1. Stem Cell Culture
4.2. Cerebral Organoid Generation and Culture Conditions
4.3. Immunohistochemistry and Immunofluorescence
4.4. RNA Extraction and qPCR
4.5. Hydrogel Bio-Ink Characterization and 3D Printing Parameters
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duval, K.; Grover, H.; Han, L.H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Benito-Kwiecinski, S.; Lancaster, M.A. Brain Organoids: Human Neurodevelopment in a Dish. Cold Spring Harb. Perspect. Biol. 2020, 12, 035709. [Google Scholar] [CrossRef]
- Olgasi, C.; Cucci, A.; Follenzi, A. iPSC-Derived Liver Organoids: A Journey from Drug Screening, to Disease Modeling, Arriving at Regenerative Medicine. Int. J. Mol. Sci. 2020, 17, 6215. [Google Scholar] [CrossRef] [PubMed]
- Karakikes, I.; Ameen, M.; Termglinchan, V.; Wu, J.C. Human induced pluripotent stem cell-derived cardiomyocytes: Insights into molecular, cellular, and functional phenotypes. Circ. Res. 2015, 1, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Liu, Y.; Ray, E.; Giuliano, A.E.; Cui, X. Human Breast Organoid Models for Lactation Research. Reprod. Breed. 2023, 3, 125–130. [Google Scholar] [CrossRef]
- Pagliuca, F.W.; Millman, J.R.; Gürtler, M.; Segel, M.; Van Dervort, A.; Ryu, J.H.; Peterson, Q.P.; Greiner, D.; Melton, D.A. Generation of functional human pancreatic β cells In Vitro. Cell 2014, 2, 428–439. [Google Scholar] [CrossRef]
- Lindborg, B.A.; Brekke, J.H.; Vegoe, A.L.; Ulrich, C.B.; Haider, K.T.; Subramaniam, S.; Venhuizen, S.L.; Eide, C.R.; Orchard, P.J.; Chen, W.; et al. Rapid Induction of Cerebral Organoids From Human Induced Pluripotent Stem Cells Using a Chemically Defined Hydrogel and Defined Cell Culture Medium. Stem Cells Transl. Med. 2016, 7, 970–979. [Google Scholar] [CrossRef]
- Qian, X.; Song, H.; Ming, G.L. Brain organoids: Advances, applications and challenges. Development 2019, 8, 166074. [Google Scholar] [CrossRef]
- Acharya, P.; Choi, N.Y.; Shrestha, S.; Jeong, S.; Lee, M.Y. Brain organoids: A revolutionary tool for modeling neurological disorders and development of therapeutics. Biotechnol. Bioeng. 2024, 2, 489–506. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 7467, 373–379. [Google Scholar] [CrossRef]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement membrane matrix with biological activity. Semin. Cancer Biol. 2005, 5, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, M.T.; Crook, C.J.; Ku, H.T. Towards organoid culture without Matrigel. Commun. Biol. 2021, 4, 1387. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, Z.X.; Stappenbeck, T.S.; Miner, J.H. Laminin alpha 5 influences the architecture of the mouse small intestine mucosa. J. Cell Sci. 2008, 121 Pt 15, 2493–2502. [Google Scholar] [CrossRef]
- Aisenbrey, E.A.; Murphy, W.L. Synthetic alternatives to Matrigel. Nat. Rev. Mater. 2020, 5, 539–551. [Google Scholar] [CrossRef]
