Abstract
Classic paracoccidioidomycosis (PCM) is a potentially deadly neglected tropical systemic mycosis caused by members of the Paracoccidioides brasiliensis complex (P. brasiliensis s. str., P. americana, P. restrepiensis, and P. venezuelensis) and P. lutzii. The laboratorial diagnosis of PCM relies on observing pathognomonic structures such as the “steering wheel” or “Mickey Mouse” shape in the direct mycological examination, fresh biopsied tissue in 10% KOH, histopathological analysis, and/or the isolation of the fungus in culture. However, these procedures are time-consuming and do not allow for the speciation of Paracoccidioides due to overlapping morphologies. Here, we propose a new one-tube multiplex probe-based qPCR assay to detect and recognize agents of the P. brasiliensis complex and P. lutzii. Primers (Paracoco-F and Paracoco-R) and TaqMan probes (PbraCx-Fam, Plu-Ned, and Paracoco-Vic) were developed to target the rDNA (ITS2/28S) in the Paracoccidioides genome. A panel of 77 Paracoccidioides isolates revealed a 100% specificity (AUC = 1.0, 95% CI 0.964–1.000, p < 0.0001) without cross-reacting with other medically relevant fungi or human and murine DNA. The lower limit of detection was 10 fg of gDNA and three copies of the partial rDNA amplicon. Speciation using qPCR was in perfect agreement with AFLP and TUB1-RFLP markers (kappa = 1.0). As a proof of concept, we assessed a panel of 16 formalin-fixed and paraffin-embedded specimens from histopathologically confirmed PCM patients to reveal a significant sensitivity of 81.25% and specificity of 100% (AUC = 0.906 ± 0.05, 95% CI = 0.756–0.979, p < 0.0001, Youden index J = 0.8125). Our assay achieved maximum sensitivity (100%) and specificity (100%) using fresh clinical samples (n = 9) such as sputum, bronchoalveolar lavage, and tissue fragments from PCM patients (AUC = 1.0, 95% CI 0.872–1.000, p < 0.0001, Youden index J = 1.0). Overall, our qPCR assay simplifies the molecular diagnosis of PCM and can be easily implemented in any routine laboratory, decreasing a critical bottleneck for the early treatment of PCM patients across a vast area of the Americas.
1. Introduction
In recent years, a great deal of diversity has been described among Paracoccidioides species, and there is an intense debate about how such diversity translates into taxonomic changes [1,2]. In 2009, the classic Paracoccidioides brasiliensis was split into the P. brasiliensis complex and P. lutzii [3,4]. The P. brasiliensis complex defines a monophyletic clade harboring four cryptic species (P. brasiliensis sensu stricto, P. americana, P. restrepiensis, and P. venezuelensis) with equivalent clinical relevance [5,6]. The recent discovery of P. cetii (a sister species of the P. brasiliensis complex), which is associated with infections in dolphins, and the reclassification of the ancient P. loboi (formerly Lacazia loboi), which causes subcutaneous mycoses in humans, brought to the genus two culture-resistant pathogens, delineating seven species in a genus formerly assumed to be monotypic [1,7].
The interaction of Paracoccidioides species with the warm-blooded vertebrate host leads to the development of paracoccidioidomycosis (PCM), a mycosis ranging from systemic to subcutaneous disease [1]. The classic PCM described in 1908 by Adolpho Lutz remains a life-threatening systemic granulomatous disease acquired by the inhalation of propagules from members of the P. brasiliensis complex and P. lutzii [8]. However, P. loboi (PCM loboi) and P. cetii (PCM ceti) are the culprits of a subcutaneous mycosis caused by implantation, which is characterized by a wide variety of cutaneous lesions, particularly keloidiform nodes [7].
Molecular epidemiology shows that most Paracoccidioides species occur in sympatry with a predominant agent in more than 80% of cases [9]. For example, Paracoccidioides brasiliensis s. str. is the leading agent in Brazil, Argentina, Bolivia, Paraguay, Peru, Uruguay, Venezuela, and the Guadeloupe Islands. To a lesser extent, P. americana is described in Argentina, Brazil, Uruguay, and Venezuela. The two sister species Paracoccidioides restrepiensis and P. venezuelensis are autochthonous to Colombia and Venezuela, respectively, with sporadic cases reported from Argentina, Brazil, Peru, and Uruguay. Paracoccidioides lutzii has an elevated disease burden in the Brazilian Midwest, as well as scattered cases in the Amazon and Peru [4,9,10,11]. Paracoccidioides cetii from cetaceans is detected through the nearshore zones of the Americas, while P. loboi predominates in riverside populations in the Amazon basin and South America [1,7,12].
PCM diagnosis follows clinical suspicion and laboratory analysis. Direct mycological examination of fresh biopsied tissue, sputum, or pus from infected individuals reveals P. brasiliensis complex and P. lutzii multiple budding yeast-like cells producing “steering wheels”, typically pathognomonic in classic PCM. Cultures of clinical specimens showing thermal dimorphism are the reference diagnosis and permit a generic identification [13]. However, a major drawback is that P. brasiliensis complex and P. lutzii have a fastidious nature growing in vitro, taking up to 30 days for colonies to appear [8]. The resistance to culture observed in P. cetii and P. loboi is a symplesiomorphy that demands direct mycological examination and histopathology to detect chains of even-sized yeast-like cells (7–25 μm) in laboratory diagnosis [7].
Conventional methods can be challenging when identifying and distinguishing agents from classic PCM due to overlapping phenotypes [13]. On the other hand, molecular tools can be helpful because they enable differentiation and significantly reduce diagnostics’ turnaround to a few hours. In the current scenario, nested PCR [14], PCR-RFLP (restriction fragment length polymorphism) [15], AFLP fingerprinting [9], multilocus microsatellite typing (MLMT) [16], DNA sequencing followed by phylogenetic analysis (e.g., ITS, ARF, TUB1, and GP43) [17,18], and whole-genome sequencing [19,20] are recommended for recognizing cryptic Paracoccidioides species. However, due to practical and technical challenges, such approaches are not frequently used in clinical laboratories.
Therefore, this study sought to develop a single-tube probe-based quantitative polymerase chain reaction (qPCR) to diagnose classic PCM due to members of the P. brasiliensis complex and P. lutzii. Our strategy uses one to three hydrolysis (TaqMan) probes with fluorescence resonance energy transfer (FRET) technology and a single pair of primers to detect Paracoccidioides DNA originating from isolated cultures, fresh specimens, or formalin-fixed paraffin-embedded tissue samples (FFPE). Moreover, our multiplex qPCR system can potentially boost diagnostic power in Paracoccidioides-affected regions, helping to reduce the time and costs without losing diagnostic sensitivity and specificity.
2. Materials and Methods
2.1. Ethics Approval
Approval was obtained from the ethics committee of the Federal University of São Paulo under protocol numbers 9771060120 and 3147220120.
2.2. Fungal Isolates and Culture Conditions
Seventy-seven Paracoccidioides isolates were obtained from the fungal collection of the Laboratory of Emerging Fungal Pathogens at the Federal University of São Paulo (UNIFESP), São Paulo, Brazil. All isolates were cultivated as slants on a Fava-Netto medium [21,22] at 37 °C in the yeast phase. Paracoccidioides isolates were previously identified at the species level using TUB1-RFLP [15] and AFLP markers [9]. Selected isolates represent various genotypes, origins, and clinical manifestations, and reference strains were used in all experiments (Supplementary Table S1).
2.3. DNA Extraction
Following the manufacturer’s instructions, DNA extraction and purification were performed on 10-day-old yeast colonies employing the Fast DNA kit methodology (MP Biomedicals, Vista, CA, USA) [23]. The NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) was used to determine the concentration and purity of the DNA using the default setting of 1 OD (optical density) = 50 ng/µL double-stranded DNA at wavelengths of 260 and 280 nm. Only DNAs with ODs of 260/280 ratios between 1.8 and 2.0 were kept in this investigation. Before use, DNAs were diluted in nuclease-free water to a concentration of 50 ng/μL and stored at −20 °C. PCRs were conducted as a quality control procedure for Paracoccidioides DNA extraction using the ribosomal DNA operon-targeting universal primers ITS1 and ITS4 [24]. Samples that were successfully amplified were judged to be absent of inhibiting factors.
2.4. Primer and Probe Design and In Silico PCR
The rDNA sequences (18S, ITS1/2 + 5.8S, 28S, IGS) from 31 Paracoccidioides strains were used to develop genus-specific primer pairs and complex-specific probes (Supplementary Table S2) [20]. In addition, DNA sequences previously deposited in GenBank were retrieved to increase variability in the dataset [4,7,17,25,26]. The rDNA sequences were aligned using MAFFT software v. 7 [27] and manually curated in MEGA v7 [28] to prevent mispairing. The development of primers was based on the choice of short regions (18–25 nucleotides) containing parsimonious informative sites that were conserved at genus level and, whenever possible, divergent from other closely related Onygenales. Candidate sequences were evaluated using Primer3 [29] to calculate melting temperatures, %GC contents, dimer sequences, and mismatches. The Mfold software [30] evaluated putative secondary structures in candidate primers that would decrease amplification effectiveness. After all, DNA-based TaqMan probes (20–40 nucleotides) were designed to distinguish P. lutzii (species-specific probe), members of the P. brasiliensis complex (complex-specific probe), or Paracoccidioides species (genus-specific probe). Melting temperatures (Tm), annealing temperature, %GC contents, and mismatches in DNA-based TaqMan probes were calculated using the Primer Express software (Applied Biosystems, Carlsbad, CA, USA), considering MGB or QSY probes.
