Abstract
Asexual and sexual morphs of saprobic bambusicolous fungi were collected from freshwater and terrestrial habitats in Sichuan Province, China. Taxonomic identification of these fungi was carried out on the basis of morphological comparison, culture characteristics, and molecular phylogeny. Multi-gene phylogeny based on combined SSU, ITS, LSU, rpb2, and tef1α sequence data was performed to determine their phylogenetic placement, and the result showed that these fungi belong to Savoryellaceae. Morphologically, four asexual morphs are similar to Canalisporium and Dematiosporium, while a sexual morph well-fits to Savoryella. Three new species, Canalisporium sichuanense, Dematiosporium bambusicola, and Savoryella bambusicola are identified and described. Two new records, C. dehongense and D. aquaticum, were recovered from the bamboo hosts in terrestrial and freshwater habitats, respectively. In addition, the nomenclatural confusion of C. dehongense and C. thailandense is discussed.
1. Introduction
Bamboo fungi are a highly diverse group of organisms [1,2,3,4,5], and research on bambusicolous fungi also provides opportunities to control bamboo diseases and promote bamboo cultivation [6]. Approximately 150 basidiomycetes and 1150 ascomycetes have been reported from bamboo, including 350 asexual morphs, 240 hyphomycetes, and 110 coelomycetes [4,6]. China is rich in bamboo resources, and its bamboo species account for more than 50% of the world’s bamboo [7,8]. Sichuan Province has a variety of bamboo forests and inhabits a large number of bamboo fungi due to its complex topography and climate conditions [9]. Some well-represented families of bamboo fungi in Sichuan Province are Astrosphaeriellaceae [10], Bambusicolaceae [9,11], Occultibambusaceae [9,12] distributed in Dothideomycetes, and Apiosporaceae [12,13], Nectriaceae [10,14], Phyllachoraceae [10,15] distributed in Sordariomycetes.
Savoryellaceae (Savoryellales, Sordariomycetes), with an estimated divergence crown age of 182 MYA [16], contains generally saprobic fungi on bamboos, palms, Pandanus, Machilus sp., Pinus sp., or other unidentified woody substrates in terrestrial, freshwater, marine, brackish water, and water-cooling towers [16,17,18,19,20,21,22]. The family currently comprises six genera: Ascotaiwania, Bactrodesmium, Canalisporium, Dematiosporium, Neoascotaiwania, and Savoryella [17,23], which are characterized by immersed or superficial, globose to pyriform ascomata with paraphyses, two-to-eight-spored, clavate to cylindrical, unitunicate asci with an inamyloid apical ring, and ellipsoid, fusiform ascospores, with or without a gelatinous sheath, as well as a dematiaceous hyphomycetous asexual morph with globose to subglobose or obovate to oval conidia [24,25].
Members of Savoryellaceae are often found in freshwater habitats [17,23,26]. For example, the genus Canalisporium is found in freshwater habitats and typified by C. caribense [27]. Most Canalisporium species are characterized by dark brown and muriform conidia [27]; Canalisporium grenadoideum (=Ascothailandia grenadoidea) is the only species that represents a sexual morph: globose, dark brown, ostiolate ascomata, eight-spored, long cylindrical, unitunicate asci, fusiform, three-euseptate ascospores [28]. Dematiosporium, typified by D. aquaticum, and currently consist of species, in particular, from freshwater habitat [17,26]. This genus is characterized by cylindrical, unbranched, aseptate conidiophores and globose to subglobose, dictyospores conidia with a pore in each cell [17,26]. The generic type of Savoryella, S. lignicola, was initially discovered during a survey of cellulolytic fungi in a simulated aquatic environment and has been subsequently recovered from different woody substrates (i.e., Bambusa sp.) in aquatic or marine habitats [18,21,29]. Members of Savoryella are characterized by immersed, partly immersed, or superficial, globose, subglobose, or ellipsoidal ascostromata, two-to-eight-spored, cylindrical or clavate, unitunicate asci with an apical truncate non-amyloid apical thickening containing a pore and uni- or biseriate, ellipsoidal, three-septate ascospores [18,30]; as well as dematiaceous hyphomycetes, which produce micronematous conidiophores, holoblastic, terminal, and intercalary conidiogenous cells, solitary or aggregated, pyriform to obovoid, septate conidia [19].
During a survey of saprobic fungi from bamboo in Sichuan Province, China, a series of asexual and sexual fungi were collected. Multi-gene phylogeny integrated with morphological comparison was carried out to determine the taxonomic status of these new collections, of which new species and new host records contributed to Canalisporium, Dematiosporium, and Savoryella were introduced and justified to the Savoryellaceae.
2. Materials and Methods
2.1. Isolation and Morphological Examination
Fungi associated with decaying bamboo were collected from Dujiangyan and Qionglai in Sichuan Province, China in 2021. Specimens were placed in envelopes or plastic bags and taken to the laboratory. Morphological characteristics were observed using a Nikon ECLIPSE E200 stereo microscope and photographed by a Nikon ECLIPSE Ni-U compound microscope fitted with a Nikon DS-Ri2 digital camera as per the guidelines provided in Senanayake et al. [31]. Single-spore isolations were used to obtain pure cultures following the methods described by Senanayake et al. [31]. Measurements were made with the Tarosoft (R) Image Framework program v. 0.9.7, following Liu et al. [32]. Photo plates representing fungal structures were processed in Adobe Photoshop CS6 software (Adobe Systems Inc., San Jose, CA, USA). Herbarium specimens (dry branches with fungal material) were deposited in the herbarium of Cryptogams, Kunming Institute of Botany Academia Sinica (KUN-HKAS), Kunming, China and the herbarium of the University of Electronic Science and Technology (HUEST), Chengdu, China. The isolates obtained in this study were deposited in China General Microbiological Culture Collection Center (CGMCC) in Beijing, China and the University of Electronic Science and Technology Culture Collection (UESTCC) in Chengdu, China. Names of the new taxa were registered in MycoBank [33].
