
Hericium erinaceus in Neurodegenerative Diseases: From Bench to Bedside and Beyond, How Far from the Shoreline?
 ,
, 
 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Hericium erinaceus
3. Untargeted Metabolomics as a New Approach for Hericium erinaceus Metabolite Detection
4. Hericium erinaceus Bioactive Metabolites

5. H. erinaceus In Vitro Preclinical Studies and Molecular Mechanism Underlying the Nootropic Effects
6. H. Erinaceus and Its Potential within the Gut-Microbiome–Brain Axis
| Model/Condition/Disease | Treatment (Doses and Duration) | Increased Bacteria | Decreased Bacteria | Other Effects | Reference |
|---|---|---|---|---|---|
| In vitro (after simulated gastrointestinal digestion and 24-h-fermentation) on fermentation broth of human feces | HEPs 1 (HEP-30, HEP-50, and HEP-70) | SCFA 2-producing bacteria: Lactobacillus, Faecalibacterium, Bifidobacterium, Blautia, Butyricicoccus | Pathogenic bacteria: Klebsiella, Escherichia-Shigella, Enterobacter | - | [110] |
| In vitro (static batch culture 24 h fermentations and fecal inocula from elderly donors) | HEBS 3 and HEOLRP 4 | - | - | Increasing propionate and butyrate levels. | [113] |
| Healthy adults (30.0 ± 4.9 years old) | 1 g of H. erinaceus powder three times a day (7 days of washout) | SCFA-producing bacteria: Eubacterium rectale, Faecalibacterium prausnitzii, Kineothrix alysoides, Gemmiger formicilis, Fusicatenibacter saccharivorans | Pathobionts: Bacteroides caccae, Streptococcus thermophilus, Romboutsia timonensis | Increasing in GM 5 alpha diversity. GM changes related to blood ALP 6, LDL 7, UA 8, and CREA 9. | [112] |
| Aged dogs | H. erinaceus 0.8 g per bw 10 with daily diets for 16 weeks | Bacteroidetes (order Bacteroidales) | Firmicutes: Streptococcus, Tyzzerella, Campylobacteraceae (genus Campylobacter) | - | [114] |
| Middle (8-week-old) and old (6-month-old)-aged mice | HEPs from the H. erinaceus sporophore 0.5 or 1 g/bw/day by gavage for 28 days | Lachnospiraceae, Akkermansiaceae | Rikenellaceae, Bacteroidaceae | Anti-inflammatory activity and immunomodulatory activity. | [115] |
| Acetic-acid-induced UC 11 adult rats | EP-1 12 0.6 and 1.2 g/kg by gavage for 11 days | SCFA producers | - | Antioxidant, anti-inflammatory, and immunoregulatory activities. Restoring a normal GM. | [116] |
| DSS 13-induced UC C57BL/6 adult male mice | HEP10 14 50, 100, or 200 mg/kg by gavage for 7 days | Akkermansia muciniphila. | Proteobacteria | Restoring GM-alpha and -beta diversities. Anti-inflammatory activity in colons. | [117] |
| IBD 15 adult rats and mice | HE/HEP3 100 mg/(kg * day) by gavage for 14 days | Anti-inflammatory bacteria, Corynebacterium, Bacteroides, Enterobacter, Acinetobacter, Desulfovibrio, Lactobacillus | Pro-inflammatory bacteria | Anti-inflammatory activity. Improving the GM composition. | [118,119] |
| CTX 16-immunodeficient adult mice | HEP and FHEP 300 mg/kg/day by gavage for 21 days | Bacteriodetes, Firmicutes | Klebsiella and Shigella | Anti-inflammatory activity. Improving TJ 17 and MUC 18 expression. | [111] |
| CTX-immunodeficient adult mice | HEPs 75, 150, and 300 mg/kg by gavage for 4 weeks | SCFA-producing bacteria, Alistipse, Muribaculaceae, Lachnospiraceae_NK4A136_group, Lachnospiracea, Ruminococcaceae, Ruminococcaceae_UCG-014 | Lactobacillus, Bacteroides, Alloprevotella | Improving the body weight and immune organ index. Increasing OTUs and adjusting GM-alpha and -beta diversities. | [121] |
| Xenograft adult male Balb/C mice, implanting with 100,000 CT-26 wt cancer cells | HEP 100 mg/kg/day per os for 21 days | Probiotic bacteria: Bifidobacterium, Gemellales, Blautia, Sutterella, Anaerostipes, Roseburia, Lachnobacterium, Lactobacillus, and Desulfovibrio | Parabacteroides, Christensenellaceae, Anoxybacillus, Staphylococcus, Aggregatibacter, Comamonadaceae, Desulfovibrionaceae, Sporosarcina, Planococcaceae, Aerococcaceae, Flavobacteriaceae, and Bilophila | Reducing the 5-Fu-induced GM dysbiosis, suppressing tumor growth, and inhibiting inflammatory markers. | [120] |
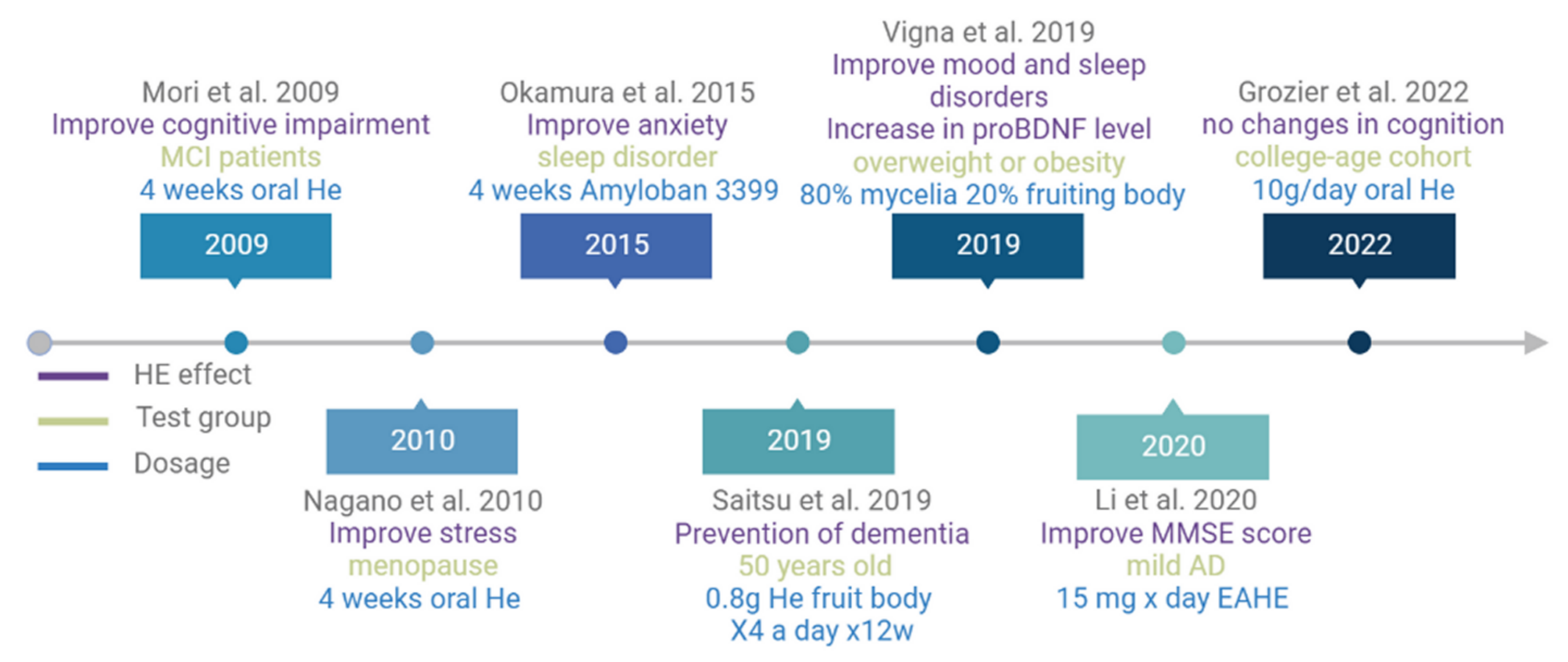
7. H. Erinaceus Pilot Clinical Trials on Antidepressants
8. H. Erinaceus Pilot Clinical Trials on Cognitive Functions
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sindi, S.; Kåreholt, I.; Eskelinen, M.; Hooshmand, B.; Lehtisalo, J.; Soininen, H.; Ngandu, T.; Kivipelto, M. Healthy Dietary Changes in Midlife Are Associated with Reduced Dementia Risk Later in Life. Nutrients 2018, 10, 1649. [Google Scholar] [CrossRef]
- Park, B.-Y.; Lee, M.J.; Kim, M.; Kim, S.-H.; Park, H. Structural and Functional Brain Connectivity Changes between People with Abdominal and Non-abdominal Obesity and Their Association With Behaviors of Eating Disorders. Front. Neurosci. 2018, 12, 741. [Google Scholar] [CrossRef]
- Bolton, J.L.; Bilbo, S.D. Developmental programming of brain and behavior by perinatal diet: Focus on inflammatory mechanisms. Dialogues Clin. Neurosci. 2014, 16, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.J.; Korosi, A.; Layé, S.; Shukitt-Hale, B.; Barrientos, R.M. Food for thought: How nutrition impacts cognition and emotion. NPJ Sci. Food 2017, 1, 7. [Google Scholar] [CrossRef]