2.5. In Silico Validation of Primers and Probe Sequences
Primer-BLAST was used to test the specificity of the primers Paracoco-F and Paracoco-R and to conduct additional in silico PCR analysis [31]. To find unintentional, comparable targets, the stringency of primer specificity was tuned to detect sequences with at least 2 total mismatches, and at least 1 mismatch had to occur within the last 5 bps at the 3′ end. Targets with a single primer and one or more mismatches were eliminated. Following this, we mined the Primer-BLAST findings for sequences using the candidate probe sequences.
2.6. qPCR Optimization
The triplex-probe qPCRs were run in a final volume reaction of 20 μL using an Applied Biosystems StepOne Plus real-time PCR system (Applied Biosystems, Carlsbad, CA, USA). Reactions contained 2 μL of gDNA (25–50 ng/μL) as a template, 1.8 µL of sense primer, 0.6 µL antisense primer (10 pmol/μL; Integrated DNA Technologies, San Diego, CA, USA), 0.5 µL of each TaqMan probe (250 nM), 10 μL of TaqMan Genotyping Master Mix (2×) containing the passive internal reference dye ROX (Catalog No. 4371355; Applied Biosystems), and 4.1 μL of nuclease-free water. TaqMan probes (6000 pmol) were purchased from Thermo Fisher Scientific and labeled with FAM (518 nm), VIC (554 nm), or NED (575 nm) at the 5′ ends and with QSY quenchers or the non-fluorescent quencher-minor groove binder (NFQ-MGB) at their 3′-end. Primers and DNA-based FRET probe sequences are listed in Table 1. For the duplex-probe qPCR assay, the P. brasiliensis complex and P. lutzii TaqMan probes were added (0.5 µL each), and the reaction volume was set using nuclease-free water. A unique TaqMan probe (0.5 µL) was added for the singleplex-probe qPCR assay, and the reaction volume was adjusted using nuclease-free water.

Table 1.
Primer and TaqMan probe sequences used in the Paracoccidioides multiplex qPCR assay.
Standard thermal cycling conditions were performed as follows: (1) an initial step of 2 min at 50 °C and 10 min at 95 °C, followed by (2) 40 cycles of 15 s at 95 °C (denature) and 1 min at 60 °C (anneal/extend). The fluorescent signal collection was allowed at the extension step. All qPCR experiments were run with negative controls using nuclease-free water and positive controls (Pb18 and Pb01 gDNA). Thus, positive and negative results were considered valid for a given experiment only after the absence of signal detection for negative controls, along with specific detection for positive controls.
Amplicons generated with primers Paracoco-F and Paracoco-R were purified employing the Wizard SV Gel and PCR Clean-Up System (Promega, Madison, WI, USA). To improve the quality of the sequenced data (Phred ≥ 30), the fungal ITS2 amplicons were control sequenced in two reactions. A BigDye Terminator v3.1 Cycle Sequencing Kit from Applied Biosystems was used for the Sanger sequencing procedures, and a SeqStudio Genetic Analyzer System was used to identify the sequences (Applied Biosystems).
2.7. Determining Assay Specificity In Vitro
The specificity of the TaqMan probes was validated using a triplex-probe qPCR assay along with Paracoccidioides species DNA (n = 77; Supplementary Table S1). Moreover, to investigate our assays’ cross-reactivity with other medically relevant fungi, we used a set of non-target DNAs from agents of superficial, subcutaneous, and systemic mycoses in humans and animals (Supplementary Table S3). Human DNA isolated from A549 cell samples [32] and BALB/c DNA isolated from mouse lungs [33] were used to investigate if our assay cross-reacts with host DNA. A multiplex PCR-based quality-control assay using primers targeting nonoverlapping sites in the GAPDH gene (chr12) of the human genome was used to check the quality of our A549 cell DNA extraction [34], and samples that returned 100, 200, 300, and 400 bp amplicons were deemed to be free of PCR inhibitors [33]. Murine DNA quality was evaluated by conventional PCR using the primers targeting the β-actin gene in the BALB/c genome, as described by Pahl et al. [35], and samples with positive amplification were regarded to be free of PCR inhibitors.
2.8. Determining Assay Sensitivity
A 10-fold serial dilution regression analysis was used to ascertain the detection limits, beginning with 100 ng/µL and finishing with 0.01 fg/µL of Paracoccidioides gDNA. Likewise, we performed a 10-fold serial dilution regression analysis of ITS2 amplicons (amplified using primers Paracoco-F and Paracoco-R), beginning with 3 × 106 copies per tube and finishing with 3 copies per tube. As previously described, each probe’s limit of detection (LoD) was observed under singleplex-, duplex-, and triplex-probe settings [36]. All standard curves were performed in triplicate.
2.9. Target Template Competition
Serial 10-fold dilutions were tested to establish the performance of our triplex-probe qPCR assay to detect a mixed infection of any member of the P. brasiliensis complex and P. lutzii, as previously described by Della Terra et al. [36]. In the first experiment, increasing concentrations of P. brasiliensis complex (Pb18) and P. lutzii (Pb01) were included in the qPCR assays. In a second experiment, to evaluate the template competition, the highest tested concentration of the P. brasiliensis gDNA (i.e., 100 ng) was combined with the lowest tested amount of P. lutzii gDNA (i.e., 0.01 fg) and added to a triplex-probe qPCR assay. The permutations were carried out until the maximum amount of the opposing species’ DNA (i.e., 100 ng of P. lutzii DNA) was combined with the first species’ lowest tested DNA amount (i.e., 0.01 fg of P. brasiliensis DNA). All reactions were run in the presence of 50 ng of human DNA as a spike. In addition, two negative controls using nuclease-free water spiked with or without A549 DNA were run for each qPCR assay.
2.10. Detection of Paracoccidioides DNA from Formalin-Fixed, Paraffin-Embedded (FFPE) Tissue and Other Biological Samples
For the clinical validation assay, sixteen formalin-fixed, paraffin-embedded (FFPE) tissue samples from the tibia (n = 2), duodenum (n = 1), lungs (n = 2), lymph node (n = 5), oral mucosa (n = 2), kidney (n = 1), and skin (n = 3) were obtained from 12 Brazilian patients (14–66 years old; Supplementary Table S4). Histopathology confirmed paracoccidioidomycosis in each sample. Eighteen FFPE tissue samples from patients with other diseases (non-PCM) were included as negative controls: human sporotrichosis, n = 1; human leishmaniasis, n = 10; tuberculosis, n = 6; and histoplasmosis, n = 1. DNA was extracted from three 5 µm thick sections of each FFPE using the QIAamp DNA FFPE Tissue Kit (Qiagen, Hilden, Germany), observing the guidelines provided by the manufacturer [37,38]. All FFPE samples were taken in strict accordance with legal requirements and best-practice standards [39,40,41]. The FFPE isolated DNA was resuspended in 70 µL of elution buffer [38,42], and DNA quality was evaluated by running a quadruplex assay targeting the GAPDH gene (chr12) [34]. Those FFPE DNAs that produced at least three fragments (e.g., 100, 200, and 300 bp) were considered non-refractory to further qPCR analyses. For the triplex-probe qPCR assay, we added 5 µL of FFPE DNA. The standard qPCR thermal cycling conditions were used.
In addition, nine fresh clinical samples (sputum = 1; fresh tissue biopsy = 3; bronchoalveolar lavage = 4; cerebrospinal fluid = 1) were taken from nine patients (30–73 years old) in Brazil (Supplementary Table S4). During the direct mycological examination, PCM was proved in all nine patients by laboratory demonstration of pathognomonic “steering wheel” budding yeast cells. DNA extraction and purification were performed using the Fast DNA kit procedure and the CLS-TS component (MP Biomedicals, Vista, CA, USA), following the manufacturer’s directions [43]. DNA concentration and purity were assessed as described above for FFPE samples. A volume of 5 µL of clinical samples’ DNA was investigated using the triplex-probe qPCR assay.
2.11. Detection of Paracoccidioides DNA from Spiked Soil Samples
Attempts were made to extract DNA from artificially contaminated soil (spiked) to establish the assay’s feasibility for sensitive and specific detection of Paracoccidioides DNA. An aliquot of 50–100 mg soil samples was used for DNA extraction employing the Fast DNA kit procedure and the CLS-VF/PPS component (MP Biomedicals, Vista, CA, USA), following the manufacturer’s directions [43]. Subsequently, we used 2 μL of the soil DNA (100 ng/μL) along with a 10-fold serial dilution of the Paracoccidioides DNA (spiked reactions), beginning with 100 ng/μL and finishing with 0.01 fg/μL. Non-spiked reactions were included as negative controls [33]. The triplex-probe qPCR assay was run as previously specified using the standard cycling conditions.