- Hospodiuk, M.; Dey, M.; Sosnoski, D.; Ozbolat, I.T. The bioink: A comprehensive review on bioprintable materials. Biotechnol. Adv. 2017, 2, 217–239. [Google Scholar] [CrossRef]
- Zhu, L.; Yuhan, J.; Yu, H.; Zhang, B.; Huang, K.; Zhu, L. Decellularized Extracellular Matrix for Remodeling Bioengineering Organoid’s Microenvironment. Small 2023, 25, 2207752. [Google Scholar] [CrossRef] [PubMed]
- Rovetta, R.; Ginestra, P.; Ferraro, R.M.; Zohar-Hauber, K.; Giliani, S.; Ceretti, E. Building Orientation and Post Processing of Ti6Al4V Produced by Laser Powder Bed Fusion Process. J. Manuf. Mater. Process. 2023, 7, 43. [Google Scholar] [CrossRef]
- Dado, D.; Levenberg, S. Cell-scaffold mechanical interplay within engineered tissue. Semin. Cell Dev. Biol. 2009, 6, 656–664. [Google Scholar] [CrossRef]
- Seiti, M.; Ginestra, P.S.; Ceretti, E.; Ferraris, E.; Ranga, A. Emerging Three-Dimensional Integrated Systems for Biomimetic Neural In Vitro Cultures. Adv. Mater. Interfaces 2022, 9, 2101297. [Google Scholar] [CrossRef]
- Tasoglu, S.; Demirci, U. Bioprinting for stem cell research. Trends Biotechnol. 2013, 1, 10–19. [Google Scholar] [CrossRef]
- Gungor-Ozkerim, P.S.; Inci, I.; Zhang, Y.S.; Khademhosseini, A.; Dokmeci, M.R. Bioinks for 3D bioprinting: An overview. Biomater. Sci. 2018, 5, 915–946. [Google Scholar] [CrossRef]
- Groll, J.; Burdick, J.A.; Cho, D.W.; Derby, B.; Gelinsky, M.; Heilshorn, S.C.; Jüngst, T.; Malda, J.; Mironov, W.; Nakayama, K.; et al. A definition of bioinks and their distinction from biomaterial inks. Biofabrication 2018, 11, 013001. [Google Scholar] [CrossRef]
- Tian, S.; Zhao, H.; Lewinski, H. Key parameters and applications of extrusion-based bioprinting. Bioprinting 2021, 23, e00156. [Google Scholar] [CrossRef]
- Ur Rehman, W.; Asim, M.; Hussain, S.; Khan, S.A.; Khan, S.B. Hydrogel: A Promising Material in Pharmaceutics. Curr. Pharm. Des. 2020, 26, 5892–5908. [Google Scholar] [CrossRef]
- Bedell, M.L.; Navara, A.M.; Du, Y.; Zhang, S.; Mikos, A.G. Polymeric Systems for Bioprinting. Chem. Rev. 2020, 120, 10744–10792. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.E.; Janmey, P.; Wang, Y. Tissue Cells Feel and Respond to the Stiffness of Their Substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Ferraro, R.M.; Ginestra, P.S.; Giliani, S.; Ceretti, E. Carbonization of polymer precursor substrates to direct human iPSC-derived neurons differentiation and maturation. Procedia CIRP 2020, 89, 39–44. [Google Scholar] [CrossRef]
- Piras, C.C.; Smith, D.K. Multicomponent polysaccharide alginate-based bioinks. J. Mater. Chem. B 2020, 36, 8171–8188. [Google Scholar] [CrossRef]
- Mei, Q.; Rao, J.; Bei, H.P.; Liu, Y.; Zhao, X. 3D Bioprinting Photo-Crosslinkable Hydrogels for Bone and Cartilage Repair. Int. J. Bioprint. 2021, 3, 367. [Google Scholar] [CrossRef]
- Rastogi, P.; Kandasubramanian, B. Review of alginate-based hydrogel bioprinting for application in tissue engineering. Biofabrication 2019, 4, 042001. [Google Scholar] [CrossRef]