- Feng, L.; Cheah, I.K.-M.; Ng, M.M.-X.; Li, J.; Chan, S.M.; Lim, S.L.; Mahendran, R.; Kua, E.-H.; Halliwell, B. The Association between Mushroom Consumption and Mild Cognitive Impairment: A Community-Based Cross-Sectional Study in Singapore. J. Alzheimers Dis. 2019, 68, 197–203. [Google Scholar] [CrossRef]
- Yu, R.; Sun, Y.; Ye, K.X.; Feng, Q.; Lim, S.L.; Mahendran, R.; Cheah, I.K.-M.; Foo, R.S.Y.; Chua, R.Y.; Gwee, X.; et al. Cohort profile: The Diet and Healthy Aging (DaHA) study in Singapore. Aging 2020, 12, 23889–23899. [Google Scholar] [CrossRef] [PubMed]
- Gargano, M.L.; Venturella, G.; Bosco, F.; Fabrini, M.G.; Gao, W.; Pecoraro, L. The Role of Medicinal Mushrooms in Brain Cancer Therapies: Review. Int. J. Med. Mushrooms 2021, 23, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhou, Z.; Zhang, L. An overview of fungal glycan-based therapeutics. Prog. Mol. Biol. Transl. Sci. 2019, 163, 135–163. [Google Scholar] [CrossRef]
- Tsukagoshi, S.; Hashimoto, Y.; Fujii, G.; Kobayashi, H.; Nomoto, K.; Orita, K. Krestin (PSK). Cancer Treat. Rev. 1984, 11, 131–155. [Google Scholar] [CrossRef]
- Tabata, K.; Ito, W.; Kojima, T.; Kawabata, S.; Misaki, A. Ultrasonic degradation of schizophyllan, an antitumor polysaccharide produced by Schizophyllum commune Fries. Carbohydr. Res. 1981, 89, 121–135. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef]
- Blagodatski, A.; Yatsunskaya, M.; Mikhailova, V.; Tiasto, V.; Kagansky, A.; Katanaev, V.L. Medicinal mushrooms as an attractive new source of natural compounds for future cancer therapy. Oncotarget 2018, 9, 29259–29274. [Google Scholar] [CrossRef]
- Hericium erinaceus—An Overview|ScienceDirect Topics. Available online: https://www.sciencedirect.com/topics/pharmacology-toxicology-and-pharmaceutical-science/Hericium-erinaceus (accessed on 6 April 2023).
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal mushroom science: Current perspectives, advances, evidences, and challenges. Biomed. J. 2014, 37, 345–356. [Google Scholar] [CrossRef]
- Li, I.-C.; Lee, L.-Y.; Tzeng, T.-T.; Chen, W.-P.; Chen, Y.-P.; Shiao, Y.-J.; Chen, C.-C. Neurohealth Properties of Hericium erinaceus Mycelia Enriched with Erinacines. Behav. Neurol. 2018, 2018, 5802634. [Google Scholar] [CrossRef]
- Larsson, E.; Larsson, K.-H. Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa. Mycologia 2003, 95, 1037–1065. [Google Scholar] [CrossRef]
- Larsson, K.-H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef]
- Hallenberg, N.; Nilsson, R.H.; Robledo, G. Species complexes in Hericium (Russulales, Agaricomycota) and a new species-Hericium rajchenbergii-from southern South America. Mycol. Prog. 2013, 12, 413–420. [Google Scholar] [CrossRef]
- Das, K.; Stalpers, J.A.; Stielow, J.B. Two new species of hydnoid-fungi from India. IMA Fungus 2013, 4, 359–369. [Google Scholar] [CrossRef]
- Park, H.G.; Kim, S.H.; Ko, H.G.; Park, W.M. Molecular Identification of Asian Isolates of Medicinal Mushroom Hericium erinaceum by Phylogenetic Analysis of Nuclear ITS rDNA. J. Microbiol. Biotechnol. 2004, 14, 816–821. [Google Scholar]
- Cesaroni, V.; Brusoni, M.; Cusaro, C.M.; Girometta, C.; Perini, C.; Picco, A.M.; Rossi, P.; Salerni, E.; Savino, E. Phylogenetic Comparison between Italian and Worldwide Hericium Species (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bishop, K.S.; Tanambell, H.; Buchanan, P.; Quek, S.Y. Assessment of In Vitro Bioactivities of Polysaccharides Isolated from Hericium Novae-Zealandiae. Antioxidants 2019, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Jumbam, B.; Haelewaters, D.; Koch, R.A.; Dentinger, B.T.M.; Henkel, T.W.; Aime, M.C. A new and unusual species of Hericium (Basidiomycota: Russulales, Hericiaceae) from the Dja Biosphere Reserve, Cameroon. Mycol. Prog. 2019, 18, 1253–1262. [Google Scholar] [CrossRef]
- Singh, U.; Das, K. Hericium rajendrae sp. nov. (Hericiaceae, Russulales): An edible mushroom from Indian Himalaya. Nova Hedwig. 2019, 108, 505–515. [Google Scholar] [CrossRef]
- Sugawara, R.; Maekawa, N.; Sotome, K.; Nakagiri, A.; Endo, N. Systematic revision of Hydnum species in Japan. Mycologia 2022, 114, 413–452. [Google Scholar] [CrossRef]
- Ouali, Z.; Sbissi, I.; Boudagga, S.; Rhaiem, A.; Hamdi, C.; Venturella, G.; Saporita, P.; Jaouani, A.; Gargano, M.L. First report of the rare tooth fungus Hericium erinaceus in North African temperate forests. Plant Biosyst. 2020, 154, 24–28. [Google Scholar] [CrossRef]
- Bernicchia, A.; Gorjón, S.P. Corticiaceae s.l.; Candusso, Ed.; Libreria «l’Antro di Ulisse»: Alassio (SV), Italy, 2010. [Google Scholar]
- Kałucka, I.L.; Olariaga Ibarguren, I. IUCN Hericium erinaceus: The IUCN Red List of Threatened Species 2019: E.T70401627A70401637. Available online: https://redlist.info/iucn/species_view/356812/ (accessed on 6 April 2023).