2.2. DNA Extraction, PCR Amplification and Sequencing
A Trelief TM Plant Genomic DNA Kit (Beijing TsingKe Biotech Co., Ltd., Beijing, China) was used to extract total genomic DNA from fungal mycelia. DNA amplification was performed by a polymerase chain reaction (PCR). Five partial gene regions, the small subunit of nuclear ribosomal RNA gene (SSU), the nuclear ribosomal internal transcribed spacer (ITS: ITS1-5.8S-ITS2), the large subunit of nuclear ribosomal RNA gene (LSU), the second-largest subunit of RNA polymerase II (rpb2), and the translation elongation factor 1-alpha (tef1α), were used in this study. Sequences of SSU, ITS, LSU, rpb2, and tef1α were amplified using primer pairs NS1/NS4, ITS5/ITS4, LR0R/LR5, fRPB2-5F/fRPB2-7cR, and 983F/2218R, respectively [34,35,36,37]. The amplification reactions were performed in 50 μL PCR mixtures containing 25 μL 2× Taq Plus MasterMix (Dye) (CoWin Biosciences, (Taizhou), Co., Ltd., Taizhou, China), 20 μL ddH2O, 1μL DNA template, and 2μL of each primer (10µM/L). The PCR thermal cycle program for SSU, ITS, LSU, rpb2, and tef1α amplification was as follows: initial denaturing step of 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 56 °C (SSU, ITS, LSU, rpb2), 62 °C (tef1α) for 30 s, elongation at 72 °C for 30 s, and final extension at 72 °C for 5 min. PCR products were checked on 1% agarose electrophoresis gels stained with Gel Red. The sequencing reactions were carried out with primers mentioned above by Beijing Tsingke Biotechnology Co., Ltd., Chengdu, China.
2.3. Phylogenetic Analyses
The BLAST searches were performed for finding similar sequences that match our data. A concatenated dataset of the SSU, ITS, LSU, rpb2, and tef1α sequences were used for phylogenetic analyses with the inclusion of reference taxa from GenBank (Table 1). The sequences were aligned by using the online multiple alignment program MAFFT v.7 (http://mafft.cbrc.jp/alignment/server/ (accessed on 27 May 2022)) [38], and the alignment was manually optimized in BioEdit v.7.0.9 [39]. The five-gene dataset was concatenated by Mesquite v. 3.11 (http://www.mesquiteproject.org/ (accessed on 27 May 2022)) for multi-gene phylogenetic analyses. Maximum likelihood (ML) and Bayesian inference (BI) were carried out as detailed in Dissanayake et al. [40]. Maximum likelihood (ML) analysis was performed using RAxML-HPC v.8 tool via the CIPRES Science Gateway V3.3 (https://www.phylo.org/portal2/home.action, accessed on 27 May 2022) with rapid bootstrap analysis, and a general time-reversible model (GTR) was applied with a discrete GAMMA distribution. The Bayesian inference (BI) analyses were performed by using PAUP v.4.0b10 [41] and MrBayes v. 3.1.2 [42,43]. The best model for different genes partition in the concatenated dataset was determined by MrModeltest 2.3. [44], and posterior probabilities were determined by Markov Chain Monte Carlo sampling (MCMC) in MrBayes v.3.1.2 [42,43]. The final alignment and phylogram were submitted to TreeBASE (https://www.treebase.org/, accessed on 5 May 2023, submission ID: 30350). The phylogenetic trees were visualized by Treeview v. 1.6.6 [45].

Table 1.
Taxa used in the phylogenetic analyses and their GenBank accession numbers. Newly generated sequences are indicated with * and the ex-type strains are in bold. “N/A” sequence is unavailable.
3. Results
3.1. Phylogenetic Analyses
To determine the phylogenetic placement of the new collections in this study, the combined SSU, ITS, LSU, rpb2, and tef1α data set comprised 55 sequences with Pleurotheciella aquatica (MFLUCC 17-0464) and P. erumpens (CBS 142447) as the outgroup taxa. The concatenated matrix comprised a total of 4870 characters (SSU: 1081 bp; ITS: 841 bp; LSU: 921 bp; rpb2: 1056 bp; tef1α: 971 bp) including gaps. Maximum likelihood (ML) and Bayesian inference (BI) analyses were resulting in generally congruent topologies. The best-scoring ML tree (Figure 1) was selected to represent the relationships among taxa, in which a final likelihood value of –37,950.559206 is presented. The evolutionary models for Bayesian analysis were selected for each locus, and the best-fit model GTR+I+G for ITS, LSU, rpb2, and tef1α, SYM+I+G for SSU, respectively. Six simultaneous Markov chains were run for 165,000 generations, and trees were sampled every 1000 generations and 165 trees were obtained. The first 33 trees representing the burn-in phase of the analyses were discarded, while the remaining 132 trees were used for calculating posterior probabilities in the majority rule consensus tree (critical value for the topological convergence diagnostic is 0.01).
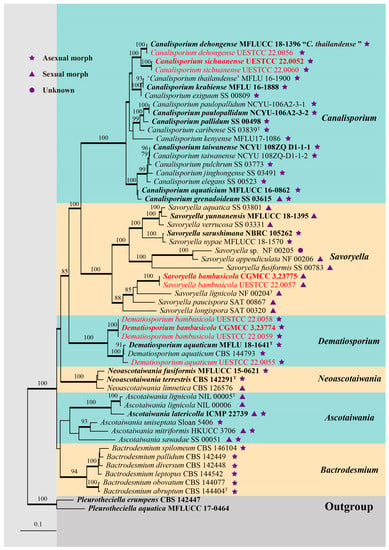
Figure 1.
RAxML tree generated from combined SSU, ITS, LSU, rpb2, and tef1α sequence data of Savoryellaceae. Bootstrap values for ML equal to or greater than 75% are placed above the branches. Branches with Bayesian posterior probabilities (BYPP) from MCMC analysis equal to or greater than 0.95 were in bold. The ex-type strains were indicated in bold, and newly generated sequences were indicated in red. “T” represents the type species of each genus.
