
Study on the Structure and Bioactivity of Ganoderma lucidum Polysaccharides under Cassava Stalk Stress

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. The Cultivation of G. lucidum
2.2. Materials and Reagents
2.3. Extraction of GLPs
2.4. Determination of GLPs Content
2.5. Analysis of Monosaccharide Composition of GLPs
2.6. Determination of the Molecular Weight Distribution of GLPs
2.7. Determination of the Antioxidant Activity of GLPs In Vitro
2.8. Determination of G. lucidum Polysaccharides by Near-Infrared Spectroscopy
2.9. Effect of GLPs on the Growth of Lactobacillus rhamnosus LGG
2.10. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Total Sugar and Monosaccharide Content in GLPs
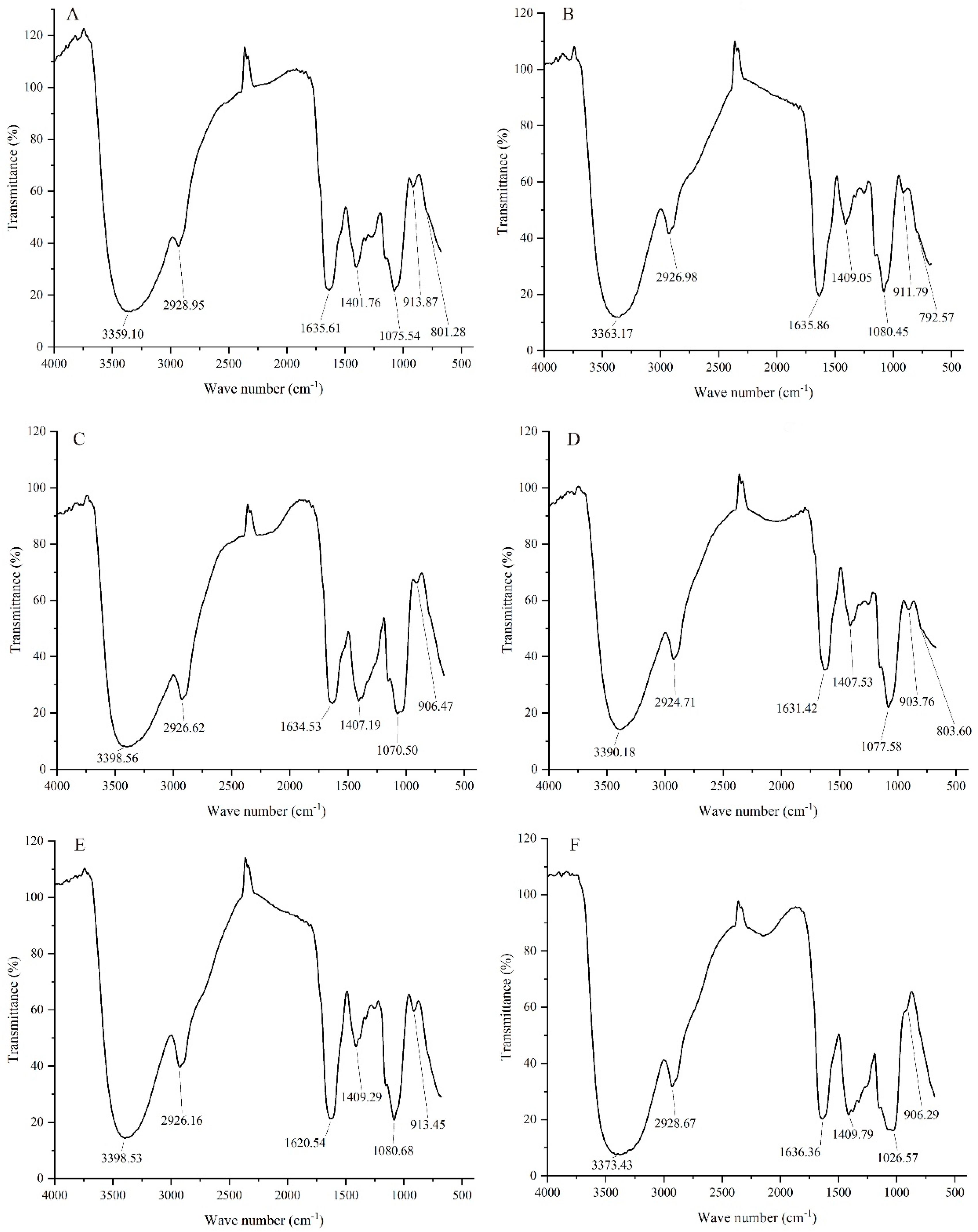
3.2. NIR Analysis of Different GLPs
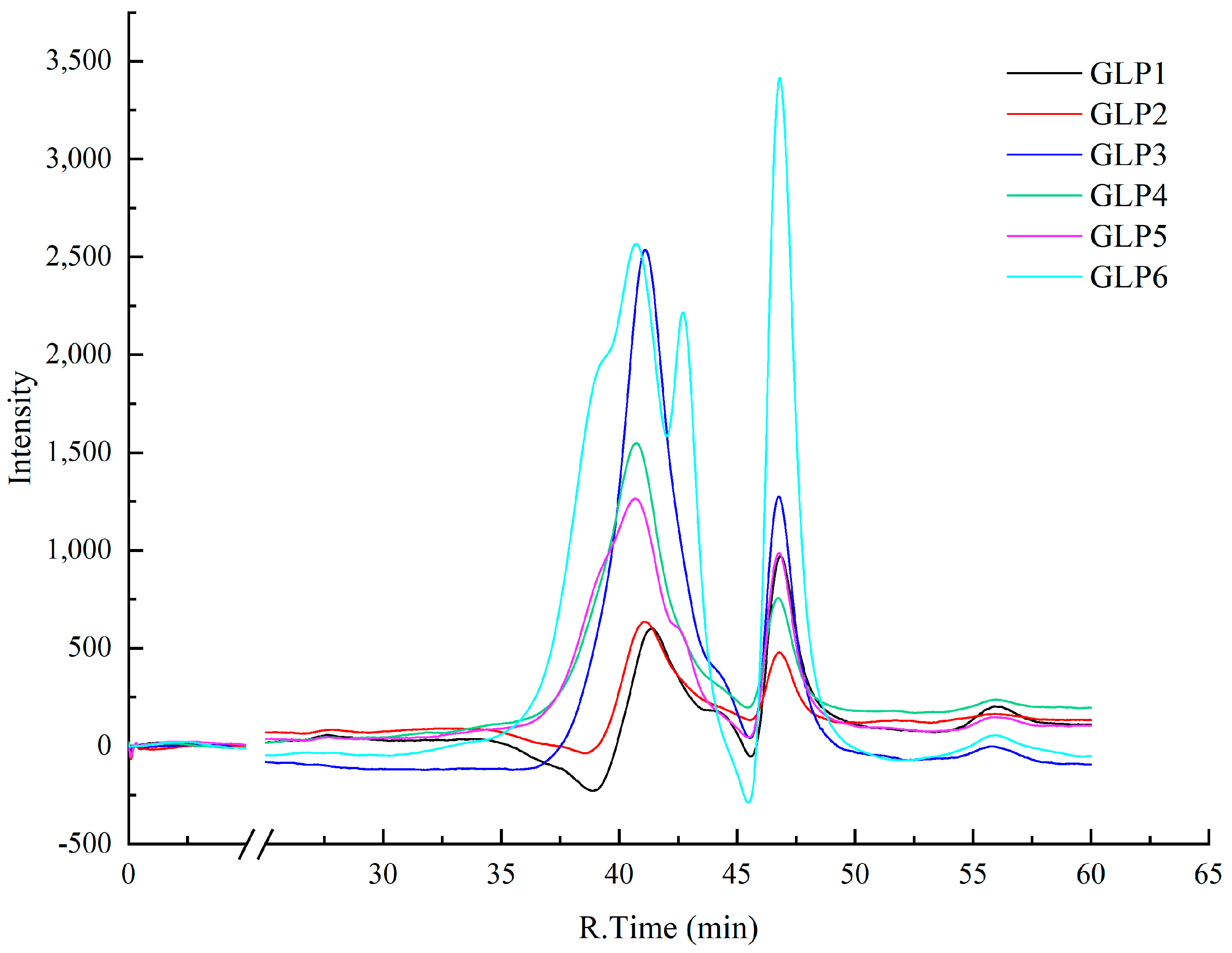
3.3. Analysis of Molecular Weight Distribution of Different GLPs
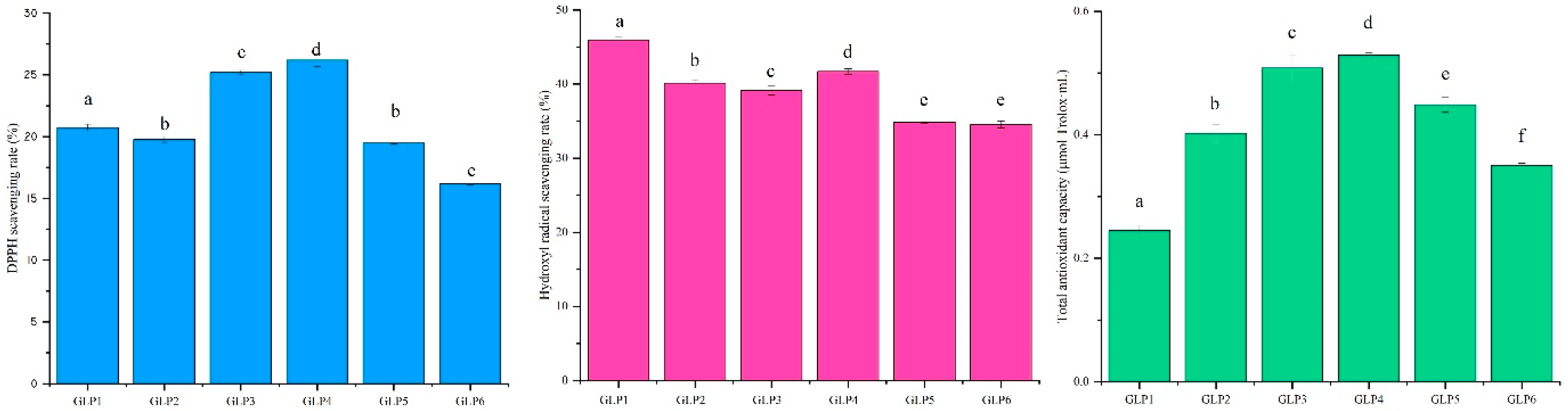
3.4. Antioxidant Activity In Vitro Analysis of Different GLPs
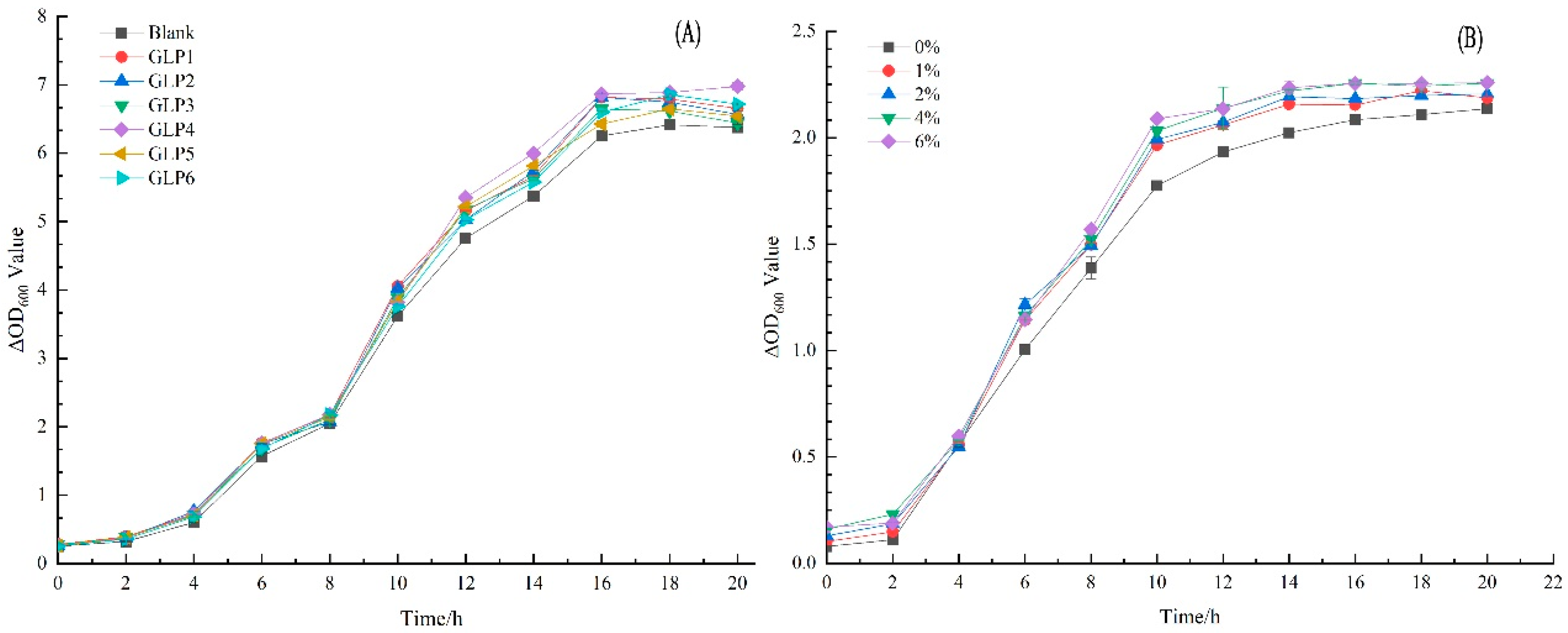
3.5. Effects of GLPs on the Growth of L. rhamnosus LGG
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Molina, J.L.; El-Sharkawy, M.A. Increasing crop productivity in cassava by fertilizing production of planting material. Field Crops Res. 1995, 44, 151–157. [Google Scholar] [CrossRef]
- Jiang, H.C.; Wei, P.L.; Li, N.; Luo, J.J. Fiber morphology and chemical components of cassava stem. Chin. J. Trop. Crops 2015, 36, 1186–1190. [Google Scholar] [CrossRef]