- Stamets, P. Growing Gourmet and Medicinal Mushrooms, 3rd ed.; Ten Speed Press: Berkeley, CA, USA, 2000. [Google Scholar]
- Pelle, J.; Castelli, F.A.; Rudler, M.; Alioua, I.; Colsch, B.; Fenaille, F.; Junot, C.; Thabut, D.; Weiss, N. Metabolomics in the understanding and management of hepatic encephalopathy. Anal. Biochem. 2022, 636, 114477. [Google Scholar] [CrossRef]
- Yang, F.; Wang, H.; Feng, G.; Zhang, S.; Wang, J.; Cui, L. Rapid Identification of Chemical Constituents in Hericium erinaceus Based on LC-MS/MS Metabolomics. J. Food Qual. 2021, 2021, e5560626. [Google Scholar] [CrossRef]
- Commisso, M.; Negri, S.; Bianconi, M.; Gambini, S.; Avesani, S.; Ceoldo, S.; Avesani, L.; Guzzo, F. Untargeted and Targeted Metabolomics and Tryptophan Decarboxylase In Vivo Characterization Provide Novel Insight on the Development of Kiwifruits (Actinidia deliciosa). Int. J. Mol. Sci. 2019, 20, 897. [Google Scholar] [CrossRef]
- Satria, D.; Tamrakar, S.; Suhara, H.; Kaneko, S.; Shimizu, K. Mass Spectrometry-Based Untargeted Metabolomics and α-Glucosidase Inhibitory Activity of Lingzhi (Ganoderma lingzhi) During the Developmental Stages. Molecules 2019, 24, 2044. [Google Scholar] [CrossRef]
- Flores, G.A.; Girometta, C.E.; Cusumano, G.; Angelini, P.; Tirillini, B.; Ianni, F.; Blasi, F.; Cossignani, L.; Pellegrino, R.M.; Emiliani, C.; et al. Untargeted Metabolomics Used to Describe the Chemical Composition, Antioxidant and Antimicrobial Effects of Extracts from Pleurotus spp. Mycelium Grown in Different Culture Media. Antibiotics 2022, 11, 1468. [Google Scholar] [CrossRef]
- Lee, L.-Y.; Chou, W.; Chen, W.-P.; Wang, M.-F.; Chen, Y.-J.; Chen, C.-C.; Tung, K.-C. Erinacine A-Enriched Hericium erinaceus Mycelium Delays Progression of Age-Related Cognitive Decline in Senescence Accelerated Mouse Prone 8 (SAMP8) Mice. Nutrients 2021, 13, 3659. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Zhang, D.-D.; Yin, J.-Y.; Nie, S.-P.; Xie, M.-Y. Recent developments in Hericium erinaceus polysaccharides: Extraction, purification, structural characteristics and biological activities. Crit. Rev. Food Sci. Nutr. 2019, 59, S96–S115. [Google Scholar] [CrossRef]
- Tripodi, F.; Falletta, E.; Leri, M.; Angeloni, C.; Beghelli, D.; Giusti, L.; Milanesi, R.; Sampaio-Marques, B.; Ludovico, P.; Goppa, L.; et al. Anti-Aging and Neuroprotective Properties of Grifola frondosa and Hericium erinaceus Extracts. Nutrients 2022, 14, 4368. [Google Scholar] [CrossRef]
- Roda, E.; Priori, E.C.; Ratto, D.; De Luca, F.; Di Iorio, C.; Angelone, P.; Locatelli, C.A.; Desiderio, A.; Goppa, L.; Savino, E.; et al. Neuroprotective Metabolites of Hericium erinaceus Promote Neuro-Healthy Aging. Int. J. Mol. Sci. 2021, 22, 6379. [Google Scholar] [CrossRef]
- Roda, E.; De Luca, F.; Ratto, D.; Priori, E.C.; Savino, E.; Bottone, M.G.; Rossi, P. Cognitive Healthy Aging in Mice: Boosting Memory by an Ergothioneine-Rich Hericium erinaceus Primordium Extract. Biology 2023, 12, 196. [Google Scholar] [CrossRef]
- Roda, E.; Ratto, D.; De Luca, F.; Desiderio, A.; Ramieri, M.; Goppa, L.; Savino, E.; Bottone, M.G.; Locatelli, C.A.; Rossi, P. Searching for a Longevity Food, We Bump into Hericium erinaceus Primordium Rich in Ergothioneine: The “Longevity Vitamin” Improves Locomotor Performances during Aging. Nutrients 2022, 14, 1177. [Google Scholar] [CrossRef]
- Corana, F.; Cesaroni, V.; Mannucci, B.; Baiguera, R.M.; Picco, A.M.; Savino, E.; Ratto, D.; Perini, C.; Kawagishi, H.; Girometta, C.E.; et al. Array of Metabolites in Italian Hericium erinaceus Mycelium, Primordium, and Sporophore. Molecules 2019, 24, 3511. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef]
- Venturella, G.; Ferraro, V.; Cirlincione, F.; Gargano, M.L. Medicinal Mushrooms: Bioactive Compounds, Use, and Clinical Trials. Int. J. Mol. Sci. 2021, 22, 634. [Google Scholar] [CrossRef]
- Kawagishi, H. Chemical studies on bioactive compounds related to higher fungi. Biosci. Biotechnol. Biochem. 2021, 85, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-F.; Chen, J.-H.; Teng, C.-C.; Shen, C.-H.; Hsieh, M.-C.; Lu, C.-C.; Lee, K.-C.; Lee, L.-Y.; Chen, W.-P.; Chen, C.-C.; et al. Protective effects of Hericium erinaceus mycelium and its isolated erinacine A against ischemia-injury-induced neuronal cell death via the inhibition of iNOS/p38 MAPK and nitrotyrosine. Int. J. Mol. Sci. 2014, 15, 15073–15089. [Google Scholar] [CrossRef] [PubMed]
- Ratto, D.; Corana, F.; Mannucci, B.; Priori, E.C.; Cobelli, F.; Roda, E.; Ferrari, B.; Occhinegro, A.; Di Iorio, C.; De Luca, F.; et al. Hericium erinaceus Improves Recognition Memory and Induces Hippocampal and Cerebellar Neurogenesis in Frail Mice during Aging. Nutrients 2019, 11, 715. [Google Scholar] [CrossRef] [PubMed]