2.12. Statistical Analysis
Performance indicators such as sensitivity, specificity, positive predictive value (PPV), and negative predictive value (NPV) were calculated for the triplex-probe qPCR assay. To evaluate the usefulness of various tests for diagnosing PCM, we employed the receiver operating characteristic (ROC) curves and the area under the curves (AUC) to assess the diagnostic accuracy of our assay. In addition, we determined the kappa statistic and its 95% confidence interval (CI) to establish the degree of concordance between the results from our triplex-probe qPCR assay and those from the conventional duplex PCR previously reported by our group [33]. The kappa rates were interpreted as follows: values between 0.00–0.20, poor agreement; 0.21–0.40, fair agreement; 0.41–0.60, moderate agreement; 0.61–0.80, good agreement; and 0.81–1.00, perfect agreement [44]. All analyses were carried out using the statistical program MedCalc (v.20.114), and if the p-values were 0.05 or lower, the results were deemed as significant.
3. Results
3.1. Primers and Probes Design
The full dataset comprised 4073 bp long rDNA sequences aligned with MAFFT software and partially covered the 18S, ITS1/2 + 5.8S, and 28S, including 4026 invariable characters (98.84%), 39 variable parsimony-informative sites (0.95%), and 6 singletons. Primers Paracoco-F and Paracoco-R targeted the conserved region (invariable characters) of ITS2 and 28S, respectively. In silico PCR using the primers Paracoco-F and Paracoco-R generated a 143 bp fragment for members of the P. brasiliensis complex and a 137 bp amplicon for P. lutzii. DNA-based FRET probes lying in the ITS2 were designed to deliver a generic identification (Paracoco-Vic probe) or to recognize members of the P. brasiliensis complex (PbraCx-Fam probe) and P. lutzii (Plu-Ned probe) in a single-round reaction (Figure 1; Table 1). To validate our results, we performed an in silico analysis with the primer BLAST software, and no target templates were found in the nucleotide collection (nt) database for species different from Paracoccidioides (Supplementary Table S5). Moreover, our analyses revealed that probe-binding regions were polymorphic among other closely related Onygenales, such as P. cetii and P. loboi, preventing unspecific recognition (Supplementary Figure S1). Therefore, the assay accuracy relies on using the TaqMan probes.
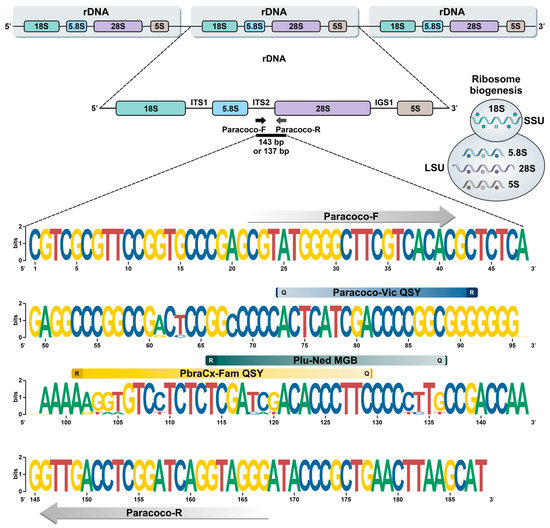
Figure 1.
Graphic representation using WebLogo of polymorphism patterns within the alignment of 31 target sequences of members of the P. brasiliensis complex and P. lutzii (see Supplementary Table S2). Paracoco-F and Paracoco-R primers recognize a highly conserved region for Paracoccidioides species with no mismatches. Parsimony-informative sites in the ITS2 region uncovered candidate areas for DNA-based FRET probes design, i.e., PbraCx-Fam, Plu-Ned, and Paracoco-Vic. R: reporter; Q: quencher.
3.2. qPCR Standardization
Our initial attempts to standardize the triplex-probe qPCR assay using a 3 × 3 matrix (100 nM, 300 nM, and 900 nM) indicated that the forward primer at a final concentration of 900 nM, reverse primer at a final concentration of 300 nM, and probes at a final concentration of 250 nM are the ideal combination for P. brasiliensis (Cq = 20.25 ± 0.09) and P. lutzii DNAs (Cq = 18.28 ± 0.71) as templates.
3.3. qPCR Efficiency
The 10-fold serial dilution regression analysis under a singleplex-, duplex-, and triplex-probe condition with their respective Paracoccidioides species is shown in Figure 2. For the gDNA, it was observed that the amplification covers all dilutions of the qPCR efficiency at good Cq values for the P. brasiliensis probe (singleplex-probe Cq = 15.96–36.70; duplex-probe Cq = 15.28–36.99; triplex-probe Cq = 17.37–36.97), P. lutzii probe (singleplex-probe Cq = 15.62–36.56; duplex-probe Cq = 17.66–37.01; triplex-probe Cq = 16.92–36.42), and Paracoccidioides species probe (P. brasiliensis DNA Cq = 17.33–36.95 and P. lutzii DNA Cq = 16.86–36.31).
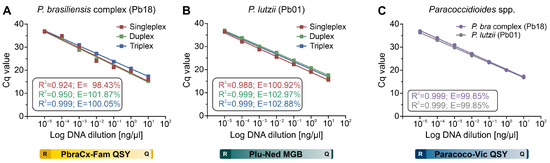
Figure 2.
Standard curves were constructed for (A) P. brasiliensis probe (Pb18 DNA, PbraCx-Fam), (B) P. lutzii probe (Pb01 DNA, Plu-Ned), and (C) Paracoccidioides species probe (Pb18 or Pb01 DNAs, Paracoco-Vic) under a singleplex-probe (red line), a duplex-probe (green line), or a triplex-probe qPCR assay (blue line), showing comparable efficiencies and that the qPCR settings are multiplex-compatible. The results are typical of two separate experiments. R: reporter; Q: quencher.
All standard curves revealed a strong linear correlation (R2 = 0.92–0.99) for all tested probes under any plex-probe condition. Our qPCR assay showed excellent efficiency and reproducibility among replicates in the P. brasiliensis probe (singleplex-probe E = 98.43%; duplex-probe E = 101.87%; triplex-probe E = 100.05%), P. lutzii probe (singleplex-probe E = 100.92%; duplex-probe E = 102.97%; triplex-probe E = 102.88%), and Paracoccidioides species probe (P. brasiliensis DNA E = 99.85% and P. lutzii DNA E = 99.85%). As a result, qPCR efficiency analysis revealed that the primer-probe combinations were consistent with formats ranging from single- to multiplexing.
3.4. Analytical Sensitivity (LoD)
By testing amplification using 10-fold serial gDNA dilutions, we could estimate our assay’s analytical sensitivity, and regardless of the test set, the gDNA detection limit for all targets was 10 fg (singleplex, Figure 3A; duplex, Figure 3B; or triplex, Figure 3C,D). To perform qPCR analysis with greater accuracy, we assessed the analytical sensitivity using the ITS2 amplicon template generated with primers Paracoco-F and Paracoco-R, and under any plex-probe scenarios, three copies of each target were the lowest concentration found (Figure 3E–H).
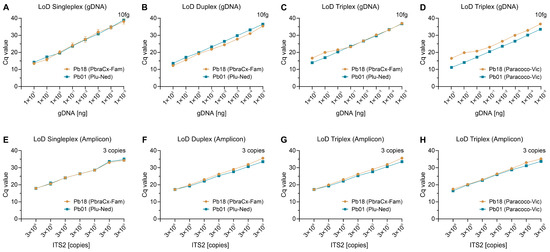
Figure 3.
Assessment of analytical sensitivity of a singleplex- (A,E), duplex- (B,F), and triplex-probe qPCR assay (C,D,G,H). (A–D) For P. brasiliensis (Pb18) and P. lutzii (Pb01), the lower detection limit of gDNA was 10 fg, independent of the format. (E,F) For all species, the lower limit of detection of the ITS2 amplicon (amplified with primers Paracoco-F and Paracoco-R) was three copies. For each format, the detected probe appears in parentheses in the legend. The results are typical of two separate experiments.
3.5. Analytical Specificity
A panel of 77 Paracoccidioides DNA samples (Supplementary Table S1) was used to assess assay specificity in this study, and amplification plots are depicted in Figure 4A (PbraCx-Fam), Figure 4B (Paracoco-Vic), Figure 4D (Plu-Ned), and Figure 4E (Paracoco-Vic). The qPCR resulted in specific amplification products from all members of the P. brasiliensis complex (n = 56; PbraCx-Fam Cq = 10.15–30.97; PbraCx-Fam Median Cq = 15.13; Paracoco-Vic Cq = 9.26–30.97; Paracoco-Vic Median Cq = 16.02; Figure 4C), and P. lutzii (n = 21; Cq = 11.75–18.28; Median Cq = 14.25; Paracoco-Vic Cq = 11.53–19.59; Paracoco-Vic Median Cq = 14.95; Figure 4F). Non-specific amplification products were absent. Our triplex-probe qPCR assay was in full conformity (100%) with the identifications achieved with molecular TUB1-RFLP [15] and AFLP markers [9] (kappa = 1.0; 95% CI = 1.0–1.0; perfect agreement) performed previously at our laboratory, reinforcing the value of differentiating the critical agents of classic PCM. Thus, no cross-reactivity was observed.