- Huang, X.X.; Li, J.; Li, F.S.; He, D.C. Cultivation techniques of auricularia auricula with cassava stalk. Edible Fungi 2016, 38, 55–56. [Google Scholar] [CrossRef]
- Deng, L.J.; Zou, Z.M.; Chen, Z.S.; Huang, Z.W. Experimental study on Pleurotus geesteranus cultivated by the south advantage resource. Edible Fungi China 2013, 32, 20–22. [Google Scholar] [CrossRef]
- Liu, Y.J.; Bu, M.T.; Liu, Y.Y.; Gong, X.; Tang, Y.D.; Li, J.H. Study on the nutrition quality change in different growth stages of Ganoderma lucidum. J. Yunnan Agric. Univ. 2020, 35, 1061–1066. [Google Scholar] [CrossRef]
- Chen, S.D.; Guan, X.Y.; Yong, T.Q.; Gao, X.; Xiao, C.; Xie, Y.Z.; Chen, D.L.; Hu, H.P.; Wu, Q.P. Structural characterization and hepatoprotective activity of an acidic polysaccharide from Ganoderma lucidum. Food Chem. X 2022, 13, 100204. [Google Scholar] [CrossRef]
- Lin, Z.B. Modern Research on Ganoderma Lucidum, 2nd ed.; Beijing Medical University Press: Beijing, China, 2001. [Google Scholar]
- Peng, L.; Qiao, S.; Xu, Z.; Guan, F.; Ding, Z.Y.; Gu, Z.; Zhang, L.; Shi, G.Y. Effects of culture conditions on monosaccharide composition of Ganoderma lucidum exopolysaccharide and on activities of related enzymes. Carbohydr. Polym. 2015, 133, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qian, Y.; Wang, C.; He, Y.; Zhu, C.; Chen, G.; Lin, L.; Chen, Y. Study of the Metabolite Changes in Ganoderma lucidum under Pineapple Leaf Residue Stress via LC-MS/MS Coupled with a Non-Targeted Metabolomics Approach. Metabolites 2023, 13, 487. [Google Scholar] [CrossRef]