- Krzyczkowski, W.; Malinowska, E.; Herold, F. Erinacine A biosynthesis in submerged cultivation of Hericium erinaceum: Quantification and improved cultivation. Eng. Life Sci. 2010, 10, 446–457. [Google Scholar] [CrossRef]
- Lee, D.G.; Kang, H.-W.; Park, C.-G.; Ahn, Y.-S.; Shin, Y. Isolation and identification of phytochemicals and biological activities of Hericium ernaceus and their contents in Hericium strains using HPLC/UV analysis. J. Ethnopharmacol. 2016, 184, 219–225. [Google Scholar] [CrossRef]
- Beelman, R.B.; Kalaras, M.D.; Phillips, A.T.; Richie, J.P. Is ergothioneine a “longevity vitamin” limited in the American diet? J. Nutr. Sci. 2020, 9, e52. [Google Scholar] [CrossRef]
- Borodina, I.; Kenny, L.C.; McCarthy, C.M.; Paramasivan, K.; Pretorius, E.; Roberts, T.J.; van der Hoek, S.A.; Kell, D.B. The biology of ergothioneine, an antioxidant nutraceutical. Nutr. Res. Rev. 2020, 33, 190–217. [Google Scholar] [CrossRef]
- Cheah, I.K.; Halliwell, B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochim. Biophys. Acta 2012, 1822, 784–793. [Google Scholar] [CrossRef]
- Jang, J.-H.; Aruoma, O.I.; Jen, L.-S.; Chung, H.Y.; Surh, Y.-J. Ergothioneine rescues PC12 cells from beta-amyloid-induced apoptotic death. Free Radic. Biol. Med. 2004, 36, 288–299. [Google Scholar] [CrossRef]
- Salama, S.A.; Abd-Allah, G.M.; Mohamadin, A.M.; Elshafey, M.M.; Gad, H.S. Ergothioneine mitigates cisplatin-evoked nephrotoxicity via targeting Nrf2, NF-κB, and apoptotic signaling and inhibiting γ-glutamyl transpeptidase. Life Sci. 2021, 278, 119572. [Google Scholar] [CrossRef]
- Tang, R.M.Y.; Cheah, I.K.-M.; Yew, T.S.K.; Halliwell, B. Distribution and accumulation of dietary ergothioneine and its metabolites in mouse tissues. Sci. Rep. 2018, 8, 1601. [Google Scholar] [CrossRef]
- Cheah, I.K.; Tang, R.M.Y.; Yew, T.S.Z.; Lim, K.H.C.; Halliwell, B. Administration of Pure Ergothioneine to Healthy Human Subjects: Uptake, Metabolism, and Effects on Biomarkers of Oxidative Damage and Inflammation. Antioxid. Redox Signal. 2017, 26, 193–206. [Google Scholar] [CrossRef]
- Halliwell, B.; Cheah, I.K.; Drum, C.L. Ergothioneine, an adaptive antioxidant for the protection of injured tissues? A hypothesis. Biochem. Biophys. Res. Commun. 2016, 470, 245–250. [Google Scholar] [CrossRef]
- Halliwell, B.; Cheah, I.K.; Tang, R.M.Y. Ergothioneine—A diet-derived antioxidant with therapeutic potential. FEBS Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef]
- Yang, N.-C.; Lin, H.-C.; Wu, J.-H.; Ou, H.-C.; Chai, Y.-C.; Tseng, C.-Y.; Liao, J.-W.; Song, T.-Y. Ergothioneine protects against neuronal injury induced by β-amyloid in mice. Food Chem. Toxicol. 2012, 50, 3902–3911. [Google Scholar] [CrossRef]
- Whitmore, C.A.; Haynes, J.R.; Behof, W.J.; Rosenberg, A.J.; Tantawy, M.N.; Hachey, B.C.; Wadzinski, B.E.; Spiller, B.W.; Peterson, T.E.; Paffenroth, K.C.; et al. Longitudinal Consumption of Ergothioneine Reduces Oxidative Stress and Amyloid Plaques and Restores Glucose Metabolism in the 5XFAD Mouse Model of Alzheimer’s Disease. Pharmaceuticals 2022, 15, 742. [Google Scholar] [CrossRef]
- Song, T.-Y.; Lin, H.-C.; Chen, C.-L.; Wu, J.-H.; Liao, J.-W.; Hu, M.-L. Ergothioneine and melatonin attenuate oxidative stress and protect against learning and memory deficits in C57BL/6J mice treated with D-galactose. Free Radic. Res. 2014, 48, 1049–1060. [Google Scholar] [CrossRef]
- Song, T.-Y.; Chen, C.-L.; Liao, J.-W.; Ou, H.-C.; Tsai, M.-S. Ergothioneine protects against neuronal injury induced by cisplatin both in vitro and in vivo. Food Chem. Toxicol. 2010, 48, 3492–3499. [Google Scholar] [CrossRef]
- Nakamichi, N.; Nakao, S.; Nishiyama, M.; Takeda, Y.; Ishimoto, T.; Masuo, Y.; Matsumoto, S.; Suzuki, M.; Kato, Y. Oral Administration of the Food-Derived Hydrophilic Antioxidant Ergothioneine Enhances Object Recognition Memory in Mice. Curr. Mol. Pharmacol. 2021, 14, 220–233. [Google Scholar] [CrossRef]
- Rossi, P.; Cesaroni, V.; Brandalise, F.; Occhinegro, A.; Ratto, D.; Perrucci, F.; Lanaia, V.; Girometta, C.; Orrù, G.; Savino, E. Dietary Supplementation of Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Agaricomycetes), and Spatial Memory in Wild-Type Mice. Int. J. Med. Mushrooms 2018, 20, 485–494. [Google Scholar] [CrossRef]
- Dubost, N.J.; Beelman, R.B.; Royse, D.J. Influence of selected cultural factors and postharvest storage on ergothioneine content of common button mushroom Agaricus bisporus (J. Lge) Imbach (Agaricomycetes). Int. J. Med. Mushrooms 2007, 9, 163–176. [Google Scholar] [CrossRef]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A rich source of the antioxidants ergothioneine and glutathione. Food Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.R.; Lee, W.Y.; Ka, K.H. Comparative Study on the Sawdust Cultivation and the Antioxidants of Hericium spp. Korean J. Mycol. 2009, 37, 80–85. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Ho, K.-J.; Hsieh, Y.-J.; Wang, L.-T.; Mau, J.-L. Contents of lovastatin, γ-aminobutyric acid and ergothioneine in mushroom fruiting bodies and mycelia. LWT-Food Sci. Technol. 2012, 47, 274–278. [Google Scholar] [CrossRef]