Forty-three representative species of Savoryellaceae are included in our phylogenetic analysis (Figure 1). Two isolates of Canalisporium sichuanense (CGMCC 3.23926, UESTCC 22.0060) were sister to C. dehongense (MFLUCC 18-1396 and UESTCC 22.0056) and were well supported (100% ML/1.00 BYPP). Three isolates of Dematiosporium bambusicola (CGMCC 3.23774, UESTCC 22.0058, UESTCC 22.0059) formed a distinct clade sister to Dematiosporium aquaticum (CBS 144793, MFLU 18-1641, UESTCC 22.0055) with high statistical support (100% ML/1.00 BYPP). Two isolates of Savoryella bambusicola (CGMCC 3.23775 and UESTCC 22.0057) were nested in the genus Savoryella and close to S. lignicola (NF 00204).
3.2. Taxonomy
Canalisporium dehongense W. Dong, H. Zhang, and K.D. Hyde, Fungal Diversity 96: 159 (2019) Figure 2 and Figure 3

Figure 2.
Canalisporium dehongense (HUEST 22.0057) (a–c) colonies on bamboo substrate. (d–g) Conidiophores and conidia. (h–l) Conidia with conidiogenous cells. (m,n) Colonies on PDA, above (m) and below (n). Scale bars: (d–l) = 10 μm.
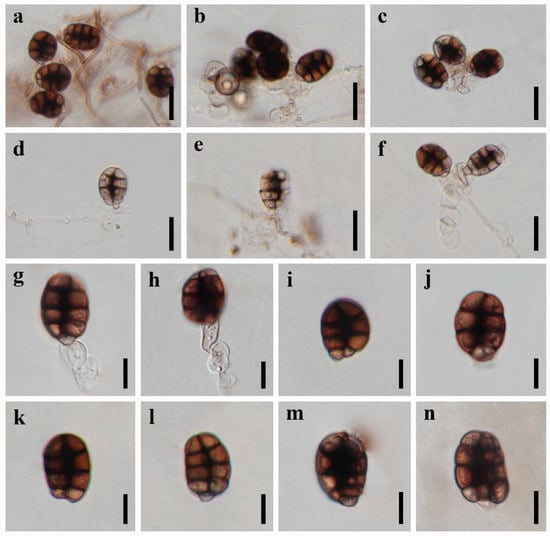
Figure 3.
Asexual reproduction of Canalisporium dehongense (UESTCC 22.0056) on PDA medium about three months. (a) Hyphae and conidiophores with conidia. (b–h) Conidiogenous cells and conidia. (i–n) Conidia. Scale bars: (a–f) = 20 μm, (g–n) = 10 μm.
MycoBank: MB 555407
Saprobic on dead branches of bamboo in terrestrial habitat. Sexual morph: Undetermined. Asexual morph: hyphomycetous (Figure 2 and Figure 3). Colonies sporodochial, scattered, punctiform, pulvinate, granular, black, glistening. Mycelium immersed, consisting of branched, septate, thin-walled, smooth, pale to brown hyphae. Conidiophores up to 60 μm long, micronematous, mononematous, vesiculate, consisted of one to five subglobose to cylindrical, hyaline cells, smooth, unbranched, septate, constricted at the septa. Conidiogenous cells 8.5–12.5 × 7–10 µm ( = 9.8 × 8.8 µm, n = 10), holoblastic, monoblastic, integrated, terminal, determinate, subglobose, ellipsoidal, sometimes cuneiform, hyaline, smooth, thin-walled. Conidia 25–32 × 17–23 µm ( = 29 × 20 µm, n = 50), solitary, acrogenous, ellipsoidal to obovoid, muriform, smooth, brown, comprising of one straight column of vertical septa and three to four rows of transverse septa, slightly constricted at the septa, darkened and thickly banded at the septa, canals in the septa obscured by dark pigmentation; basal cell single, cuneiform, 4.5–6.5 µm wide, sometimes swollen, hyaline to pale brown.
Culture characteristics: Colonies on PDA reaching 20–30 mm after 5 months at 25 °C, circular, raised to umbonate, rough surface, dense, entire at the edge, brown, dry, reverse dark brown to black. Mycelium subhyaline to pale brown, 1.5–2.5 µm wide in culture. Conidiophores subhyaline. Conidiogenous cells integrated, subhyaline. Conidia pale brown to brown, septate, subglobose to obovoid, muriform, smooth, constricted at the septa, 20–25 µm ( = 22.5 µm, n = 30) long, 15–17 µm ( = 16.5 µm, n = 30) wide (Figure 3).
Material examined: CHINA, Sichuan Province, Chengdu City, Qionglai County, Chuanxi Bamboo Sea Area, 30°20′32″ N, 103°18′26″ E, 540 m elevation, on dead branches of bamboo in terrestrial habitat, 12 October 2021, X.D. Yu, A5 (HUEST 22.0057); living culture UESTCC 22.0056.
Notes: Canalisporium dehongense was introduced by Hyde et al. [46] from a submerged wood in Yunnan Province, China and has been considered as a later synonym of C. thailandense [47]. However, Goh and Kuo [48] treated Canalisporium dehongense and C. thailandense as distinct species, and this is followed in this study. The new collection clustered with Canalisporium dehongense in the single- and multi-gene phylogeny and share the similar conidiogenous cells and conidial morphology. Therefore, we identify our collection as C. dehongense, and an additional bamboo host record is provided herein.

Figure 4.
Canalisporium sichuanense (HKAS 124625, holotype) (a–h) colonies on natural substrate. (a,b) Sporodochia on a bamboo substrate. (c) Squash mount of sporodochium showing conidiophores and conidia. (d) Conidiophores comprising of vesiculate cells. (e–h) Conidia with conidiogenous cells. (i,j) Colonies on PDA, above (i) and below (j). Scale bars: (c) = 50 μm, (d) = 20 μm, (e–h) = 10 μm.

Figure 5.