- Lu, J.H.; He, R.J.; Sun, P.L.; Zhang, F.M.; Linhardt, R.J.; Zhang, A.Q. Molecular mechanisms of bioactive polysaccharides from Ganoderma lucidum (Lingzhi), a review. Int. J. Biol. Macromol. 2020, 150, 765–774. [Google Scholar] [CrossRef]
- Nie, S.P.; Zhang, H.; Li, W.J.; Xie, M.Y. Current development of polysaccharides from Ganoderma: Isolation, structure and bioactivities. Bioact. Carbohydr. Diet. Fiber 2013, 1, 10–20. [Google Scholar] [CrossRef]
- Hanyu, X.; Lanyue, L.; Miao, D.; Wentao, F.; Cangran, C.; Hui, S. Effect of Ganoderma applanatum polysaccharides on MAPK/ERK pathway affecting autophagy in breast cancer MCF-7 cells. Int. J. Biol. Macromol. 2020, 146, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nie, S.P.; Ai, L.Z.; Xia, Y.J.; Xiong, Z.Q.; Wang, G.Q. Review on structure and characterization methodology of polysaccharides from Ganoderma. J. Chin. Inst. Food Sci. Technol. 2020, 20, 290–301. [Google Scholar] [CrossRef]
- Michels, M.; Jesus, G.F.A.; Abatti, M.R.; Corneo, E.; Cuker, L.; Medeiros, B.H.; Silva, M.N.; Rocha, L.B.; Dias, R.; Simon, C.S.; et al. Effects of different probiotic strains B. lactis, L. rhamnosus and L. reuteri on brain-intestinal axis immunomodulation in an endotoxin-induced inflammation. Mol. Neurobiol. 2022, 59, 5168–5178. [Google Scholar] [CrossRef]
- Zhao, J.Y.; Li, X.; Xue, B.W.; Wang, J.H.; Li, Z.Z.; Zhang, F.H.; Bai, J. Effect of jujube polysaccharide on growth and metabolism of probiotics. Feed Res. 2022, 45, 73–77. [Google Scholar] [CrossRef]