- Tzeng, T.-T.; Chen, C.-C.; Chen, C.-C.; Tsay, H.-J.; Lee, L.-Y.; Chen, W.-P.; Shen, C.-C.; Shiao, Y.-J. The Cyanthin Diterpenoid and Sesterterpene Constituents of Hericium erinaceus Mycelium Ameliorate Alzheimer’s Disease-Related Pathologies in APP/PS1 Transgenic Mice. Int. J. Mol. Sci. 2018, 19, 598. [Google Scholar] [CrossRef]
- Chiu, C.-H.; Chyau, C.-C.; Chen, C.-C.; Lee, L.-Y.; Chen, W.-P.; Liu, J.-L.; Lin, W.-H.; Mong, M.-C. Erinacine A-Enriched Hericium erinaceus Mycelium Produces Antidepressant-Like Effects through Modulating BDNF/PI3K/Akt/GSK-3β Signaling in Mice. Int. J. Mol. Sci. 2018, 19, 341. [Google Scholar] [CrossRef]
- Li, I.-C.; Lee, L.-Y.; Chen, Y.-J.; Chou, M.-Y.; Wang, M.-F.; Chen, W.-P.; Chen, Y.-P.; Chen, C.-C. Erinacine A-enriched Hericium erinaceus mycelia promotes longevity in Drosophila melanogaster and aged mice. PLoS ONE 2019, 14, e0217226. [Google Scholar] [CrossRef]
- Nagano, M.; Shimizu, K.; Kondo, R.; Hayashi, C.; Sato, D.; Kitagawa, K.; Ohnuki, K. Reduction of depression and anxiety by 4 weeks Hericium erinaceus intake. Biomed. Res. Tokyo Jpn. 2010, 31, 231–237. [Google Scholar] [CrossRef]
- Mori, K.; Inatomi, S.; Ouchi, K.; Azumi, Y.; Tuchida, T. Improving effects of the mushroom Yamabushitake (Hericium erinaceus) on mild cognitive impairment: A double-blind placebo-controlled clinical trial. Phytother. Res. 2009, 23, 367–372. [Google Scholar] [CrossRef]
- Saitsu, Y.; Nishide, A.; Kikushima, K.; Shimizu, K.; Ohnuki, K. Improvement of cognitive functions by oral intake of Hericium erinaceus. Biomed. Res. Tokyo Jpn. 2019, 40, 125–131. [Google Scholar] [CrossRef]
- Tsai, P.-C.; Wu, Y.-K.; Hu, J.-H.; Li, I.-C.; Lin, T.-W.; Chen, C.-C.; Kuo, C.-F. Preclinical Bioavailability, Tissue Distribution, and Protein Binding Studies of Erinacine A, a Bioactive Compound from Hericium erinaceus Mycelia Using Validated LC-MS/MS Method. Molecules 2021, 26, 4510. [Google Scholar] [CrossRef]
- Hu, J.-H.; Li, I.-C.; Lin, T.-W.; Chen, W.-P.; Lee, L.-Y.; Chen, C.-C.; Kuo, C.-F. Absolute Bioavailability, Tissue Distribution, and Excretion of Erinacine S in Hericium erinaceus Mycelia. Molecules 2019, 24, 1624. [Google Scholar] [CrossRef]
- Zhang, C.-C.; Cao, C.-Y.; Kubo, M.; Harada, K.; Yan, X.-T.; Fukuyama, Y.; Gao, J.-M. Chemical Constituents from Hericium erinaceus Promote Neuronal Survival and Potentiate Neurite Outgrowth via the TrkA/Erk1/2 Pathway. Int. J. Mol. Sci. 2017, 18, 1659. [Google Scholar] [CrossRef]
- Shimbo, M.; Kawagishi, H.; Yokogoshi, H. Erinacine A increases catecholamine and nerve growth factor content in the central nervous system of rats. Nutr. Res. 2005, 25, 617–623. [Google Scholar] [CrossRef]
- Chang, C.-H.; Chen, Y.; Yew, X.-X.; Chen, H.-X.; Kim, J.-X.; Chang, C.-C.; Peng, C.-C.; Peng, R.Y. Improvement of erinacine A productivity in Hericium erinaceus mycelia and its neuroprotective bioactivity against the glutamate-insulted apoptosis. LWT-Food Sci. Technol. 2016, 65, 1100–1108. [Google Scholar] [CrossRef]
- Üstün, R.; Ayhan, P. Regenerative activity of Hericium erinaceus on axonal injury model using in vitro laser microdissection technique. Neurol. Res. 2019, 41, 265–274. [Google Scholar] [CrossRef]
- Lai, P.-L.; Naidu, M.; Sabaratnam, V.; Wong, K.-H.; David, R.P.; Kuppusamy, U.R.; Abdullah, N.; Malek, S.N.A. Neurotrophic properties of the Lion’s mane medicinal mushroom, Hericium erinaceus (Higher Basidiomycetes) from Malaysia. Int. J. Med. Mushrooms 2013, 15, 539–554. [Google Scholar] [CrossRef]
- Mori, K.; Obara, Y.; Hirota, M.; Azumi, Y.; Kinugasa, S.; Inatomi, S.; Nakahata, N. Nerve growth factor-inducing activity of Hericium erinaceus in 1321N1 human astrocytoma cells. Biol. Pharm. Bull. 2008, 31, 1727–1732. [Google Scholar] [CrossRef]
- Phan, C.-W.; Lee, G.-S.; Hong, S.-L.; Wong, Y.-T.; Brkljača, R.; Urban, S.; Abd Malek, S.N.; Sabaratnam, V. Hericium erinaceus (Bull.: Fr) Pers. cultivated under tropical conditions: Isolation of hericenones and demonstration of NGF-mediated neurite outgrowth in PC12 cells via MEK/ERK and PI3K-Akt signaling pathways. Food Funct. 2014, 5, 3160–3169. [Google Scholar] [CrossRef]
- Zhang, C.-C.; Yin, X.; Cao, C.-Y.; Wei, J.; Zhang, Q.; Gao, J.-M. Chemical constituents from Hericium erinaceus and their ability to stimulate NGF-mediated neurite outgrowth on PC12 cells. Bioorganic Med. Chem. Lett. 2015, 25, 5078–5082. [Google Scholar] [CrossRef]