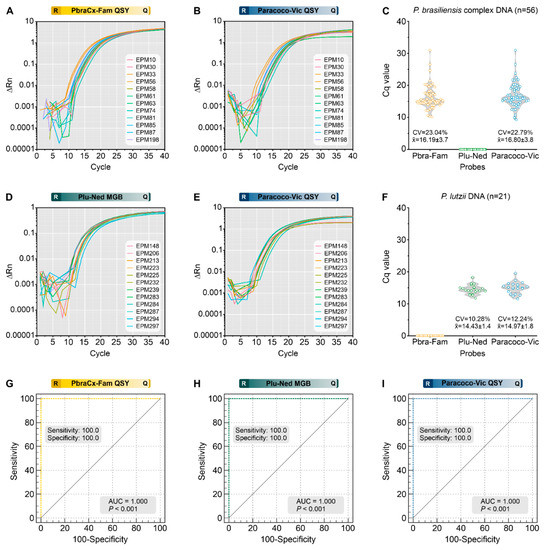
Figure 4.
Assessment of analytical specificity of a triplex-probe qPCR assay for Paracoccidioides species. The representative amplification plot for PbraCx-Fam probe (A) or Paracoco-Vic probe (B) and the DNA of members of the P. brasiliensis complex; (C) Scatter plot of Cq values (y-axis) from a group of 56 P. brasiliensis complex DNA, including P. brasiliensis s. str. (n = 42), P. americana (n = 6), P. restrepiensis (n = 5), and P. venezuelensis (n = 3). The representative amplification plot for Plu-Ned probe (D) or Paracoco-Vic probe (E) and the DNA of P. lutzii; (F) Scatter plot of Cq values (y-axis) from a group of 21 P. lutzii DNA. (G–I) ROC analyses were calculated for 56 isolates of P. brasiliensis complex, 21 isolates of P. lutzii, and 28 isolates of other pathogenic fungi (Supplementary Table S3), revealing the utility of our qPCR test (AUC = 1.000, 95% CI = 0.964–1.000, p < 0.0001) in identifying Paracoccidioides species. (G) ROC analysis for the PbraCx-Fam probe reached 100% specificity and sensitivity as only 56 isolates of P. brasiliensis complex were positive. (H) ROC analysis for the Plu-Ned probe revealed 100% specificity and sensitivity as only 21 isolates of P. lutzii were positive. (I) ROC analysis for the Paracoco-Vic probe showed 100% specificity and sensitivity as 77 Paracoccidioides spp. isolates were positive. CV: coefficient of variation. The results are typical of two separate experiments. R: reporter; Q: quencher.
Additionally, we used DNAs from 25 medically relevant fungi other than Paracoccidioides to confirm the specificity of our assay, and no measurable fluorescent signal was observed. Moreover, the ability of our triplex-probe assay to reveal Paracoccidioides in the presence of host DNA was further supported by the absence of cross-reactivity with human (A549 cell line) or murine DNAs (BALB/c). This finding corroborates the high specificity projected by the BLASTn-search assessments. Finally, AUC investigation provided a clear indication of the assay’s usefulness in discriminating between members of the P. brasiliensis complex and P. lutzii (Figure 4G,H) or by achieving genus-level identification (AUC = 1.000, 95% CI = 0.964–1.000, p < 0.0001) (Figure 4I).
3.6. Target Template Competition
In a hypothetical setting where we had two different targets (for example, P. brasiliensis and P. lutzii) in the same qPCR with varying quantities of gDNA, we assessed the viability of our assay to recognize both targets. In all simulations evaluated, we observed signals for a single Paracoccidioides species along with the signal for the genus probe. We detected both species in those reactions with up to a 10-fold difference in DNA concentration (for example, 10−2 ng of P. brasiliensis DNA and 10−1 ng of P. lutzii DNA) (Figure 5A). On the other hand, equimolar concentrations of the different targets used in a 10-fold serial dilution did not affect the templates’ capacity to be detected by our triplex-probe assay (Figure 5B). Therefore, this suggests that we could detect PCM co-infections, but a higher concentration of one of the targets under a triplex-probe setting will favor its detection over the less abundant target.
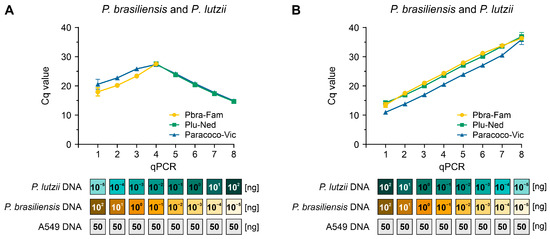
Figure 5.
Assessment of target/template competition. (A) Using a triplex-probe qPCR assay, we combined the gDNA of P. brasiliensis (Pb18) and P. lutzii (Pb01) in a manner that the highest DNA concentration for the first species in the curve was mixed with the lowest concentration in the opposite species’ curve. (B) P. brasiliensis (Pb18) and P. lutzii (Pb01) gDNA were added to the triplex-probe qPCR assay in equimolar amounts. All qPCRs were run in the presence of 50 ng human DNA. The results are typical of two separate experiments.
3.7. Detection of Paracoccidioides DNA from Clinical Samples
FFPE tissue biopsies from patients with confirmed PCM infection (n = 21) and patients with other disorders (n = 18) were used to measure our assay’s ability to detect Paracoccidioides DNA (Supplementary Figure S2). Of the 39 FFPE samples included in the study, 34 tested positive for at least three bands using the GAPDH control, and five samples from the PCM group were removed due to the low quality of FFPE DNA (i.e., no amplification of GAPDH control; Supplementary Figure S2). From the 16 GAPDH-positive samples of the PCM group, 12 specimens were positive for the P. brasiliensis complex probe (Cq = 5.05–24.96; Median Cq = 10.96) and a single specimen for the P. lutzii probe (Plu-Ned Cq = 5.28). All thirteen samples were positive for the Paracoccidioides probe (Cq = 5.30–26.17; Median Cq = 10.25, Figure 6A). Three samples remained negative under the triplex-probe assay after two rounds of qPCR. For the non-PCM group, the 18 samples tested negative for the 3 probes evaluated (Supplementary Figure S2). Sensitivity (81.25%), specificity (100%), the area under the ROC curve (0.906 ± 0.0504; p < 0.0001), Youden index (0.8125), and negative (NPV = 85.67%) and positive predictive values (PPV = 100%) of qPCR were calculated using the results obtained with 16 FFPE samples of PCM patients and 18 FFPE samples of other diseases (95% CI = 68.319–94.313) (Figure 6D).
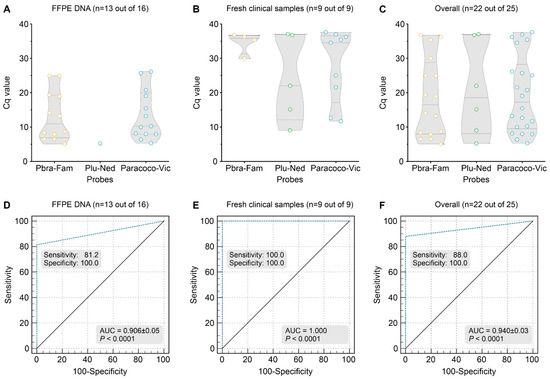
Figure 6.
Summary of FFPE tissues (n = 16) or miscellaneous fresh clinical samples (n = 9), including sputum, pus, tissue biopsy, bronchoalveolar lavage, and cerebrospinal fluid, employed in our triplex-probe qPCR. (A) Scatter plot of Cq values (y-axis) from a group of 16 DNAs revealed that 13 specimens were positive for the Paracoco-Vic probe (81.2%; Mean Cq = 10.25), with 12 P. brasiliensis complex (PbraCx-Fam) and a single P. lutzii detected (Plu-Ned probe). (B) Scatter plot of Cq values (y-axis) from a group of nine DNAs revealed that nine specimens were positive for the Paracoco-Vic probe (100%; Mean Cq = 27.98), with four P. brasiliensis complex (PbraCx-Fam) and five P. lutzii detected (Plu-Ned probe). (C) Scatter plot of Cq values (y-axis) from overall samples (FFPE and fresh clinical samples). (D) ROC analysis for the triplex-probe qPCR reached 100% specificity, and 81.2% sensitivity as 13 FFPE samples were positive and only 3 samples were false-negative (AUC = 0.906 ± 0.05; p < 0.0001, 95% CI = 68.319–94.313, Youden index J = 0.8125, NPV = 85.67%, PPV = 100%). The 18 non-PCM samples remained negative. (E) ROC analysis for the triplex-probe qPCR reached 100% specificity and sensitivity as 9 fresh clinical samples were positive, and the 18 non-PCM samples remained negative (AUC = 1.000; p < 0.0001, 95% CI = 0.872–1.000, Youden index J = 1.000, NPV = 100%, PPV = 100%). (F) ROC analysis for the triplex-probe qPCR reached 100% specificity, and 88% sensitivity as 22 specimens were positive (13 FFPE samples and 9 fresh clinical samples) and only 3 samples were false-negative (AUC = 0.940 ± 0.03; p < 0.0001, 95% CI = 0.823–0.989, Youden index J = 0.8800, NPV = 85.7%, PPV = 100%). The 18 non-PCM samples remained negative. The results are typical of two separate experiments.