Asexual reproduction of Canalisporium sichuanense (CGMCC 3.23926, ex-type) on PDA medium about three months. (a,b) Hyphae and conidiophores with conidia. (c–i) Conidiogenous cells and conidia. Scale bars: (a,b) = 50 μm, (c–i) = 10 μm.
MycoBank: MB 847552
Etymology: The epithet refers to Sichuan Province where the fungus was collected.
Holotype: HKAS 124625
Saprobic on dead branches of bamboo in terrestrial habitat. Sexual morph: Undetermined. Asexual morph: hyphomycetous (Figure 4 and Figure 5). Colonies sporodochial, scattered, punctiform, pulvinate, granular, black, and shiny. Mycelium immersed, consisting of branched, septate, thin-walled, smooth, hyaline to pale brown hyphae. Conidiophores up to 60 µm long, micronematous, mononematous, vesiculate, consisted of one to six subglobose to cylindrical, hyaline cells, smooth, unbranched, septate, constricted at the septa. Conidiogenous cells 11–16 × 8–10 µm ( = 13.5 × 9 µm, n = 10), holoblastic, monoblastic, integrated, determinate, terminal, cylindrical, sometimes swelling to subglobose, hyaline, smooth, thin-walled. Conidia 27–37 × 17–21 µm ( = 31 × 20 µm, n = 50), solitary, acrogenous, cylindrical to obovoid, muriform, smooth, brown, comprising of a single, straight column of vertical septa and two to four rows of transverse septa, slightly constricted at the septa, darkened and thickly banded at the septa, canals in the septa obscured by dark pigmentation; basal cell cuneiform, 5.0–7.0 µm wide, sometimes swollen, hyaline to pale brown.
Culture characteristics: Colonies on PDA reaching 5–10 mm after 3 months at 25 °C, irregular, raised to umbonate, surface rough, dense, greyish-white, dry, reverse dark brown. Mycelium subhyaline to pale brown, 1.5–2.5 µm wide in culture. Conidiophores (Figure 5) subhyaline, 10–35 × 3.5–10.5 µm ( = 20.0 × 6.5 µm, n = 10). Conidiogenous cells (Figure 5) integrated, subhyaline, 4.5–10.5 × 5.5–10.5 µm ( = 8 × 7.5 µm, n = 10). Conidia (Figure 5) pale brown to brown, septate, cylindrical to obovoid, muriform, smooth, constricted at the septa, and the septa becoming progressively darker with conidial maturity, 15–24 µm ( = 20.5 µm, n = 30) long × 9–15 µm ( = 12.0 µm, n = 30) wide.
Material examined: CHINA, Sichuan Province, Chengdu City, Qionglai County, Lugou Bamboo Sea Area, 30°22′37″ N, 103°16′45″ E, 730 m elevation, on dead branches of bamboo in a terrestrial habitat, 12 October 2021, X.D. Yu, A44 (HKAS 124625, holotype); ex-type living culture CGMCC 3.23926; Dujiangyan County, Qingcheng Mountain, 30°54′29″ N, 103°32′36″ E, 960 m elevation, on dead branches of bamboo in a terrestrial habitat, 02 December 2021, Y. Yang, Q10-4 (HUEST 22.0061, paratype); ex-paratype living culture UESTCC 22.0060.
Notes: Morphologically, Canalisporium sichuanense resembles C. dehongense and C. thailandense in having hyaline, septate conidiophores, holoblastic, monoblastic conidiogenous cells, and muriform conidia [46,49]. However, they can be distinguished by the size of the basal cell (ca. 4 μm long in C. sichuanense, ca. 5 μm long in C. dehongense, ca. 2 μm long in C. thailandense) and conidial size (27–37 × 17–21 μm in C. sichuanense, 20–30 × 12–19 μm in C. dehongense, 22.5–31 × 17–22 μm in C. thailandense) [46,49]. Phylogenetically, they are distinct from each other (Figure 1), of which C. sichuanense is closer and sister to C. dehongense with high statistical support (100% ML/1.00 BYPP). The comparison of nucleotide differences between C. sichuanense and C. dehongense (MFLUCC 18-1396) showed that there are 0.10% (1/970) differences in SSU, 6.69% (44/658, including six gaps) in ITS, and 1.45% (11/761) in LSU gene regions, which also supports the introduction of C. sichuanense as a new species.
Dematiosporium aquaticum Z.L. Luo, K.D. Hyde, and H.Y. Su, Fungal Diversity 99: 573 (2019) Figure 6
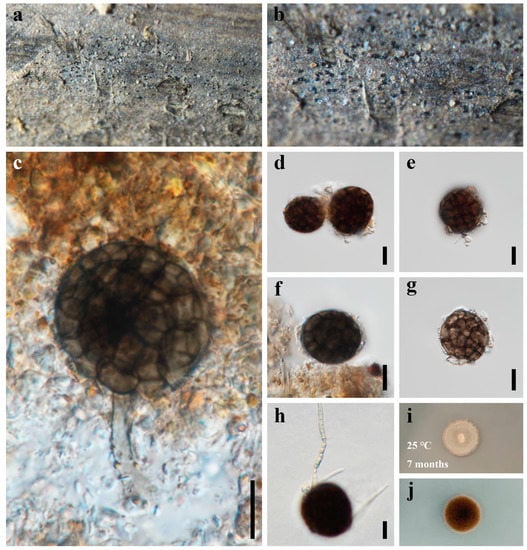
Figure 6.
Dematiosporium aquaticum (HUEST 22.0056) (a,b) colonies on bamboo substrate. (c) Conidiophores and conidia. (d–f) Conidia. (g) Fragile conidium. (h) Germinating conidium. (i,j) Colonies on PDA, above (i) and below (j). Scale bars: (c–h) = 10 μm.