- Vigna, L.; Morelli, F.; Agnelli, G.M.; Napolitano, F.; Ratto, D.; Occhinegro, A.; Di Iorio, C.; Savino, E.; Girometta, C.; Brandalise, F.; et al. Hericium erinaceus Improves Mood and Sleep Disorders in Patients Affected by Overweight or Obesity: Could Circulating Pro-BDNF and BDNF Be Potential Biomarkers? Evid.-Based Complement. Altern. Med. 2019, 2019, 7861297. [Google Scholar] [CrossRef] [PubMed]
- Grozier, C.D.; Alves, V.A.; Killen, L.G.; Simpson, J.D.; O’Neal, E.K.; Waldman, H.S. Four Weeks of Hericium erinaceus Supplementation Does Not Impact Markers of Metabolic Flexibility or Cognition. Int. J. Exerc. Sci. 2022, 15, 1366–1380. [Google Scholar] [PubMed]
- Tsai-Teng, T.; Chin-Chu, C.; Li-Ya, L.; Wan-Ping, C.; Chung-Kuang, L.; Chien-Chang, S.; Chi-Ying, H.F.; Chien-Chih, C.; Shiao, Y.J. Erinacine A-Enriched Hericium erinaceus Mycelium Ameliorates Alzheimer’s Disease-Related Pathologies in APPswe/PS1dE9 Transgenic Mice. J. Biomed. Sci. 2016, 23, 49. [Google Scholar] [CrossRef] [PubMed]
- Brandalise, F.; Cesaroni, V.; Gregori, A.; Repetti, M.; Romano, C.; Orrù, G.; Botta, L.; Girometta, C.; Guglielminetti, M.L.; Savino, E.; et al. Dietary Supplementation of Hericium erinaceus Increases Mossy Fiber-CA3 Hippocampal Neurotransmission and Recognition Memory in Wild-Type Mice. Evid.-Based Complement. Altern. Med. 2017, 2017, 3864340. [Google Scholar] [CrossRef]
- Ryu, S.; Kim, H.G.; Kim, J.Y.; Kim, S.Y.; Cho, K.-O. Hericium erinaceus Extract Reduces Anxiety and Depressive Behaviors by Promoting Hippocampal Neurogenesis in the Adult Mouse Brain. J. Med. Food 2018, 21, 174–180. [Google Scholar] [CrossRef]
- Bernard, J.A.; Seidler, R.D. Moving forward: Age effects on the cerebellum underlie cognitive and motor declines. Neurosci. Biobehav. Rev. 2014, 42, 193–207. [Google Scholar] [CrossRef]
- Cordaro, M.; Salinaro, A.T.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Cuzzocrea, S.; Di Paola, R.; Fusco, R.; et al. Key Mechanisms and Potential Implications of Hericium erinaceus in NLRP3 Inflammasome Activation by Reactive Oxygen Species during Alzheimer’s Disease. Antioxidants 2021, 10, 1664. [Google Scholar] [CrossRef]
- Mori, K.; Obara, Y.; Moriya, T.; Inatomi, S.; Nakahata, N. Effects of Hericium erinaceus on amyloid β(25-35) peptide-induced learning and memory deficits in mice. Biomed. Res. Tokyo Jpn. 2011, 32, 67–72. [Google Scholar] [CrossRef]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The Neuroprotective Properties of Hericium erinaceus in Glutamate-Damaged Differentiated PC12 Cells and an Alzheimer’s Disease Mouse Model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, Z.; Yang, X.; Qi, L.; Guo, Y.; Tang, X.; Xie, Y.; Chen, D. Improvement of extraction from Hericium erinaceus on the gut-brain axis in AD-like mice. Brain Res. 2022, 1793, 148038. [Google Scholar] [CrossRef]
- Forsythe, P.; Kunze, W.A.; Bienenstock, J. On communication between gut microbes and the brain. Curr. Opin. Gastroenterol. 2012, 28, 557–562. [Google Scholar] [CrossRef]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef]
- Foster, J.A.; McVey Neufeld, K.-A. Gut-brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Leccioli, V.; Oliveri, M.; Romeo, M.; Berretta, M.; Rossi, P. A New Proposal for the Pathogenic Mechanism of Non-Coeliac/Non-Allergic Gluten/Wheat Sensitivity: Piecing Together the Puzzle of Recent Scientific Evidence. Nutrients 2017, 9, 1203. [Google Scholar] [CrossRef]
- Zhu, X.; Li, B.; Lou, P.; Dai, T.; Chen, Y.; Zhuge, A.; Yuan, Y.; Li, L. The Relationship Between the Gut Microbiome and Neurodegenerative Diseases. Neurosci. Bull. 2021, 37, 1510–1522. [Google Scholar] [CrossRef]
- Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.M.; Martínez-Lara, E.; Blanco, S.; Martín, M.J.; Castanys, E.; Buck, R.; et al. Effects of a human milk oligosaccharide, 2′-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J. Nutr. Biochem. 2015, 26, 455–465. [Google Scholar] [CrossRef]
- Allen, A.P.; Hutch, W.; Borre, Y.E.; Kennedy, P.J.; Temko, A.; Boylan, G.; Murphy, E.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Bifidobacterium longum 1714 as a translational psychobiotic: Modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry 2016, 6, e939. [Google Scholar] [CrossRef]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef]
- Yang, X.-D.; Wang, L.-K.; Wu, H.-Y.; Jiao, L. Effects of prebiotic galacto-oligosaccharide on postoperative cognitive dysfunction and neuroinflammation through targeting of the gut-brain axis. BMC Anesthesiol. 2018, 18, 177. [Google Scholar] [CrossRef]