- Durfee, W.K.; Iaizzo, P.A. Medical Applications of 3D Printing. In Engineering in Medicine; Academic Press: Cambridge, MA, USA, 2019; pp. 527–543. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 6194, 1247125. [Google Scholar] [CrossRef]
- Hong, W.; Liang, Y.; Yanning, Y. A review of sodium alginate-based hydrogels: Structure, mechanisms, applications, and perspectives. Int. J. Biol. Macromol. 2025, 292, 139151. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Xuan, Y.; Zhang, S. Synthesis and Applications of Carboxymethyl Cellulose Hydrogels. Gels 2022, 8, 529. [Google Scholar] [CrossRef]
- Barros, D.; Conde-Sousa, E.; Gonçalves, A.M.; Han, W.M.; García, A.J.; Amaral, I.F.; Pêgo, A.P. Engineering hydrogels with affinity-bound laminin as 3D neural stem cell culture systems. Biomater. Sci. 2019, 12, 5338–5349. [Google Scholar] [CrossRef]
- Rauch, U. Brain matrix: Structure, turnover and necessity. Biochem. Soc. Trans. 2007, 4, 656–660. [Google Scholar] [CrossRef]
- Layrolle, P.; Payoux, P.; Chavanas, S. Message in a Scaffold: Natural Biomaterials for Three-Dimensional (3D) Bioprinting of Human Brain Organoids. Biomolecules 2023, 13, 25. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, R.; Hu, Z.; Ni, R.; Zhu, T.; Zhang, H.; Zhu, Y. Gel-Based Suspension Medium Used in 3D Bioprinting for Constructing Tissue/Organ Analogs. Gels 2024, 10, 644. [Google Scholar] [CrossRef]
- Akbarzadeh Solbu, A.; Caballero, D.; Damigos, S.; Kundu, S.; Reis, L.; Halaas, O.; Chahal, A.; Strand, B. Assessing cell migration in hydrogels: An overview of relevant materials and methods. Mater. Today Bio 2023, 18, 100537. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Corsini, N.S.; Wolfinger, S.; Gustafson, E.H.; Phillips, A.W.; Burkard, T.R.; Otani, T.; Livesey, F.J.; Knoblich, J.A. Guided self-organization and cortical plate formation in human brain organoids. Nat. Biotechnol. 2017, 35, 659–666. [Google Scholar] [CrossRef]
- Estridge, R.C.; O’Neill, J.E.; Keung, A.J. Matrigel Tunes H9 Stem Cell-Derived Human Cerebral Organoid Development. Organoids 2023, 2, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.; Qin, X.; Liu, H.; Liu, J.; Qin, J. Recent advances in defined hydrogels in organoid research. Bioact. Mater. 2023, 28, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Chiaradia, I.; Lancaster, M.A. Brain organoids for the study of human neurobiology at the interface of in vitro and In Vivo. Nat. Neurosci. 2020, 12, 1496–1508. [Google Scholar] [CrossRef]
- Shaker, M.R.; Slonchak, A.; Al-Mhanawi, B.; Morrison, S.D.; Sng, J.D.J.; Cooper-White, J.; Khromykh, A.A.; Wolvetang, E.J. Choroid plexus defects in Down syndrome brain organoids enhance neurotropism of SARS-CoV-2. Sci. Adv. 2024, 23, 4735. [Google Scholar] [CrossRef]