All fresh tissue samples tested positive for at least three bands using the GAPDH control (Supplementary Figure S2). From the nine samples of the PCM group, four samples were positive for the P. brasiliensis complex probe (Cq = 29.99–36.78; Median Cq = 35.84) and five samples for P. lutzii probe (Cq = 9.03–37.07; Median Cq = 22.05). All nine samples were positive for the Paracoccidioides probe (Cq = 11.70–37.62; Median Cq = 34.56, Figure 6B). For the non-PCM group, the 18 samples tested negative for the 3 probes evaluated (Supplementary Figure S2). Sensitivity (100%), specificity (100%), the area under the ROC curve (1.000; p < 0.0001), Youden index (1.000), and negative (100%) and positive predictive values (100%) of qPCR were calculated using the results obtained with 9 clinical specimens of diseased patients and 18 samples of other diseases (95% CI = 0.872–1.000) (Figure 6E).
Using kappa statistics, a comparison between qPCR and a conventional duplex PCR result for FFPE or fresh clinical samples revealed significant agreement (i.e., 13 detectable and 3 undetectable for FFPE samples, and 9 detectable for fresh clinical samples; kappa = 1.0; see Supplementary Figure S2).
3.8. Detection of Paracoccidioides DNA from Spiked Soil Samples
The sensitivity of our assay to detect Paracoccidioides DNA was evaluated using soil samples spiked with P. brasiliensis or P. lutzii DNAs. The amplification curves indicated positive signals from spiked soil material, and the Cq values correlated with the input amount of DNA. The minimum detection level for Paracoccidioides in soil for all probes was 10 fg of gDNA (Figure 7). There was no amplification beyond the cycle threshold in the negative control samples, which included sterile, non-spiked soil (no false positives). This suggests that our qPCR can specifically detect DNA from Paracoccidioides, demonstrating its high specificity and potential use in environmental research.
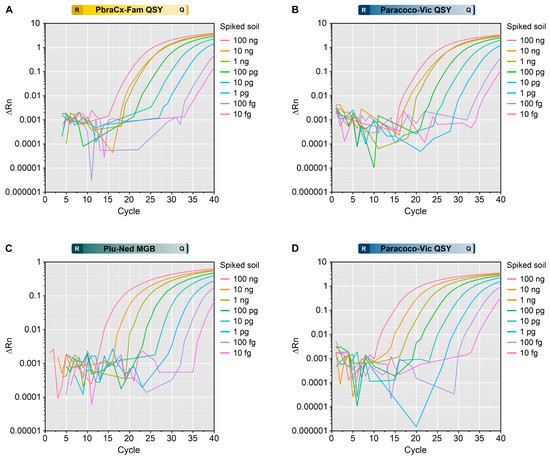
Figure 7.
Assessment of a triplex-probe qPCR to detect (A,B) P. brasiliensis or (C,D) P. lutzii DNA in the spiked soil. The amplification plot for the PbraCx-Fam probe (A) or Paracoco-Vic probe (B) and the DNA from Pb18. The amplification plot for the Plu-Ned probe (C) or Paracoco-Vic probe (D) and the DNA from Pb01. Different colors of amplification curves represent samples from soil spiked with different inputs of Paracoccidioides DNA (100 ng–10 fg). ΔRn relates to the magnitude of the normalized reporter value (fluorescence signal) used to construct the standard curve. The results are typical of two separate experiments. R: reporter; Q: quencher.
4. Discussion
Infections caused by fungi threaten human health worldwide, and presumably, millions of people live with severe fungal infections [45,46,47]. As observed for PCM, a neglected mycosis with significant morbidity and mortality in Latin America, many lives could be preserved by applying quick and accurate diagnosis [48]. Despite PCM clinical relevance, diagnostics tools have been challenging, mainly because classical techniques such as culturing, histopathology, and phenotypic methods are laborious and time-consuming. We developed a triplex-probe assay targeting the Paracoccidioides rDNA region (ITS2/28S) to overcome this problem of detecting DNA from isolated cultures to clinical samples. Our qPCR assay is an important tool for understanding epidemiological patterns, mapping the affected areas, and guiding early treatment of patients affected by the disease [49].
Our multiplex qPCR presents the following improvements: (i) it simplifies the differentiation of P. lutzii from other members of the P. brasiliensis complex; (ii) it recognizes the main culprits of classic PCM in a one-tube reaction, thus saving qPCR components and the steps required to distinguish Paracoccidioides; (iii) it is more sensitive than isolation in culture [5] and is at least 100 times more sensitive than our previous duplex PCR [33]; (iv) it is ideal for detecting Paracoccidioides in fresh specimens or FFPE tissue samples and exhibits no cross-reaction with human or murine DNA; (v) it is suitable for the detection of Paracoccidioides in environmental samples; and (vi) it is straightforward to interpret (Supplementary Figure S3), expanding the offer of a highly effective assay to distant areas in South America, a critical bottleneck in a neglected tropical disease scenario.
The species boundaries in Paracoccidioides impact the development of diagnostic tests [49]. Thus, despite good PCR-based tools available in the literature [4,10,11,50,51,52,53], our assay has the advantage of covering the recent taxonomic developments in the genus [1,4,17,18,54]. The P. brasiliensis complex comprises cryptic species accumulating genetic differences without adding significant clinical relevance [1]. Recognizing members of the P. brasiliensis complex down to the species level requires laborious approaches such as MLSA schemes [1,18] or whole-genome sequencing [19,20,55], a scenario distant from the reality of clinical laboratories in South America, where PCM is endemic [56].
From a serological perspective, it is not uncommon to find patients with negative serological results for the B-339 antigen in double immunodiffusion assay but positive in direct mycological examination [5,57,58,59,60,61]. In these paradoxical cases, identifying P. lutzii may be essential to recognizing false negative cases in serology and mapping the areas where regional antigens from P. lutzii are needed. To our knowledge, qPCR assays that allow the identification of all agents embedded in the P. brasiliensis complex are inexistent due to the technical complexity of multiplexing more than three DNA-based FRET probes labeled with fluorescent dyes in addition to the unreasonable cost generated by this experimental design. Moreover, judging from the clinical perspective, speciating members of the P. brasiliensis complex has no impact on patient management [5,6,8,62,63,64]. Therefore, differentiating between members of the P. brasiliensis complex and P. lutzii seems to be, at this moment, the safest and most cost-effective strategy for diagnosing classic PCM [1,33,49].
Our qPCR test can be used in different formats according to the needs and budget of each laboratory. For example, the singleplex assay employing only the Paracoco-Vic probe grants the genus-level diagnosis of classic PCM (Supplementary Figure S3). The duplex assay with the PbraCx-Fam and Plu-Ned probes allows the diagnosis of mixed infections by any of the members of the P. brasiliensis complex and P. lutzii (Supplementary Figure S3). However, similar to our previous conventional duplex PCR assay targeting the GP43 gene [33], here it is impossible to detect mixed infections by any two members of the P. brasiliensis complex (e.g., P. americana and P. brasiliensis s. str., or P. restrepiensis and P. venezuelensis) since a single positive signal would be detected (i.e., PbraCx-Fam). From our perspective, the triplex assay is the most attractive option because it offers a double confirmation: first, generic (Paracoccidioides spp.) and second, specific, which allows a significant coverage of the diversity of Paracoccidioides, bringing more confidence to the assay (Supplementary Figure S3).
The high sensitivity and specificity of our qPCR assay results from combining accurate genome-based design [20,55] and using fluorescently-labeled TaqMan probes. As for efficiency, we showed great linearity values (R2 nearly 1.0) and excellent slopes (~−3.33) for all curves. The amplification scale ranges from 1.0 (no amplification) to 2.0 (perfect exponential duplication), with scales ranging between 1.8 and 2.0 accepted as good [65], which is the case of all amplification results we had with a single-to-multiplex assay. These results reinforce that the presence of more than one probe in a single-round reaction does not reduce the efficiency of our assay. The number of isolates used here allowed us to verify that the AUC coincided with the total area of the ROC curve, and we assumed nearly 100% sensitivity and specificity in any format employed. The analytical sensitivity for gDNA was 10 fg/μL for gDNA, and using the ITS2 amplicon, we could detect up to three copies, according to Poisson’s distribution probability [6,66].
Sputum and tissue biopsies, including FFPE tissues, are the main clinical specimens used to diagnose classic PCM. A great performance of our test was observed with clinical samples, reducing the need for isolation of Paracoccidioides, thus saving critical time. Overall, the application of PCR-based diagnostics shows that the test might work better on sputum than in blood samples [50], whereas the tissue detection would have no problem if the available Paracoccidioides species’ DNA is greater than 10 fg [67], which is in agreement with our results. Therefore, we recommend detecting Paracoccidioides DNA in clinical routine and emphasize that the major feature of maintaining diagnostic accuracy is acquiring an excellent clinical specimen free of PCR inhibitors [49]. Moreover, judging from the retrospective nature of our study, we report a significant occurrence of P. lutzii as the etiological agent in areas remote from the epicenter of P. lutzii-mycosis in Central-West Brazil, confirming that Paracoccidioides species are pathogens on the move and epidemiological trends are not fully elucidated [9,15,68,69,70].