- Cai, X.L.; He, W.; Yang, G.Q. Research progress on biological activity of Ganoderma lucidum polysaccharide. Edible Fungi 2018, 40, 1–4. [Google Scholar] [CrossRef]
- Song, L.L.; Zhang, Z.Y. The status of biological activity of mushroom polysaccharides. Edible Fungi China 2019, 38, 14–16. [Google Scholar] [CrossRef]
- Han, X.Q.; Yue, G.L.; Yue, R.Q.; Dong, C.X.; Chan, C.L.; Ko, C.H.; Cheung, W.S.; Luo, K.W.; Dai, H.; Wong, C.K.; et al. Structure elucidation and immunomodulatory activity of a beta glucan from the fruiting bodies of Ganoderma sinense. PLoS ONE 2014, 9, e100380. [Google Scholar] [CrossRef]
- Gao, Y.H.; Zhou, S.F.; Jiang, W.Q.; Huang, M.; Dai, X.H. Effects of ganopoly (a Ganoderma lucidum polysaccharide extract) on the immune functions in advanced-stage cancer patients. Immunol. Investig. 2003, 32, 201–215. [Google Scholar] [CrossRef]
- Luo, J.M.; Zhang, C.; Liu, R.; Gao, L.; Ou, S.Y.; Liu, L.; Peng, X.C. Ganoderma lucidum polysaccharide alleviating colorectal cancer by alteration of special gut bacteria and regulation of gene expression of colonic epithelial cells. J. Funct. Foods 2018, 47, 127–135. [Google Scholar] [CrossRef]
- Pan, Y.N.; Yuan, S.L.; Teng, Y.L.; Zhang, Z.; He, Y.M.; Zhang, Y.; Liang, H.H.; Wu, X.; Li, J.Q.; Yang, H.J.; et al. Antioxidation of a proteoglycan from Ganoderma lucidum protects pancreatic β-cells against oxidative stress-induced apoptosis in vitro and in vivo. Int. J. Biol. Macromol. 2022, 200, 470–486. [Google Scholar] [CrossRef]