MycoBank: MB 555673
Saprobic on dead bamboo in freshwater habitat. Sexual morph: Undetermined. Asexual morph: hyphomycetous. Colonies on natural substratum, effuse, scattered to gregarious, dark brown to black, glistening, superficial. Mycelium immersed in natural substrate, unbranched, septate, hyaline to subhyaline hyphae. Conidiophores reduced to conidiogenous cells, or rarely 20–30 × 3.5–4.5 µm, micronematous, mononematous, cylindrical, unbranched. Conidiogenous cells not observed. Conidia 21–28 µm diam ( = 25 µm, n = 50), solitary, acrogenous, globose to subglobose, brown to dark brown, dictyospores, slightly constricted at the septa, with a pore in each cell.
Culture characteristics: Colonies on PDA reaching 10–15 mm after 7 months at 25 °C, circular, with dense mycelium on the surface, dark grayish of the inner ring, and brown.
Material examined: CHINA, Sichuan Province, Chengdu City, Qionglai County, Lugou Bamboo Sea Area, 30°22′37″ N, 103°16′45″ E, 730 m elevation, on dead branches of bamboo in a freshwater habitat, 12 October 2021, X.D. Yu, A1 (HUEST 22.0056); living culture UESTCC 22.0055.
Notes: Luo et al. [26] introduced a monotypic genus Dematiosporium, which was collected from decaying submerged wood in Erhai lake, Dali City, Yunnan Province, China. Subsequently, Réblová et al. [17] recollected and described it in France, where it occurs on decaying submerged wood of Alnus glutinosa and other unidentified substrates. Species of Dematiosporium are often found on decaying wood in aquatic environments and it is characterized by cylindrical, unbranched, aseptate conidiophores and globose to subglobose, dictyospores conidia with a pore in each cell [17,26]. Our new collection was found from decaying bamboo in freshwater environments. We identify it as D. aquaticum based on the morphological characters and phylogenetic analyses. The SSU, ITS, rpb2, and tef1α regions were attempted, and we were able to obtain the LSU sequences. This is the first report of D. aquaticum from bamboo in a freshwater habitat. We describe D. aquaticum in detail and supplement the description of the conidiophore that was missing from the previous description.
Dematiosporium bambusicola X.D. Yu, S.N. Zhang, and Jian K. Liu, sp. nov., Figure 7.

Figure 7.
Dematiosporium bambusicola (HKAS 124626, holotype) (a,b) colonies on a bamboo substrate. (c–e) Conidiophores and conidia. (f–j) Conidia. (j) The arrow indicated pore. (k) Germinating conidium. (l,m) Colonies on PDA, above (l) and below (m). Scale bars: (c–k) = 20 μm.
MycoBank: MB 847553
Etymology: Refers to the bamboo host from which the fungi was found.
Holotype: HKAS 124626
Saprobic on dead bamboo in freshwater habitat. Sexual morph: Undetermined. Asexual morph: hyphomycetous. Colonies on natural substratum, effuse, scattered to gregarious, dark brown to black, glistening, superficial. Mycelium immersed in natural substrate, composed of hyaline, septate, branched, smooth hyphae. Conidiophores 6.5–11.5 × 2–4.5 µm ( = 9 × 3 µm, n = 10), micronematous, mononematous, cylindrical, straight or slightly flexuous, unbranched, aseptate. Conidiogenous cells holoblastic, monoblastic, cuneiform, integrated, terminal, determinate, hyaline. Conidia 35–60 × 27–41 µm ( = 46 × 34 µm, n = 50), solitary, acrogenous, ellipsoidal to subglobose, dark brown to black, smooth-walled, dictyospores, with a pore in each cell (Figure 7j), broadly rounded at apex, subtruncate at the base.
Culture characteristics: Colonies on PDA reaching 30–40 mm after 4 months at 25℃, circular, with sparse mycelium on the surface, slightly raised, umbonate, greyish-green, and reverse dark brown to black.
Material examined: CHINA, Sichuan Province, Chengdu City, Qionglai County, Lugou Bamboo Sea Area, 30°22′37″ N, 103°16′45″ E, 730m elevation, on dead branches of bamboo in freshwater habitat, 12 October 2021, X.D. Yu, A22 (HKAS 124626, holotype); ex-type living culture CGMCC 3.23774, living culture UESTCC 22.0058, UESTCC 22.0059.
Notes: The phylogenetic analyses showed that three isolates of Dematiosporium bambusicola formed a monophyletic clade in Savoryellaceae and are sister to D. aquaticum with absolute statistical support (100% ML/1.00 BYPP). Dematiosporium Bambusicola resembles D. aquaticum in forming dematiaceous, dictyospores conidia with a pore [26]. However, D. bambusicola has larger conidia than that of D. aquaticus (35–60 × 27–41 µm vs. 21–28 µm diam) [26]. The conidia of D. bambusicola are ellipsoidal to subglobose and dark brown to nearly black, while D. aquaticus has globose to subglobose conidia and brown to dark brown [26]. In addition, D. bambusicola differs from the latter in having smaller conidiophores (6.5–11.5 × 2–4.5 µm vs. 20–30 × 3.5–4.5 µm) [26]. We hereby introduce the new species based on the distinctiveness of morphology and multi-gene phylogeny.
Savoryella bambusicola X.D. Yu, S.N. Zhang, and Jian K. Liu, sp. nov., Figure 8.

Figure 8.
Savoryella bambusicola (HKAS 124627, holotype) (a,b) ascomata on host substrate. (c) Neck. (d) Squash mounts of ascomata. (e,f) Peridium. (g) Paraphyses. (h) Apical ring. (i–l) Asci. (m–p) Ascospores. (q,r) Colonies on PDA, above (q) and below (r). Scale bars: (c) = 50 μm, (d) = 100 μm, (e–g,i–l) = 20 μm, (m–p) = 10 μm, (h) = 5 μm.
MycoBank: MB 847554
Etymology: Refers to the bamboo host from which the fungi was found.