FFPE samples have been effectively applied in DNA sequencing and qPCR investigations [38,42], but this success is affected by several pre- and postprocessing issues. For example, downstream molecular analyses may be impacted by the nature and quantity of tissue, the fixative buffer employed for tissue conservation, the extent of fixation, the age of the paraffin block, and storage conditions, as well as the cross-links, the length of the chosen DNA segment to be amplified, and copy number variation [39,40,41]. We included FFPE specimens preserved for a decade (2010–2020) that underwent rigorous quality control by running a quadruplex assay targeting the human GAPDH gene [34]. A remarkable positivity was reported (81.2%) for our triplex-probe qPCR assay, underlining the potential for retrospective studies in FFPE tissue collections in Latin America. Moreover, the odds of detection were improved by choosing a target in the genic and spacer regions of the ribosomal cistron with significant copy number variation in Paracoccidioides genomes. Considering the natural fragmentation of DNA during the processing of FFPE tissue (cross-links), the smaller the size of the amplicon used for qPCR analysis, the greater the chance of detecting the specific target, and we report great success in amplifying a fragment of 137–143 bp in FFPE tissues.
A major pitfall of our study is the absence of clinical samples from patients with PCM loboi or cetaceans with PCM ceti [12,26,71], which is partially justified due to the unculturable nature of the etiological agents and the rarity of these infections compared with classic PCM [1,8]. A BLAST search using the Primer-BLAST tool and the sequences of Paracoco-F and Paracoco-R primers or probes did not retrieve any amplicon related to P. loboi, which could be associated with the scarcity of P. loboi rDNA sequences from public databases [7]. Judging from P. cetii sequences (GenBank accession numbers: MW566081–MW566084), a mutation in the forward primer and along the PbraCx-Fam (1 mutation out of 29; 3.44%) and Plu-Ned (10 mutations out of 18; 55.55%) probes binding regions may prevent recognition using our assay. The Paracoccidioides probe (Paracoco-Vic), on the other hand, had a 100% similarity with P. cetii, but not P. loboi (13 mutations out of 19; 68.42%). This strongly suggests that none of the probe and primer combinations would recognize P. cetii and P. loboi (Supplementary Figure S1).
Ecological studies on PCM present an important bottleneck due to the insufficient detection capacity of Paracoccidioides in its natural habitat, either by employing classical methods such as fungal isolation or molecular tools based on DNA detection [51,72,73,74,75,76,77]. Here, we demonstrated that our triplex-probe qPCR has the potential to identify Paracoccidioides in ecological studies using soil samples, the typical environmental sources of propagules [10,51,53,74,75,78]. Isolation of Paracoccidioides species from the environment is infrequently reported in endemic areas [76,77,79,80], which could be associated with the low concentration and seasonality of Paracoccidioides propagules in the soil or the fastidious development of Paracoccidioides species in nutrient-rich culture media compared to the rapidly growing Ascomycetes that lead soil diversity and abundance globally [81,82]. Recently, Arantes et al., using nested PCR and in situ hybridization methods, demonstrated that soil and aerosol samples were positive for Paracoccidioides spp. DNA, revealing the ordinary distribution of these microorganisms in the Brazilian territory [78]. Likewise, our qPCR assay employing spiked samples revealed the potential to gain insights into the distributions of Paracoccidioides and the conditions that boost their occurrence in the environment.
5. Conclusions
Diagnostics is a vital component of any successful public health policy, and the development of fast, reliable, and reasonably priced assays is critical for tailoring proper therapeutical strategies against PCM. Thus, the approach presented here simplifies laboratory diagnosis without delving into speciation processes, for which molecular markers are not yet defined and are still nebulous for routine use [1]. Our test fits current diagnostics necessities as it considerably reduces the turnaround time for fungal identification, dismisses the need for gel electrophoresis and downstream analyses, and reduces amplicon contamination and the manipulation of samples and cultures, which is a cost-effective choice for a neglected tropical disease associated with poverty. Moreover, our assay allows us to investigate eventual co-infections, which might be less rare than previously thought [33,42]. All these features are desirable to improve fungal diagnostic capacity and tackle the spread of classic PCM in a vast area of the Americas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9030358/s1, Supplementary Table S1: Paracoccidioides isolates used in this study. Supplementary Table S2: Genomes used in this study for primer design. Supplementary Table S3: DNA samples derived from non-target species, including agents of superficial, subcutaneous, and systemic mycosis in humans and animals. Supplementary Table S4: List of formalin-fixed paraffin-embedded (FFPE) tissue and other biological samples included in the analysis. Supplementary Table S5: Primer-BLAST Results (https://www.ncbi.nlm.nih.gov/tools/primer-blast/ Accessed on 2 January 2023; 13:50 h GMT). Supplementary Figure S1: Alignment of the target region (partial ITS2/28S) for all members of the genus Paracoccidioides. The primers (gray boxes) and the probes PbraCx-Fam (yellow box), Plu-Ned (green box), and Paracoco-Vic (blue box) are annotated in the figure. The percentage of conservation and the consensus sequence appears at the alignment’s bottom. In silico analyses show that the primers and probes were designed in highly conserved regions for their respective targets. However, the assay is incompatible with previously published P. cetii and P. loboi sequences. Supplementary Figure S2: STARD flow diagram of the study population for the (A) formalin-fixed, paraffin-embedded (FFPE) tissue samples and (B) fresh specimens. As a quality control, all DNAs extracted from FFPE samples or fresh clinical specimens were evaluated via PCR using a quadruplex assay targeting nonoverlapping sites in the GAPDH gene (chr12), and samples that yielded 100, 200, 300, and 400 bp amplicons were regarded to be free of PCR inhibitors. Afterward, high-quality DNA samples were subjected to the triplex probe-based qPCR assay and the conventional duplex PCR assay previously described by Pinheiro et al. Supplementary Figure S3: Simple interpretation of qPCR results for singleplex- (orange panel), duplex- (blue panel), and triplex-probe qPCR assays. Regardless of the etiological agent, the Paracoco-Vic probe allows only a generic identification and does not allow the investigation of mixed infections. On the other hand, the duplex assay (PbraCx-Fam and Plu-Ned probes) permits the differentiation of the etiological agents of the P. brasiliensis complex and P. lutzii, thus allowing the discrimination of mixed infections caused by P. brasiliensis complex and P. lutzii.
Author Contributions
Conceptualization, A.M.R.; methodology, B.G.P., A.P.P., G.R., A.S.N. and A.M.R.; validation, A.M.R. and B.G.P.; formal analysis, A.M.R. and B.G.P.; investigation, B.G.P., A.P.P. and A.M.R.; resources, A.M.R., R.C.H. and Z.P.d.C.; data curation, A.M.R. and B.G.P.; writing—original draft preparation, A.M.R. and B.G.P.; writing—review and editing, A.M.R. and F.H.; visualization, A.M.R., B.G.P., A.P.P., F.H., R.C.H. and Z.P.d.C.; supervision, A.M.R.; project administration, A.M.R.; funding acquisition, A.M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (2017/27265-5 and 2018/21460-3) http://www.fapesp.br/, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 429594/2018-6) http://www.cnpq.br/, and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES 88887.177846/2018-00) https://www.capes.gov.br/. AMR is a CNPq Research Productivity Fellow (CNPq 304902/2020-9).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of the Federal University of São Paulo (protocol codes 9771060120 and 3147220120; 20 January 2020, and 17 February 2020).
Informed Consent Statement
Informed consent was waived because of the study’s retrospective nature, and the analysis used anonymous clinical data.
Data Availability Statement
The data presented in this study are available within the article and supplementary material.