- Artiningsih, T. Ligninolytic activity of Ganoderma strains on different carbon sources. Biodiversitas 2006, 7, 307–311. [Google Scholar] [CrossRef]
- Liu, L.P.; Feng, J.; Gao, K.; Zhou, S.; Yan, M.Q.; Tang, C.H.; Zhou, J.; Liu, Y.F.; Zhang, J.S. Influence of carbon and nitrogen sources on structural features and immunomodulatory activity of exopolysaccharides from Ganoderma lucidum. Process Biochem. 2022, 119, 96–106. [Google Scholar] [CrossRef]
- Samajpati, N.; Banerjee, S. Studies on the physiology of higher fungi IV effect of different carbon sources on vegetative growth of some species of fomes and Ganoderma. Proc. Indian Natl. Sci. Acad. 2015, 37, 68–73. [Google Scholar]
- Liu, Y.J.; Qian, Y.Y.; Shu, B.; Liu, Y.Y.; Tu, X.H.; Ouyang, H.J.; Li, Y.H.; Tan, G.; Yu, Z.W.; Chen, F.; et al. Effects of four drying methods on Ganoderma lucidum volatile organic compounds analyzed via headspace solid-phase microextraction and comprehensive two-dimensional chromatography-time-of-flight mass spectrometry. Microchem. J. 2021, 166, 106258. [Google Scholar] [CrossRef]
- Liu, J.F.; Feng, M.Y.; You, L.J.; Luo, W.; Zhao, Q.Z.; Zhao, M.M. Structure and antioxidant properties of Ganoderma lucidum polysaccharides extracted by ultrasonic wave and hot water. Sci. Technol. Food Ind. 2013, 34, 88–92. [Google Scholar] [CrossRef]