- Gronier, B.; Savignac, H.M.; Di Miceli, M.; Idriss, S.M.; Tzortzis, G.; Anthony, D.; Burnet, P.W.J. Increased cortical neuronal responses to NMDA and improved attentional set-shifting performance in rats following prebiotic (B-GOS®) ingestion. Eur. Neuropsychopharmacol. 2018, 28, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The Gut-Brain Axis and the Microbiome: Mechanisms and Clinical Implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.J.; Rezoagli, E.; Major, I.; Rowan, N.J.; Laffey, J.G. β-Glucan Metabolic and Immunomodulatory Properties and Potential for Clinical Application. J. Fungi 2020, 6, 356. [Google Scholar] [CrossRef] [PubMed]
- Singdevsachan, S.K.; Auroshree, P.; Mishra, J.; Baliyarsingh, B.; Tayung, K.; Thatoi, H. Mushroom polysaccharides as potential prebiotics with their antitumor and immunomodulating properties: A review. Bioact. Carbohydr. Diet. Fibre 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Fernandes, A.; Nair, A.; Kulkarni, N.; Todewale, N.; Jobby, R. Exploring Mushroom Polysaccharides for the Development of Novel Prebiotics: A Review. Int. J. Med. Mushrooms 2023, 25, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Geng, Y.; Xu, T.; Zou, X.; Mao, R.; Pi, X.; Wu, X.; Huang, L.; Yang, K.; Zeng, X.; et al. Digestive Characteristics of Hericium erinaceus Polysaccharides and Their Positive Effects on Fecal Microbiota of Male and Female Volunteers during In Vitro Fermentation. Front. Nutr. 2022, 9, 858585. [Google Scholar] [CrossRef]
- Su, Y.; Cheng, S.; Ding, Y.; Wang, L.; Sun, M.; Man, C.; Zhang, Y.; Jiang, Y. A comparison of study on intestinal barrier protection of polysaccharides from Hericium erinaceus before and after fermentation. Int. J. Biol. Macromol. 2023, 233, 123558. [Google Scholar] [CrossRef]
- Xie, X.Q.; Geng, Y.; Guan, O.; Ren, Y.; Guo, L.; Lv, Q.; Lu, Z.M.; Shi, J.S.; Xu, Z.H. Influence of Short-Term Consumption of Hericium erinaceus on Serum Biochemical Markers and the Changes of the Gut Microbiota: A Pilot Study. Nutrients 2021, 13, 1008. [Google Scholar] [CrossRef]
- Mitsou, E.K.; Saxami, G.; Stamoulou, E.; Kerezoudi, E.; Terzi, E.; Koutrotsios, G.; Bekiaris, G.; Zervakis, G.I.; Mountzouris, K.C.; Pletsa, V.; et al. Effects of Rich in Β-Glucans Edible Mushrooms on Aging Gut Microbiota Characteristics: An In Vitro Study. Molecules 2020, 25, 2806. [Google Scholar] [CrossRef]
- Cho, H.-W.; Choi, S.; Seo, K.; Kim, K.H.; Jeon, J.-H.; Kim, C.H.; Lim, S.; Jeong, S.; Chun, J.L. Gut microbiota profiling in aged dogs after feeding pet food contained Hericium erinaceus. J. Anim. Sci. Technol. 2022, 64, 937–949. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, H.; Zhao, C.; Ren, L.; Wang, C.; Georgiev, M.I.; Xiao, J.; Zhang, T. Value added immunoregulatory polysaccharides of Hericium erinaceus and their effect on the gut microbiota. Carbohydr. Polym. 2021, 262, 117668. [Google Scholar] [CrossRef]
- Shao, S.; Wang, D.; Zheng, W.; Li, X.; Zhang, H.; Zhao, D.; Wang, M. A unique polysaccharide from Hericium erinaceus mycelium ameliorates acetic acid-induced ulcerative colitis rats by modulating the composition of the gut microbiota, short chain fatty acids levels and GPR41/43 respectors. Int. Immunopharmacol. 2019, 71, 411–422. [Google Scholar] [CrossRef]
- Ren, Y.; Sun, Q.; Gao, R.; Sheng, Y.; Guan, T.; Li, W.; Zhou, L.; Liu, C.; Li, H.; Lu, Z.; et al. Low Weight Polysaccharide of Hericium erinaceus Ameliorates Colitis via Inhibiting the NLRP3 Inflammasome Activation in Association with Gut Microbiota Modulation. Nutrients 2023, 15, 739. [Google Scholar] [CrossRef]
- Chen, D.; Zheng, C.; Yang, J.; Li, J.; Su, J.; Xie, Y.; Lai, G. Immunomodulatory Activities of a Fungal Protein Extracted from Hericium erinaceus through Regulating the Gut Microbiota. Front. Immunol. 2017, 8, 666. [Google Scholar] [CrossRef]
- Chen, D.; Yang, X.; Zheng, C.; Yang, J.; Tang, X.; Chen, J.; Shuai, O.; Xie, Y. Extracts from Hericium erinaceus Relieve In-flammatory Bowel Disease by Regulating Immunity and Gut Microbiota. Oncotarget 2017, 8, 85838–85857. [Google Scholar] [CrossRef]
- Wang, D.; Zhu, X.; Tang, X.; Li, H.; Xie, Y.; Chen, D. Auxiliary antitumor effects of fungal proteins from Hericium erinaceus by target on the gut microbiota. J. Food Sci. 2020, 85, 1872–1890. [Google Scholar] [CrossRef]
- Tian, B.; Liu, R.; Xu, T.; Cai, M.; Mao, R.; Huang, L.; Yang, K.; Zeng, X.; Peilong, S. Modulating effects of Hericium erinaceus polysaccharides on the immune response by regulating gut microbiota in cyclophosphamide-treated mice. J. Sci. Food Agric. 2023, 103, 3050–3064. [Google Scholar] [CrossRef]
- Chong, P.S.; Fung, M.-L.; Wong, K.H.; Lim, L.W. Therapeutic Potential of Hericium erinaceus for Depressive Disorder. Int. J. Mol. Sci. 2019, 21, 163. [Google Scholar] [CrossRef]