Holotype: HKAS 124627
Saprobic on dead bamboo in freshwater habitat. Sexual morph: Ascomata 270–350 μm high, 180–210 μm diam., scattered or solitary, superficial, black, ellipsoidal, uniloculate, thin-walled, laterally ostiolate, lying horizontal to the host surface with a short, brown neck. Peridium comprising several layers of brown, thick-walled cells of textura epidermoidea. Paraphyses 4.5–6.5 µm wide, numerous, cylindrical, and somehow swollen, branched, hyaline, septate, constricted at the septa. Asci 90–130 × 19–23 µm ( = 119 × 21 µm, n = 30), 8-spored, unitunicate, cylindric-clavate, straight or slightly curved, short pedicellate, rounded at the apex, with an apical truncate non-amyloid apical thickening containing a pore. Ascospores 24.5–36.5 × 9–12 µm ( = 30.5 × 10.5 µm, n = 50), uni- or biseriate, straight or slightly curved, fusiform, three-septate, constricted and thickened at the septa, central cells larger and brown, apical cells smaller and hyaline, thin-walled, without sheaths or appendages. Asexual morph: Undetermined.
Culture characteristics: Colonies on PDA reaching 10–15 mm after 2 months at 25 °C, circular, with dense mycelium on the surface, light gray of the inner ring, and light yellow of the outer ring; in reverse black.
Material examined: CHINA, Sichuan Province, Chengdu City, Qionglai County, Lugou Bamboo Sea Area, 30°22′37″ N, 103°16′45″ E, 730m elevation, on dead branches of bamboo in freshwater habitat, 12 October 2021, X.D. Yu, A23 (HKAS 124627, holotype); ex-type living culture CGMCC 3.23775, ibid., A20 (HUEST 22.0058, isotype); ex-isotype living culture UESTCC 22.0057.
Notes: The phylogenetic result based on SSU, ITS, LSU, rpb2, and tef1α sequence data showed that the new collections Savoryella bambusicola nested in Savoryella and formed a distinct lineage (Figure 1). Morphologically, S. bambusicola resembles S. curvispora and S. fusiformis in having eight-spored asci, fusiform, three-septate ascospores [29]. However, they can be recognized as different species; Savoryella bambusicola differs from S. curvispora and S. fusiformis in having relatively broader asci (90–130 × 19–23 vs. 90–115 × 15–17 vs. 80–120 × 9–14) and ascospores (24.5–36.5 × 9–12 vs. 25–28 × 7–10 vs. 25–35 × 6–9.6) [29]. The establishment of the new species S. bambusicola is justified by morphological and phylogenetic evidence.
4. Discussion
Tibpromma et al. [20] introduced Canalisporium krabiense and C. thailandense from Thailand. These two species have different conidiogenous cells, but were found to have almost identical ITS and LSU sequences and were phylogenetically clustered together (Figure 1). Koukol and Delgado [47] speculated that DNA cross-contamination happened between C. krabiense and C. thailandense. Furthermore, they provided synonymy of another species C. dehongense under C. thailandense. In this study, we deem it as a speculative inference and do not follow this treatment with the following concerns: (1) in Koukol and Delgado [47], the globose to oval conidiogenous cells connected in a chain of C. dehongense and C. thailandense was also observed in other incorrectly identified C. caribense; and inferring from this, they speculated that C. thailandense is worldwide and C. dehongense could be a further record; (2) the synonymy of C. dehongense under C. thailandense is only based on morphology; since C. dehongense and C. thailandense are represented by a single specimen at that time, there was no further evidence to refute the inference [20,46,47]; (3) additional collection of C. dehongense is provided in this study (HUEST 22.0057; UESTCC 22.0056), which supported and validated the initial identification of C. dehongense in Hyde et al. [46]; (4) Goh and Kuo [48] also pointed out that C. dehongense and C. thailandense (the basal cell ca. 5µm long vs. 2µm long) are distinct species due to morpho-phylogenetical distinction.
Species of Savoryellaceae have been found on various hosts in aquatic and terrestrial habitats [16], i.e., Ascotaiwania mauritiana on Pandanus palustris (Pandanaceae) [50], Ascotaiwania palmicola on Iriartea sp. (Arecaceae) [51], Savoryella aquatica on Machilus sp. (Lauraceae) [21], Savoryella lignicola on Bambusa sp. (Poaceae) [21], Savoryella nypae on Nypa fruticans (Arecaceae) [19], Savoryella paucispora on Pinus sp. (Pinaceae) [22]. Freshwater species are accommodated in all the genera in Savoryellaceae: Ascotaiwania (6 species) [50,52,53,54], Bactrodesmium (6 species) [17,55], Canalisporium (17 species) [27,28,46,48,56,57,58,59,60], Dematiosporium (1 species) [26], Neoascotaiwania (4 species) [23,52,61,62], and Savoryella (9 species) [16,19,21,29,63]. It seems the members of this family are more likely to be found from monocotyledons and favorable to hard tissue substrates in freshwater habitats.
Asexual morphs have been found in all six genera in Savoryellaceae, of which Bactrodesmium and Dematiosporium are represented only by asexual morphs; members of Canalisporium mostly are asexual morphs with only C. grenadoideum having a holomorph; while Savoryella is commonly known as a sexual morph with only S. nypae and S. sarushimana represented by a trichocladium-like asexual morph [19]; Ascotaiwania and Neoascotaiwania both have holomorphs, but different in asexual morphs (monodictys-like, monotosporella-like, and trichocladium-like vs. bactrodesmium-like) [53,62,64,65,66]. In this study, one sexual morph of Savoryella and four asexual morphs of Canalisporium and Dematiosporium are isolated and identified, which contributed to the taxonomy of Savoryellaceae and the diversity of bambusicolous fungi.
Author Contributions
Conceptualization, X.-D.Y., S.-N.Z. and J.-K.L.; methodology, X.-D.Y. and J.-K.L.; formal analysis, X.-D.Y. and S.-N.Z.; resources, X.-D.Y. and J.-K.L.; data curation, X.-D.Y. and S.-N.Z.; writing—original draft preparation, X.-D.Y.; writing—review and editing, X.-D.Y., S.-N.Z. and J.-K.L.; supervision, J.-K.L.; project administration, J.-K.L.; funding acquisition, J.-K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study is supported by the Joint Fund of the National Natural Science Foundation of China and the Karst Science Research Center of Guizhou Province (Grant No. U1812401).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequences and alignments data were submitted to GenBank and TreeBASE, respectively.