- He, S.C.; Chen, L.L.; Chen, H.; Yan, J.; Wang, W.J. Effect of different protein removal methods on the structure and antioxidant activity of the cyclocarya paliurus polysaccharides. Sci. Technol. Food Ind. 2023, 157, 604–615. [Google Scholar] [CrossRef]
- You, L.J.; Feng, M.Y.; Liu, J.F.; Zhao, Q.Z.; Luo, W.; Zhao, M.M. Characteristics of Ganoderma Lucidum polysaccharide prepared by different methods. Mod. Food Sci. Technol. 2013, 29, 1207–1212. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, Y.; He, J.; Ouyang, K.H.; Zhao, M.; Cai, L.; Zhao, Z.; Meng, W.Y.; Chen, L.L.; Wang, W.J. Digestive properties and effects of Chimonanthus nitens Oliv polysaccharides on antioxidant effects in vitro and in immunocompromised mice. Int. J. Biol. Macromol. 2021, 185, 306–316. [Google Scholar] [CrossRef]
- Huang, S.Q.; Li, J.W.; Li, Y.Q.; Wang, Z. Purification and structural characterization of a new water-soluble neutral polysaccharide GLP-F1-1 from Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 48, 165–169. [Google Scholar] [CrossRef]
- Liu, X.J.; Yang, L.D.; Li, G.Y.; Jiang, Y.N.; Zhang, G.Y.; Ling, J.Y. A novel promising neuroprotective agent: Ganoderma lucidum polysaccharide. Int. J. Biol. Macromol. 2023, 229, 168–180. [Google Scholar] [CrossRef]
- Liu, Y.F. Structure and Conformation Characterization of Polysaccharide from Ganoderma Lucidum and the Structure-Activity Relationship on Immunoregulation; Jiangnan University: Wuxi, China, 2018; Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CDFDLAST2019&filename=1019019637.nh (accessed on 5 April 2022).