Acknowledgments
We thank Sigrid de Sousa dos Santos from the Department of Medicine, Federal University of São Carlos, São Carlos, SP, Brazil, and Iria Visoná from the Department of Pathology, Federal University of São Paulo, São Paulo, SP, Brazil for providing some tissue sections samples (FFPE).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Rodrigues, A.M.; Hagen, F.; Puccia, R.; Hahn, R.C.; de Camargo, Z.P. Paracoccidioides and paracoccidioidomycosis in the 21st century. Mycopathologia, 2023, in press. [CrossRef]
- Kidd, S.E.; Abdolrasouli, A.; Hagen, F. Fungal nomenclature: Managing change is the name of the game. Open Forum Infect. Dis. 2023, 10, ofac559. [Google Scholar] [CrossRef] [PubMed]
- Carrero, L.L.; Nino-Vega, G.; Teixeira, M.M.; Carvalho, M.J.; Soares, C.M.; Pereira, M.; Jesuino, R.S.; McEwen, J.G.; Mendoza, L.; Taylor, J.W.; et al. New Paracoccidioides brasiliensis isolate reveals unexpected genomic variability in this human pathogen. Fungal Genet. Biol. 2008, 45, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.A.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet. Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Hahn, R.C.; Rodrigues, A.M.; Della Terra, P.P.; Nery, A.F.; Hoffmann-Santos, H.D.; Góis, H.M.; Fontes, C.J.; de Camargo, Z.P. Clinical and epidemiological features of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2019, 13, e0007437. [Google Scholar] [CrossRef] [PubMed]
- de Macedo, P.M.; Teixeira, M.D.M.; Barker, B.M.; Zancopé-Oliveira, R.M.; Almeida-Paes, R.; Francesconi do Valle, A.C. Clinical features and genetic background of the sympatric species Paracoccidioides brasiliensis and Paracoccidioides americana. PLoS Negl. Trop. Dis. 2019, 13, e0007309. [Google Scholar] [CrossRef]
- Vilela, R.; Huebner, M.; Vilela, C.; Vilela, G.; Pettersen, B.; Oliveira, C.; Mendoza, L. The taxonomy of two uncultivated fungal mammalian pathogens is revealed through phylogeny and population genetic analyses. Sci. Rep. 2021, 11, 18119. [Google Scholar] [CrossRef]
- Hahn, R.C.; Hagen, F.; Mendes, R.P.; Burger, E.; Nery, A.F.; Siqueira, N.P.; Guevara, A.; Rodrigues, A.M.; de Camargo, Z.P. Paracoccidioidomycosis: Current status and future trends. Clin. Microbiol. Rev. 2022, 35, e0023321. [Google Scholar] [CrossRef]
- Roberto, T.N.; De Carvalho, J.A.; Beale, M.A.; Hagen, F.; Fisher, M.C.; Hahn, R.C.; de Camargo, Z.P.; Rodrigues, A.M. Exploring genetic diversity, population structure, and phylogeography in Paracoccidioides species using AFLP markers. Stud. Mycol. 2021, 100, 100131. [Google Scholar] [CrossRef]
- Theodoro, R.C.; Teixeira, M.D.M.; Felipe, M.S.S.; Paduan, K.D.S.; Ribolla, P.M.; San-Blas, G.; Bagagli, E. Genus Paracoccidioides: Species recognition and biogeographic aspects. PLoS ONE 2012, 7, e37694. [Google Scholar] [CrossRef] [PubMed]
- Theodoro, R.C.; Bagagli, E.; Oliveira, C. Phylogenetic analysis of PRP8 intein in Paracoccidioides brasiliensis species complex. Fungal Genet. Biol. 2008, 45, 1284–1291. [Google Scholar] [CrossRef]
- Gonçalves, F.G.; Rosa, P.S.; Belone, A.F.F.; Carneiro, L.B.; de Barros, V.L.Q.; Bispo, R.F.; Sbardelott, Y.; Neves, S.; Vittor, A.Y.; Woods, W.J.; et al. Lobomycosis epidemiology and management: The quest for a cure for the most neglected of neglected tropical diseases. J. Fungi 2022, 8, 494. [Google Scholar] [CrossRef]
- Camargo, Z.P.; Rodrigues, A.M. Paracoccidioides complex. In Pocket Guide to Mycological Diagnosis; Cordeiro, R.D.A., Yuan, L., Eds.; Pocket Guides to Biomedical Sciences; CRC Press: Boca Raton, FL, USA, 2019; Volume 78, pp. 125–134. [Google Scholar]
- Gomes, G.M.; Cisalpino, P.S.; Taborda, C.P.; de Camargo, Z.P. PCR for diagnosis of paracoccidioidomycosis. J. Clin. Microbiol. 2000, 38, 3478–3480. [Google Scholar] [CrossRef] [PubMed]
- Roberto, T.N.; Rodrigues, A.M.; Hahn, R.C.; de Camargo, Z.P. Identifying Paracoccidioides phylogenetic species by PCR-RFLP of the alpha-tubulin gene. Med. Mycol. 2016, 54, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Matute, D.R.; Sepulveda, V.E.; Quesada, L.M.; Goldman, G.H.; Taylor, J.W.; Restrepo, A.; McEwen, J.G. Microsatellite analysis of three phylogenetic species of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2006, 44, 2153–2157. [Google Scholar] [CrossRef]
- Matute, D.R.; McEwen, J.G.; Puccia, R.; Montes, B.A.; San-Blas, G.; Bagagli, E.; Rauscher, J.T.; Restrepo, A.; Morais, F.; Niño-Vega, G.; et al. Cryptic speciation and recombination in the fungus Paracoccidioides brasiliensis as revealed by gene genealogies. Mol. Biol. Evol. 2006, 23, 65–73. [Google Scholar] [CrossRef]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef]
- Teixeira, M.D.M.; Cattana, M.E.; Matute, D.R.; Muñoz, J.F.; Arechavala, A.; Isbell, K.; Schipper, R.; Santiso, G.; Tracogna, F.; Sosa, M.D.L.Á.; et al. Genomic diversity of the human pathogen Paracoccidioides across the South American continent. Fungal Genet. Biol. 2020, 140, 103395. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.A.; et al. Genome diversity, recombination, and virulence across the major lineages of Paracoccidioides. mSphere 2016, 1, e00213-16. [Google Scholar] [CrossRef] [PubMed]
- Fava-Netto, C. Contribuição para o estudo imunológico da blastomicose de Lutz (blastomicose sul-americana). Rev. Inst. Adolfo Lutz 1961, 21, 99–194. [Google Scholar]
- Fava-Netto, C.; Vegas, V.S.; Sciannamea, I.M.; Guarnieri, D.B. The polysaccharidic antigen from Paracoccidioides brasiliensis. Study of the time of cultivation necessary for the preparation of the antigen. Rev. Inst. Med. Trop. São Paulo 1969, 11, 177–181. [Google Scholar]
- Rodrigues, A.M.; Beale, M.A.; Hagen, F.; Fisher, M.C.; Terra, P.P.D.; de Hoog, S.; Brilhante, R.S.N.; de Aguiar Cordeiro, R.; de Souza Collares Maia Castelo-Branco, D.; Rocha, M.F.G.; et al. The global epidemiology of emerging Histoplasma species in recent years. Stud. Mycol. 2020, 97, 100095. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Morais, F.V.; Barros, T.F.; Fukada, M.K.; Cisalpino, P.S.; Puccia, R. Polymorphism in the gene coding for the immunodominant antigen gp43 from the pathogenic fungus Paracoccidioides brasiliensis. J. Clin. Microbiol. 2000, 38, 3960. [Google Scholar] [CrossRef] [PubMed]
- Vilela, R.; Rosa, P.S.; Belone, A.F.; Taylor, J.W.; Diorio, S.M.; Mendoza, L. Molecular phylogeny of animal pathogen Lacazia loboi inferred from rDNA and DNA coding sequences. Mycol. Res. 2009, 113, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; de Hoog, G.S.; de Camargo, Z.P. Molecular diagnosis of pathogenic Sporothrix species. PLoS Negl. Trop. Dis. 2015, 9, e0004190. [Google Scholar] [CrossRef]
- Pinheiro, B.G.; Pôssa, A.P.; Della Terra, P.P.D.; de Carvalho, J.A.D.; Ricci, G.; Nishikaku, A.S.; Hahn, R.C.; Camargo, Z.P.D.; Rodrigues, A.M. A new duplex PCR-assay for the detection and identification of Paracoccidioides species. J. Fungi 2021, 7, 169. [Google Scholar] [CrossRef]
- van Beers, E.H.; Joosse, S.A.; Ligtenberg, M.J.; Fles, R.; Hogervorst, F.B.L.; Verhoef, S.; Nederlof, P.M. A multiplex PCR predictor for aCGH success of FFPE samples. Br. J. Cancer 2006, 94, 333–337. [Google Scholar] [CrossRef]
- Pahl, A.; Kuhlbrandt, U.; Brune, K.; Rollinghoff, M.; Gessner, A. Quantitative detection of Borrelia burgdorferi by real-time PCR. J. Clin. Microbiol. 1999, 37, 1958–1963. [Google Scholar] [CrossRef] [PubMed]
- Della Terra, P.P.; Gonsales, F.F.; de Carvalho, J.A.; Hagen, F.; Kano, R.; Bonifaz, A.; Camargo, Z.P.; Rodrigues, A.M. Development and evaluation of a multiplex qPCR assay for rapid diagnostics of emerging sporotrichosis. Transbound. Emerg. Dis. 2022, 69, e704–e716. [Google Scholar] [CrossRef] [PubMed]
- Bialek, R.; Feucht, A.; Aepinus, C.; Just-Nübling, G.; Robertson, V.J.; Knobloch, J.; Hohle, R. Evaluation of two nested PCR assays for detection of Histoplasma capsulatum DNA in human tissue. J. Clin. Microbiol. 2002, 40, 1644–1647. [Google Scholar] [CrossRef]
- Ricci, G.; Zelck, U.; Mota, F.; Lass-Florl, C.; Franco, M.F.; Bialek, R. Genotyping of Paracoccidioides brasiliensis directly from paraffin embedded tissue. Med. Mycol. 2008, 46, 31–34. [Google Scholar] [CrossRef]
- Cho, M.; Ahn, S.; Hong, M.; Bang, H.; Van Vrancken, M.; Kim, S.; Lee, J.; Park, S.H.; Park, J.O.; Park, Y.S.; et al. Tissue recommendations for precision cancer therapy using next generation sequencing: A comprehensive single cancer center’s experiences. Oncotarget 2017, 8, 42478–42486. [Google Scholar] [CrossRef]