Acknowledgments
Xiang Ma, Yi Yang and Jing-Yi Song are thanked for their help with sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Phookamsak, R.; Jiang, H.; Suwannarach, N.; Lumyong, S.; Xu, J.; Xu, S.; Liao, C.F.; Chomnunti, P. Bambusicolous Fungi in Pleosporales: Introducing Four Novel Taxa and a New Habitat Record for Anastomitrabeculia didymospora. J. Fungi 2022, 8, 630. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Shukla, A.; Singh, A.; Tiwari, D.; Ahirwar, B.K. Bambusicolous fungi: A reviewed documentation. Int. J. Pure Appl. Biosci. 2016, 4, 304–310. [Google Scholar] [CrossRef]
- Hyde, K.D.; Zhou, D.Q.; Dalisay, T. Bambusicolous fungi: A review. Fungal Divers. 2002, 9, 1–14. [Google Scholar]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Hatakeyama, S.; Harada, Y.; Sano, T.; Shirouzu, T.; Hosoya, T. Molecular taxonomy of bambusicolous fungi: Tetraplosphaeriaceae, a new pleosporalean family with Tetraploa-like anamorphs. Stud. Mycol. 2009, 64, 175–209. [Google Scholar] [CrossRef]
- Dai, D.Q.; Tang, L.Z.; Wang, H.B. A Review of Bambusicolous Ascomycetes. Bamboo Curr. Future Prospect. 2018, 165, 10. [Google Scholar] [CrossRef]
- Shi, J.Y.; Zhou, D.Q.; Ma, L.S.; Yao, J.; Zhang, D.M. Diversity of Bamboo Species in China. World Bamboo Ratt. 2020, 18, 55–65. [Google Scholar] [CrossRef]
- Jiang, H.; Phookamsak, R.; Hongsanan, S.; Bhat, D.J.; Mortimer, P.E.; Suwannarach, N.; Kakumyan, P.; Xu, J. A review of bambusicolous Ascomycota in China with an emphasis on species richness in southwest China. Stud. Fungi 2022, 7, 1–33. [Google Scholar] [CrossRef]
- Yu, X.D.; Zhang, S.N.; Liu, J.K. Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China. J. Fungi 2022, 8, 720. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. [Google Scholar]
- Yang, C.L.; Xu, X.L.; Liu, Y.G. Two new species of Bambusicola (Bambusicolaceae, Pleosporales) on Phyllostachys heteroclada from Sichuan, China. Nova Hedwig. 2019, 108, 527–545. [Google Scholar] [CrossRef]
- Zeng, Q.; Lv, Y.C.; Xu, X.L.; Deng, Y.; Wang, F.H.; Liu, S.Y.; Liu, L.J.; Yang, C.L.; Liu, Y.G. Morpho-Molecular Characterization of Microfungi Associated with Phyllostachys (Poaceae) in Sichuan, China. J. Fungi 2022, 8, 702. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Dong, W.; Wanasinghe, D.N.; Liu, Y.G.; Hyde, K.D. Introducing Arthrinium phyllostachium sp. nov.(Apiosporaceae, Xylariales) on Phyllostachys heteroclada from Sichuan province, China. Phytotaxa 2019, 406, 91–110. [Google Scholar] [CrossRef]
- Li, L.; Guan, M.; Liu, H.; Zhu, T.; Li, S.; Han, S.; Lin, T.; Qiao, T.; Liu, Y.; Yang, C. Fusarium proliferatum Associated with Basal Rot Disease of Bambusa pervariabilis × Dendrocalamopsis grandis in China. Plant Dis. 2022, 6, 2531. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Xu, X.; Liu, Y.; Hyde, K.D.; Mckenzie, E.H. A new species of Phyllachora (Phyllachoraceae, Phyllachorales) on Phyllostachys heteroclada from Sichuan, China. Phytotaxa 2019, 392, 186–196. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; Dong, W.; Devadatha, B.; Yang, J.; Ekanayaka, A.H.; De Silva, W.; Sarma, V.V.; Al-Sadi, A.M.; et al. Phylogenetic Revision of Savoryellaceae and Evidence for Its Ranking as a Subclass. Front. Microbiol. 2019, 10, 840. [Google Scholar] [CrossRef] [PubMed]
- Réblová, M.; Hernández-Restrepo, M.; Fournier, J.; Nekvindová, J. New insights into the systematics of Bactrodesmium and its allies and introducing new genera, species and morphological patterns in the Pleurotheciales and Savoryellales (Sordariomycetes). Stud. Mycol. 2020, 95, 415–466. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G.; Eaton, R. Savoryella lignicola gen. et sp. nov. from water-cooling towers. Trans. Br. Mycol. Soc. 1969, 52, 161–165. [Google Scholar] [CrossRef]
- Zhang, S.N.; Abdel-Wahab, M.A.; Jones, E.B.G.; Hyde, K.D.; Liu, J.K. Additions to the genus Savoryella (Savoryellaceae), with the asexual morphs Savoryella nypae comb. nov. and S. sarushimana sp. nov. Phytotaxa 2019, 408, 195–207. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Hyde, K.D. Tropical Australian freshwater fungi. V.* Bombardia sp., Jahnula australiensis sp. nov., Savoryella aquatica sp. nov. and S. lignicola sp. nov. Aust. Syst. Bot. 1993, 6, 161–167. [Google Scholar] [CrossRef]
- Manimohan, P.; Amritha, M.; Sairabanu, N.K. A comparison of diversity of marine fungi on three co-habiting mangrove plants. Mycosphere 2011, 2, 533–538. [Google Scholar] [CrossRef]
- Yang, L.; Bao, D.F.; Luo, Z.L.; Su, X.J.; Su, H.Y. Neoascotaiwania aquatica sp. nov. from a freshwater habitat in Yunnan Province, China. Phytotaxa 2022, 531, 120–128. [Google Scholar] [CrossRef]
- Jaklitsch, W. Savoryellaceae Jaklitsch & Réblová. Index Fungorum 2015, 209, 1. [Google Scholar]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef]