- Galli, J.; Gavazzi, F.; De Simone, M.; Giliani, S.; Garau, J.; Valente, M.; Vairo, D.; Cattalini, M.; Mortilla, M.; Andreoli, L.; et al. Sine causa tetraparesis: A pilot study on its possible relationship with interferon signature analysis and Aicardi-Goutières syndrome-related genes analysis. Medicine 2018, 52, 13893. [Google Scholar] [CrossRef]
- Morales, X.; Cortés-Domínguez, I.; Ortiz-de-Solorzano, C. Modeling the Mechanobiology of Cancer Cell Migration Using 3D Biomimetic Hydrogels. Gels 2021, 7, 17. [Google Scholar] [CrossRef]
- Gu, Q.; Tomaskovic-Crook, E.; Lozano, R.; Chen, Y.; Kapsa, R.M.; Zhou, Q.; Wallace, G.G.; Crook, J.M. Functional 3D Neural Mini-Tissues from Printed Gel-Based Bioink and Human Neural Stem Cells. Adv. Healthc. Mater. 2016, 5, 1429–1438. [Google Scholar] [CrossRef]
- Bozza, A.; Coates, E.E.; Incitti, T.; Ferlin, K.M.; Messina, A.; Menna, E.; Bozzi, Y.; Fisher, J.P.; Casarosa, S. Neural Differentiation of Pluripotent Cells in 3D Alginate-Based Cultures. Biomaterials 2014, 35, 4636–4645. [Google Scholar] [CrossRef]
- Gilmozzi, V.; Gentile, G.; Riekschnitz, D.A.; Von Troyer, M.; Lavdas, A.A.; Kerschbamer, E.; Weichenberger, C.X.; Rosato-Siri, M.D.; Casarosa, S.; Conti, L.; et al. Generation of HiPSC-Derived Functional Dopaminergic Neurons in Alginate-Based 3D Culture. Front. Cell Dev. Biol. 2021, 9, 708389. [Google Scholar] [CrossRef]
- Trujillo, S.; Seow, M.; Lueckgen, A.; Salmeron-Sanchez, M.; Cipitria, A. Dynamic Mechanical Control of Alginate-Fibronectin Hydrogels with Dual Crosslinking: Covalent and Ionic. Polymers 2021, 13, 433. [Google Scholar] [CrossRef]
- Tomaskovic-Crook, E.; Higginbottom, S.; Zhang, B.; Bourke, J.; Wallace, G.; Crook, J. Defined, Simplified, Scalable, and Clinically Compatible Hydrogel-Based Production of Human Brain Organoids. Organoids 2023, 2, 20–36. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Cui, K.; Guo, Y.; Zhang, X.; Qin, J. Advances in Hydrogels in Organoids and Organs-on-a-Chip. Adv. Mater. 2019, 31, e1902042. [Google Scholar] [CrossRef] [PubMed]
- Gimondi, S.; Ferreira, H.; Reis, R.L.; Neves, N.M. Microfluidic Devices: A Tool for Nanoparticle Synthesis and Performance Evaluation. ACS Nano 2023, 17, 14205–14228. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, R.M.; Masneri, S.; Lanzi, G.; Barisani, C.; Piovani, G.; Savio, G.; Cattalini, M.; Galli, J.; Cereda, C.; Muzi-Falconi, M.; et al. Establishment of three iPSC lines from fibroblasts of a patient with Aicardi Goutières syndrome mutated in RNaseH2B. Stem Cell Res. 2019, 41, 101620. [Google Scholar] [CrossRef]
- Seiti, M.; Mazzoldi, E.; Pandini, S.; Giliani, S.; Ferraris, E.; Ginestra, P.S.; Ceretti, E. FRESH 3D Bioprinting of Alginate—Cellulose Gelatin Constructs for Soft Tissue Biofabrication. Procedia CIRP 2024, 125, 42–47. [Google Scholar] [CrossRef]
- Sughi, S.; Seiti, M.; Gaudenzi, G.; Ginestra, P.S. Assessing the Viscosity of Alginate—Cellulose-Based Hydrogels: A Comparison Among Different Types of Solutes, Mediums, Culture, and Gelatin Influence; ISIEA 2023, Lecture Notes in Networks and Systems; Springer: Cham, Switzerland, 2023; Volume 745. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward 5’-3’ | Reverse 5’-3’ |