- Ma, Y.H.; He, H.Q.; Wu, J.Z.; Wang, C.Y.; Chao, K.L.; Huang, Q. Assessment of polysaccharides from mycelia of genus Ganoderma by mid-infrared and near-infrared spectroscopy. Sci. Rep. 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H. Structure and Conformational Properties of Polysaccharides from Ganoderma Atrum; Nanchang University: Nanchang, China, 2015; Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CDFDLAST2017&filename=1015629970.nh (accessed on 20 October 2022).
- He, J.Z.; Shao, P.; Meng, X.H.; Sun, P.L. Analysis of structural characteristics of polysaccharide from Ganoderma Lucidum. Chin. J. Anal. Chem. 2010, 38, 372–376. [Google Scholar] [CrossRef]
- Virkki, L.; Johansson, L.; Ylinen, M.; Maunu, S.; Ekholm, P. Structural characterization of water-insoluble nonstarchy polysaccharides of oats and barley. Carbohydr. Polym. 2005, 59, 357–366. [Google Scholar] [CrossRef]
- Sezer, E.; Ayar, A.; Yilmaz, S. Fermentation of dietary fibre-added milk with yoghurt bacteria and L. rhamnosus and use in ice cream production. Fermentation. 2022, 9, 3. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GLP1 | GLP2 | GLP3 | GLP4 | GLP5 | GLP6 | ||
|---|---|---|---|---|---|---|---|
| Total sugar content (%) | 4.07 ± 0.01 | 1.53 ± 0.00 | 1.83 ± 0.00 | 2.23 ± 0.02 | 3.00 ± 0.01 | 3.34 ± 0.01 | |
| D-Fuc | /(mol%) | 0.03 | 0.034 | 0.024 | 0.029 | 0.022 | 0.012 |
| μg/mg | 9.5 | 11.6 | 8.24 | 8.69 | 6.08 | 3.5 | |
| GalN | /(mol%) | 0 | 0 | 0 | 0 | 0 | 0.002 |
| μg/mg | 0 | 0 | 0 | 0 | 0 | 0.73 | |
| L-Rha | /(mol%) | 0 | 0 | 0 | 0 | 0 | 0.011 |
| μg/mg | 0 | 0 | 0 | 0 | 0 | 3.09 | |
| L-Ara | /(mol%) | 0 | 0 | 0 | 0 | 0.029 | 0.052 |
| μg/mg | 0 | 0 | 0 | 0 | 7.5 | 13.5 | |
| GlcN | /(mol%) | 0.011 | 0.01 | 0.008 | 0.008 | 0.01 | 0.013 |
| μg/mg | 4.43 | 4.63 | 3.86 | 3.26 | 3.7 | 4.69 | |
| D-Gal | /(mol%) | 0.36 | 0.322 | 0.254 | 0.26 | 0.205 | 0.11 |
| μg/mg | 125.8 | 120.07 | 97.64 | 85.33 | 63.7 | 34.5 | |
| D-Glc | /(mol%) | 0.528 | 0.552 | 0.66 | 0.642 | 0.643 | 0.707 |
| μg/mg | 184.57 | 205.95 | 253.46 | 211.06 | 199.44 | 221.25 | |
| D-Xyl | /(mol%) | 0.007 | 0.007 | 0.008 | 0.01 | 0.035 | 0.054 |
| μg/mg | 2.1 | 2.04 | 2.46 | 2.84 | 8.95 | 14.01 | |