- Nawawi, A.; Kuthubutheen, A.J. Canalisporium, a new genus of lignicolous hyphomycetes from Malaysia. Mycotaxon 1989, 34, 475–487. [Google Scholar]
- Sri-Indrasutdhi, V.; Boonyuen, N.; Suetrong, S.; Chuaseeharonnachai, C.; Sivichai, S.; Jones, E.B.G. Wood-inhabiting freshwater fungi from Thailand: Ascothailandia grenadoidia gen. et sp nov., Canalisporium grenadoidia sp nov with a key to Canalisporium species (Sordariomycetes, Ascomycota). Mycoscience 2010, 51, 411–420. [Google Scholar] [CrossRef]
- Ho, W.H.; Hyde, K.D.; Hodgkiss, I.J. Ascomycetes from tropical freshwater habitats: The genus Savoryella, with two new species. Mycol. Res. 1997, 101, 803–809. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Senanayake, I.; Rathnayake, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E.; Jones, E.B.G.; Moslem, M.; Hyde, K.D. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Kazutaka, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; pp. 95–98. [Google Scholar]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.M.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Nylander, J.; Zoology, S.; Posada, D.; Mrmodeltest, R.; Os, F. MrModeltest2 v. 2.3 (Program for Selecting DNA Substitution Models Using PAUP*); Evolutionary Biology Centre: Uppsala, Sweden, 2008. [Google Scholar]
- Page, R.D.M. Tree View: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Koukol, O.; Delgado, G. Why morphology matters: The negative consequences of hasty descriptions of putative novelties in asexual ascomycetes. Ima Fungus 2021, 12, 1–8. [Google Scholar] [CrossRef]
- Goh, T.K.; Kuo, C.H. Reflections on Canalisporium, with descriptions of new species and records from Taiwan. Mycol. Prog. 2021, 20, 647–680. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Dulymamode, R.; Cannon, P.F.; Hyde, K.D.; Peerally, A. Four new ascomycete species from endemic Pandanus of Mauritius. Fungal Divers. 2001, 8, 87–96. [Google Scholar]
- Hyde, K. Fungi on palms XXV. A new species of Ascotaiwania. Sydowia 1995, 47, 31–37. [Google Scholar]
- Chang, H.S.; Hsieh, S.Y.; Jones, E.B.G.; Read, S.J.; Moss, S.T. New freshwater species of Ascotaiwania and Savoryella from Taiwan. Mycol. Res. 1998, 102, 709–718. [Google Scholar] [CrossRef]
- Ranghoo, V.M.; Hyde, K.D. Ascomycetes from freshwater habitats: Ascolacicola aquatica gen. et sp. nov. and a new species of Ascotaiwania from wood submerged in a reservoir in Hong Kong. Mycologia 1998, 90, 1055–1062. [Google Scholar] [CrossRef]
- Hyde, K.D.; Goh, T.K. Fungi on submerged wood from the River Coln, England. Mycol. Res. 1999, 103, 1561–1574. [Google Scholar] [CrossRef]
- Borse, B.; Pawar, N.; Patil, S. Bactrodesmium aquatica sp. nov.(mitosporic fungi) on submerged wood from freshwater, Maharashtra, India. Int. J. Adv. Res. 2019, 7, 1365–1368. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Goh, T.K.; Ho, W.H.; Hyde, K.D.; Whitton, S.R.; Umali, T.E. New records and species of Canalisporium (Hyphomycetes), with a revision of the genus. Can. J. Bot. 1998, 76, 142–152. [Google Scholar] [CrossRef]
- Cai, L.; Kq, Z.; McKenzie, E.; Lumyong, S.; Hyde, K. New species of Canalisporium and Dictyosporium from China and a note on the differences between these genera. Cryptogam. Mycol. 2003, 24, 3–11. [Google Scholar]
- Ferrer, A. New records and a new species of Canalisporium from aquatic habitats in Panama. Mycotaxon 2005, 93, 179–188. [Google Scholar]
- Goh, T.K.; Hyde, K.D. A new species of Canalisporium from Australia. Mycologia 2000, 92, 589–592. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Hyde, K.D.; Mckenzie, E.H.C.; Jones, E.B.G.; Alsadi, A.M.; Lumyong, S. Fuscosporellales, a New Order of Aquatic and Terrestrial Hypocreomycetidae (Sordariomycetes). Cryptogam. Mycologie 2016, 4, 449–475. [Google Scholar] [CrossRef]
- Du, H.Z.; Yang, J.; Liu, N.G.; Cheewangkoon, R.; Liu, J.K. Morpho-Phylogenetic Evidence Reveals New Species of Fuscosporellaceae and Savoryellaceae from Freshwater Habitats in Guizhou Province, China. J. Fungi 2022, 8, 1138. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.K.M.; Hyde, K.D. Biodiversity of fungi on submerged wood in a stream and its estuary in the Tai Ho Bay, Hong Kong. Fungal Divers. 2004, 15, 205–220. [Google Scholar]
- Hernández-Restrepo, M.; Gené, J.; Castañeda-Ruiz, R.F.; Mena-Portales, J.; Crous, P.W.; Guarro, J. Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 2017, 86, 53–97. [Google Scholar] [CrossRef]
- Chang, H. Trichocladium anamorph of Ascotaiwania hsilio and monodictys-like anamorphic states of Ascotaiwania lignicola. Fungal. Sci. 2001, 16, 35–38. [Google Scholar]
- Sivichai, S.; Hywel-Jones, N.; Jones, E. Lignicolous freshwater ascomycota from Thailand: 1. Ascotaiwania sawada and its anamorph state Monotosporella. Mycoscience 1998, 39, 307–311. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).