|---|---|---|
| OCT4 | CCTCACTTCACTGCACTGTA | CAGGTTTTCTTTCCCTAGCT |
| PAX6 | CTGAAGCGGAAGCTGCAAAG | TTGCTGGCCTGTCTTCTCTG |
| SOX2 | CCCAGCAGACTTCACATGT | CCTCCCATTTCCCTCGTTTT |
| TUBB3 | GGCCAAGTTCTGGGAAGTCAT | CTCGAGGCACGTACTTGTGA |
| DCX | TATGCGCCGAAGCAAGTCTC | TACAGGTCCTTGTGCTTCCG |
| MAP2 | GACTGCAGCTCTGCCTTTAG | AAGTAAATCTTCCTCCACTGTGAC |
| GFAP | GAGGTTGAGAGGGACAATCTGG | GTGGCTTCATCTGCTTCCTGTC |
| FOXG1 | ACAGCTCCGTGTTGACTCAG | AGGGGTTGAGGGAGTAGGTC |
| EGR2 | TTGACCAGATGAACGGAGTG | CTTGCCCATGTAAGTGAAGGT |
| FZD9 | GCGAGAACCCCGAGAAGTT | GTGAAGGCGGTGGAGAAGAA |
| BCL11B | GCCAGTGTCAGTTGTCAGGT | AGGTTGAAGGGGTTGCTGTC |
| EMX1 | GAGACGCAGGTGAAGGTGTG | CTCGTGGGTTTGTGGTTGC |
| TTR | TGGCTTCTCATCGTCTGCTC | CGGAGTCGTTGGCTGTGAAT |
| TBR1 | ACAATGGGCAGATGGTGGTT | TGACGGCGATGAACTGAGTC |
| βACTIN | CGCCGCCAGCTCACCATG | CACGATGGAGGGGAAGACGG |
| HSP90AB1 | TCCGGCGCAGTGTTGGGAC | TCCATGGTGCACTTCCTCAGGC |
| NANOG | TGAACCTCAGCTACAAACAG | TGGTGGTAGGAAGAGTAAAG |
| OCT4 | CCTCACTTCACTGCACTGTA | CAGGTTTTCTTTCCCTAGCT |
| SOX2 | CCCAGCAGACTTCACATGT | CCTCCCATTTCCCTCGTTTT |
| KLF4 | GATGAACTGACCAGGCACTA | GTGGGTCATATCCACTGTCT |
| cMYC | TGCCTCAAATTGGACTTTGG | GATTGAAATTCTGTGTAACTGC |
| Mycoplasma detection | ACTCCTACGGGAGGCAGCAGTA | TGCACCATCTGTCACTCTGTTAACCTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferraro, R.M.; Ginestra, P.S.; Seiti, M.; Bugatti, M.; Benini, G.; Ottelli, L.; Vermi, W.; Poliani, P.L.; Ceretti, E.; Giliani, S. Three-Dimensional-Bioprinted Embedded-Based Cerebral Organoids: An Alternative Approach for Mini-Brain In Vitro Modeling Beyond Conventional Generation Methods. Gels 2025, 11, 284. https://doi.org/10.3390/gels11040284
Ferraro RM, Ginestra PS, Seiti M, Bugatti M, Benini G, Ottelli L, Vermi W, Poliani PL, Ceretti E, Giliani S. Three-Dimensional-Bioprinted Embedded-Based Cerebral Organoids: An Alternative Approach for Mini-Brain In Vitro Modeling Beyond Conventional Generation Methods. Gels. 2025; 11(4):284. https://doi.org/10.3390/gels11040284
Chicago/Turabian StyleFerraro, Rosalba Monica, Paola Serena Ginestra, Miriam Seiti, Mattia Bugatti, Gabriele Benini, Luana Ottelli, William Vermi, Pietro Luigi Poliani, Elisabetta Ceretti, and Silvia Giliani. 2025. "Three-Dimensional-Bioprinted Embedded-Based Cerebral Organoids: An Alternative Approach for Mini-Brain In Vitro Modeling Beyond Conventional Generation Methods" Gels 11, no. 4: 284. https://doi.org/10.3390/gels11040284
APA StyleFerraro, R. M., Ginestra, P. S., Seiti, M., Bugatti, M., Benini, G., Ottelli, L., Vermi, W., Poliani, P. L., Ceretti, E., & Giliani, S. (2025). Three-Dimensional-Bioprinted Embedded-Based Cerebral Organoids: An Alternative Approach for Mini-Brain In Vitro Modeling Beyond Conventional Generation Methods. Gels, 11(4), 284. https://doi.org/10.3390/gels11040284