| D-Man | /(mol%) | 0.065 | 0.074 | 0.046 | 0.05 | 0.056 | 0.04 |
| μg/mg | 22.8 | 27.74 | 17.51 | 16.45 | 17.33 | 12.41 | |
| Sample | RT (min) | lgMp | lgMw | lgMn | Mp (g/mol) | Mw (g/mol) | Mn (g/mol) | Peak Area Ratio (%) |
|---|---|---|---|---|---|---|---|---|
| GLP1 | 41.426 | 4.2 | 4.2 | 4.1 | 15,137 | 17,472 | 12,765 | 84.534 |
| 43.631 | 3.8 | 3.8 | 3.7 | 6172 | 6713 | 5261 | 15.466 | |
| GLP2 | 41.132 | 4.2 | 4.3 | 4.2 | 17,060 | 19,849 | 14,367 | 100 |
| GLP3 | 41.107 | 4.2 | 4.3 | 4.2 | 17,234 | 20,065 | 14,512 | 93.779 |
| 43.729 | 3.8 | 3.8 | 3.7 | 5930 | 6434 | 5057 | 6.221 | |
| GLP4 | 40.728 | 4.3 | 4.4 | 4.2 | 20,108 | 23,651 | 16,900 | 100 |
| GLP5 | 39.796 | 4.5 | 4.5 | 4.4 | 29,380 | 35,435 | 24,582 | 32.965 |
| 40.689 | 4.3 | 4.4 | 4.2 | 20,429 | 24,054 | 17,167 | 50.530 | |
| 42.179 | 4.0 | 4.1 | 4.0 | 11,142 | 12,603 | 9431 | 12.974 | |
| 43.629 | 3.8 | 3.8 | 3.7 | 6177 | 6719 | 5265 | 3.530 | |
| GLP6 | 39.746 | 4.5 | 4.6 | 4.4 | 29,983 | 36,212 | 25,081 | 29.992 |
| 40.708 | 4.3 | 4.4 | 4.2 | 20,272 | 23,857 | 17,037 | 41.719 | |
| 42.701 | 4.0 | 4.0 | 3.9 | 9010 | 10,049 | 7645 | 28.289 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Mai, B.; Li, Z.; Feng, X.; Chen, Y.; Lin, L.; Xia, Q. Study on the Structure and Bioactivity of Ganoderma lucidum Polysaccharides under Cassava Stalk Stress. J. Fungi 2023, 9, 514. https://doi.org/10.3390/jof9050514
Liu Y, Mai B, Li Z, Feng X, Chen Y, Lin L, Xia Q. Study on the Structure and Bioactivity of Ganoderma lucidum Polysaccharides under Cassava Stalk Stress. Journal of Fungi. 2023; 9(5):514. https://doi.org/10.3390/jof9050514
Chicago/Turabian StyleLiu, Yijun, Biyi Mai, Zhiyun Li, Xingqin Feng, Yunlan Chen, Lijing Lin, and Qiuyu Xia. 2023. "Study on the Structure and Bioactivity of Ganoderma lucidum Polysaccharides under Cassava Stalk Stress" Journal of Fungi 9, no. 5: 514. https://doi.org/10.3390/jof9050514
APA StyleLiu, Y., Mai, B., Li, Z., Feng, X., Chen, Y., Lin, L., & Xia, Q. (2023). Study on the Structure and Bioactivity of Ganoderma lucidum Polysaccharides under Cassava Stalk Stress. Journal of Fungi, 9(5), 514. https://doi.org/10.3390/jof9050514