- Inanaga, K. Marked improvement of neurocognitive impairment after treatment with compounds from Hericium erinaceum: A case study of recurrent depressive disorder. Pers. Med. Universe 2014, 3, 46–48. [Google Scholar] [CrossRef]
- Okamura, H.; Anno, N.; Tsuda, A.; Inokuchi, T.; Uchimura, N.; Inanaga, K. The effects of Hericium erinaceus (Amyloban® 3399) on sleep quality and subjective well-being among female undergraduate students: A pilot study. Pers. Med. Universe 2015, 4, 76–78. [Google Scholar] [CrossRef]
- Yanshree; Yu, W.S.; Fung, M.L.; Lee, C.W.; Lim, L.W.; Wong, K.H. The Monkey Head Mushroom and Memory Enhancement in Alzheimer’s Disease. Cells 2022, 11, 2284. [Google Scholar] [CrossRef]
- Geda, Y.E. Mild Cognitive Impairment in Older Adults. Curr. Psychiatry Rep. 2012, 14, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Li, I.-C.; Chang, H.-H.; Lin, C.-H.; Chen, W.-P.; Lu, T.-H.; Lee, L.-Y.; Chen, Y.-W.; Chen, Y.-P.; Chen, C.-C.; Lin, D.P.-C. Prevention of Early Alzheimer’s Disease by Erinacine A-Enriched Hericium erinaceus Mycelia Pilot Double-Blind Placebo-Controlled Study. Front. Aging Neurosci. 2020, 12, 155. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}

| He1 | He2 | ||||
|---|---|---|---|---|---|
| Sporophore | Mycelium | Sporophore | Mycelium | Primordium | |
| Erinacine A (µg/g) | - | 150 | - | 105 | - |
| Hericenone C (µg/g) | 500 | - | 1560 | - | - |
| Hericenone D (µg/g) | <20 | - | 188 | - | - |
| L-Ergothioneine (µg/g) | 340 | 580 | 2400 (unpublished data) | 940 (unpublished data) | 1300 |
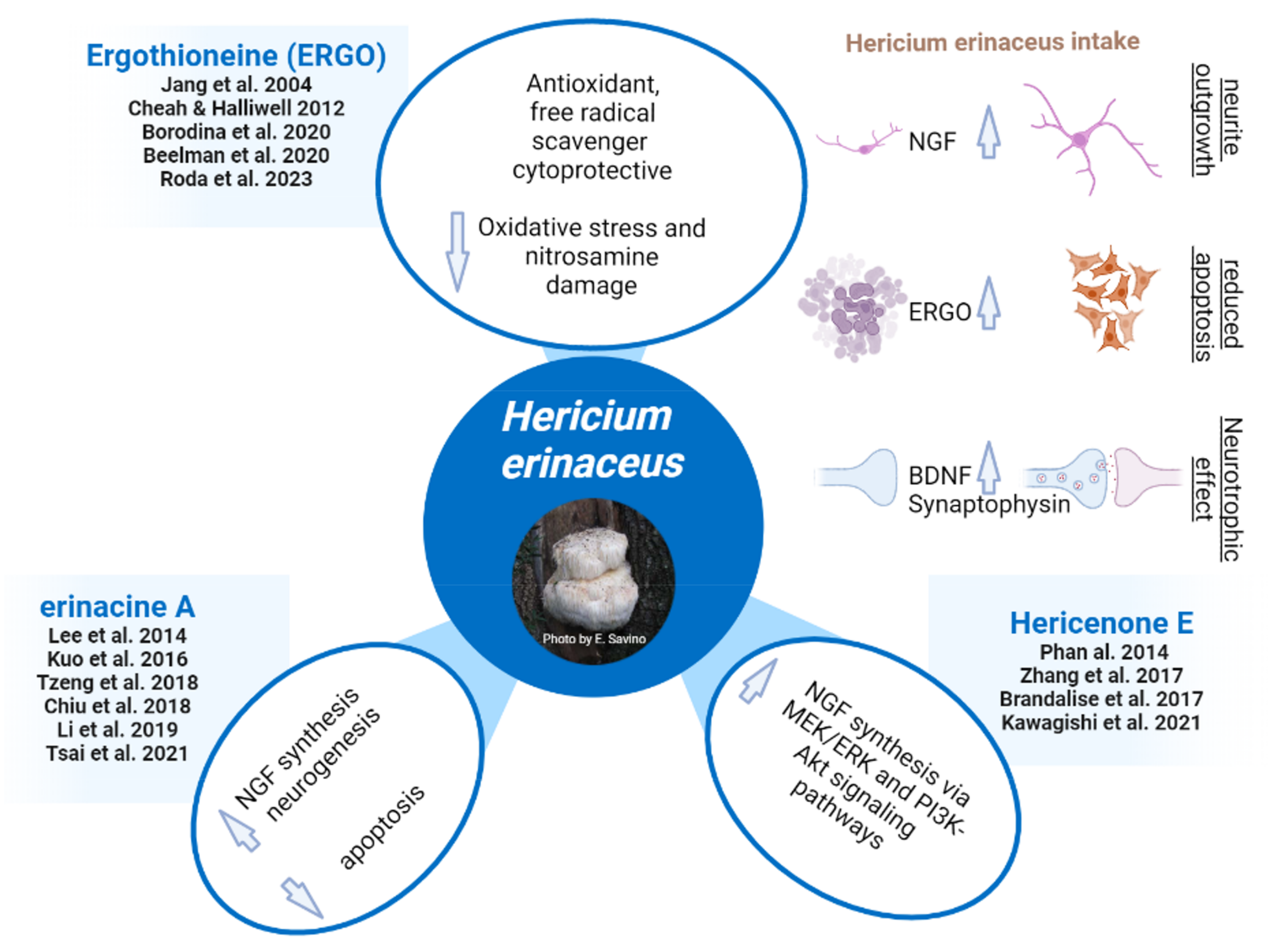
| H. erinaceus Bioactive Compound | Target | Pathway/Intracellular Mechanism | Effect | Reference |
|---|---|---|---|---|
| Erinacine A/ erinacine A-enriched H. erinaceus mycelium (EAHEM) | NGF 1, ↑ NGF/proNGF ratio ↓ iNOS 2 ↑ IDE 5 ↓ 8-OHdG 7, iNOS, and TBARS 8 levels in mice brain | TrkA/Erk1/2 pathway Others? Phosphorylation of p38 MAPK 3 and CHOP 4. ? ? | Neuroprotection and nootropic in vitro. ↑ Neurite outgrowth. ↑ Neuronal proliferation. ↓ Cognitive decline during aging in a SAMP8 mouse model. Protection against stroke in rats. ↓ Inflammation. ↓ Cerebral β-amyloid plaque loading in AD 6 APPswe/PS1dE9 transgenic mice. ↑ Cognitive performances in AD APP/PS1 mice. ↓ Oxidative stress, inflammation, brain Aβ plaque number. | [46,77,78,79,84] [46] [69] [36] |
| Erinacine S | ↑ IDE | ? | ↓ Cerebral β-amyloid plaque loading in AD APPswe/PS1dE9 transgenic mice. | [69] |
| Hericenone E | ↑ NGF | MEK/ERK and PI3K-Akt pathways | ↑ Neurite outgrowth in vitro. | [83] |
| Mix of Erinacine A and Hericenone C and D (sporophore and mycelium H. erinaceus extract) | ↑ Hippocampal and cerebellar PCNA 9 and DCX 10 expression ↓ Cerebellar expression of IL-6 11, iNOS, COX-2 12, and SOD-1 13 | ? ? | ↑ Neurogenesis and cognitive performances in aged frail mice. Inflammation and ↓ Oxidative stress in the cerebellum. ↑ Locomotor functions in aged frail mice. | [47] [39] |
| Ergothioneine-enriched primordium | ↓ Cerebellar expression of iNOS and COX-2 ↓ Hippocampal expression of IL-6, Nrf2 14, SOD-1, COX-2, and iNOS. ↓ Hippocampal NMDAR1 15 and mGlutR2 16 receptors expression | ? TGF-beta1 17 | ↓ Oxidative stress in the cerebellum and hippocampus, and of locomotor aging-related decline in frail mice. ↓ Hippocampal inflammation and oxidative stress. ↑ Glutamatergic neurotransmission. ↓ Cognitive aging-related decline in frail mice. | [41] [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandalise, F.; Roda, E.; Ratto, D.; Goppa, L.; Gargano, M.L.; Cirlincione, F.; Priori, E.C.; Venuti, M.T.; Pastorelli, E.; Savino, E.; et al. Hericium erinaceus in Neurodegenerative Diseases: From Bench to Bedside and Beyond, How Far from the Shoreline? J. Fungi 2023, 9, 551. https://doi.org/10.3390/jof9050551
Brandalise F, Roda E, Ratto D, Goppa L, Gargano ML, Cirlincione F, Priori EC, Venuti MT, Pastorelli E, Savino E, et al. Hericium erinaceus in Neurodegenerative Diseases: From Bench to Bedside and Beyond, How Far from the Shoreline? Journal of Fungi. 2023; 9(5):551. https://doi.org/10.3390/jof9050551
Chicago/Turabian StyleBrandalise, Federico, Elisa Roda, Daniela Ratto, Lorenzo Goppa, Maria Letizia Gargano, Fortunato Cirlincione, Erica Cecilia Priori, Maria Teresa Venuti, Emanuela Pastorelli, Elena Savino, and et al. 2023. "Hericium erinaceus in Neurodegenerative Diseases: From Bench to Bedside and Beyond, How Far from the Shoreline?" Journal of Fungi 9, no. 5: 551. https://doi.org/10.3390/jof9050551
APA StyleBrandalise, F., Roda, E., Ratto, D., Goppa, L., Gargano, M. L., Cirlincione, F., Priori, E. C., Venuti, M. T., Pastorelli, E., Savino, E., & Rossi, P. (2023). Hericium erinaceus in Neurodegenerative Diseases: From Bench to Bedside and Beyond, How Far from the Shoreline? Journal of Fungi, 9(5), 551. https://doi.org/10.3390/jof9050551