- Blow, N. Tissue preparation: Tissue issues. Nature 2007, 448, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Rickerts, V. Identification of fungal pathogens in Formalin-fixed, Paraffin-embedded tissue samples by molecular methods. Fungal Biol. 2016, 120, 279–287. [Google Scholar] [CrossRef]
- Ricci, G.; Campanini, E.B.; Nishikaku, A.S.; Puccia, R.; Marques, M.; Bialek, R.; Rodrigues, A.M.; Batista, W.L. PbGP43 genotyping using paraffin-embedded biopsies of human paracoccidioidomycosis reveals a genetically distinct lineage in the Paracoccidioides brasiliensis complex. Mycopathologia 2022, 187, 157–168. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Najafzadeh, M.J.; de Hoog, G.S.; de Camargo, Z.P. Rapid identification of emerging human-pathogenic Sporothrix species with rolling circle amplification. Front. Microbiol. 2015, 6, 1385. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G. Practical Statistics for Medical Research; Chapman and Hall: London, UK, 1991; p. 624. [Google Scholar]
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef]
- Bongomin, F.; Gago, S.; Oladele, R.; Denning, D. Global and multi-national prevalence of fungal diseases—Estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Coutinho, Z.F.; Wanke, B.; Travassos, C.; Oliveira, R.M.; Xavier, D.R.; Coimbra, C.E., Jr. Hospital morbidity due to paracoccidioidomycosis in Brazil (1998–2006). Trop. Med. Int. Health 2015, 20, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, B.G.; Hahn, R.C.; Camargo, Z.P.; Rodrigues, A.M. Molecular tools for detection and identification of Paracoccidioides species: Current status and future perspectives. J. Fungi 2020, 6, 293. [Google Scholar] [CrossRef] [PubMed]
- Buitrago, M.J.; Merino, P.; Puente, S.; Gomez-Lopez, A.; Arribi, A.; Zancope-Oliveira, R.M.; Gutierrez, M.C.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Utility of real-time PCR for the detection of Paracoccidioides brasiliensis DNA in the diagnosis of imported paracoccidioidomycosis. Med. Mycol. 2009, 47, 879–882. [Google Scholar] [CrossRef][Green Version]
- Theodoro, R.C.; Candeias, J.M.; Araujo, J.P., Jr.; Bosco Sde, M.; Macoris, S.A.; Padula, L.O.; Franco, M.; Bagagli, E. Molecular detection of Paracoccidioides brasiliensis in soil. Med. Mycol. 2005, 43, 725–729. [Google Scholar] [CrossRef][Green Version]
- Arantes, T.D.; Theodoro, R.C.; Teixeira, M.M.; Bagagli, E. Use of fluorescent oligonucleotide probes for differentiation between Paracoccidioides brasiliensis and Paracoccidioides lutzii in yeast and mycelial phase. Mem. Inst. Oswaldo Cruz 2017, 112, 140–145. [Google Scholar] [CrossRef]
- Arantes, T.D.; Theodoro, R.C.; Da Graça Macoris, S.A.; Bagagli, E. Detection of Paracoccidioides spp. in environmental aerosol samples. Med. Mycol. 2013, 51, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Theodoro, R.C.; Oliveira, F.F.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef]
- Munoz, J.F.; Gallo, J.E.; Misas, E.; Priest, M.; Imamovic, A.; Young, S.; Zeng, Q.; Clay, O.K.; McEwen, J.G.; Cuomo, C.A. Genome update of the dimorphic human pathogenic fungi causing paracoccidioidomycosis. PLoS Negl. Trop. Dis. 2014, 8, e3348. [Google Scholar] [CrossRef]
- Griffiths, J.; Lopes Colombo, A.; Denning, D.W. The case for paracoccidioidomycosis to be accepted as a neglected tropical (fungal) disease. PLoS Negl. Trop. Dis. 2019, 13, e0007195. [Google Scholar] [CrossRef] [PubMed]
- Maifrede, S.B.; Kruschewsky, W.L.L.; Patrício, S.A.; Falqueto, A.; Peçanha, P.M.; Malaquias, L.C.C.; Pôssa, A.P.; de Camargo, Z.P.; Rodrigues, A.M.; Gonçalves, S.S.; et al. Screening paracoccidioidomycosis by double immunodiffusion test in a referred diagnostic center in Brazilian southeastern: An accessible tool. Infection 2021, 49, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Kubitschek-Barreira, P.H.; Pinheiro, B.G.; Teixeira-Ferreira, A.; Hahn, R.C.; de Camargo, Z.P. Immunoproteomic analysis reveals novel candidate antigens for the diagnosis of paracoccidioidomycosis due to Paracoccidioides lutzii. J. Fungi 2020, 6, 357. [Google Scholar] [CrossRef]
- Dos Santos, P.O.; Rodrigues, A.M.; Fernandes, G.F.; da Silva, S.H.; Burger, E.; de Camargo, Z.P. Immunodiagnosis of paracoccidioidomycosis due to Paracoccidioides brasiliensis using a latex test: Detection of specific antibody anti-gp43 and specific antigen gp43. PLoS Negl. Trop. Dis. 2015, 9, e0003516. [Google Scholar] [CrossRef]
- Queiroz Junior, L.P.; de Camargo, Z.P.; Tadano, T.; Rodrigues, A.M.; Takarara, D.T.; Gegembauer, G.; Araujo, L.M.; Hahn, R.C. Serological and antigenic profiles of clinical isolates of Paracoccidioides spp. from Central Western Brazil. Mycoses 2014, 57, 466–472. [Google Scholar] [CrossRef]
- Gegembauer, G.; Araujo, L.M.; Pereira, E.F.; Rodrigues, A.M.; Paniago, A.M.; Hahn, R.C.; de Camargo, Z.P. Serology of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2014, 8, e2986. [Google Scholar] [CrossRef] [PubMed]
- Nery, A.F.; de Camargo, Z.P.; Rodrigues, A.M.; Portela, T.F.; Hoffmann-Santos, H.D.; Dambros, P.V.K.; de Souza, J.F.R.; Garcia, A.C.; Santos, C.A.D.; Hagen, F.; et al. Paracoccidioidomycosis due to P. lutzii: The importance of neutrophil/lymphocyte ratio in the symptomatic and asymptomatic phases in severe cases. Mycoses 2021, 64, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Nery, A.F.; de Camargo, Z.P.; Rodrigues, A.M.; Portela, T.F.; Hoffmann-Santos, H.D.; Pinheiro, B.G.; Possa, A.P.; Cavalcante, L.; Hagen, F.; Hahn, R.C. Puzzling paracoccidioidomycosis: Factors associated with the severity of Paracoccidioides lutzii infections. Int. J. Infect. Dis. 2021, 107, 284–290. [Google Scholar] [CrossRef]
- Pereira, E.F.; Gegembauer, G.; Chang, M.R.; Camargo, Z.P.D.; Nunes, T.F.; Ribeiro, S.M.; Carvalho, L.R.D.; Maldonado, B.M.; Mendes, R.P.; Paniago, A.M.M. Comparison of clinico-epidemiological and radiological features in paracoccidioidomycosis patients regarding serological classification using antigens from Paracoccidioides brasiliensis complex and Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2020, 14, e0008485. [Google Scholar] [CrossRef]
- Imai, T.; Sano, A.; Mikami, Y.; Watanabe, K.; Aoki, F.H.; Branchini, M.L.; Negroni, R.; Nishimura, K.; Miyaji, M. A new PCR primer for the identification of Paracoccidioides brasiliensis based on rRNA sequences coding the internal transcribed spacers (ITS) and 5.8S regions. Med. Mycol. 2000, 38, 323–326. [Google Scholar] [CrossRef][Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum Information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Rocha-Silva, F.; Gomes, L.I.; Gracielle-Melo, C.; Goes, A.M.; Caligiorne, R.B. Real Time Polymerase Chain Reaction (rt-PCR): A New Patent to Diagnostic Purposes for Paracoccidioidomycosis. Recent Pat. Endocr. Metab. Immune Drug Discov. 2017, 10, 143–149. [Google Scholar] [CrossRef]
- Cocio, T.A.; Nascimento, E.; von Zeska Kress, M.R.; Bagagli, E.; Martinez, R. Phylogenetic species of Paracoccidioides spp. isolated from clinical and environmental samples in a hyperendemic area of paracoccidioidomycosis in Southeastern Brazil. J. Fungi 2020, 6, 132. [Google Scholar] [CrossRef]
- Cocio, T.A.; Nascimento, E.; Kress, M.R.; Bagagli, E.; Martinez, R. Characterization of a Paracoccidioides spp. strain from southeastern Brazil genotyped as Paracoccidioides restrepiensis (PS3) and review of this phylogenetic species. Genet. Mol. Biol. 2020, 43, e20190201. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R. New trends in paracoccidioidomycosis epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Vilela, R.; Mendoza, L.; Rosa, P.S.; Belone, A.F.F.; Madeira, S.; Opromolla, D.V.A.; de Resende, M.A. Molecular model for studying the uncultivated fungal pathogen Lacazia loboi. J. Clin. Microbiol. 2005, 43, 3657–3661. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.; Bagagli, E.; Scapolio, S.; Da Silva Lacaz, C. A critical analysis of isolation of Paracoccidioides brasiliensis from soil. Med. Mycol. 2000, 38, 185–191. [Google Scholar] [CrossRef]
- Bagagli, E.; Sano, A.; Coelho, K.I.; Alquati, S.; Miyaji, M.; de Camargo, Z.P.; Gomes, G.M.; Franco, M.; Montenegro, M.R. Isolation of Paracoccidioides brasiliensis from armadillos (Dasypus noveminctus) captured in an endemic area of paracoccidioidomycosis. Am. J. Trop. Med. Hyg. 1998, 58, 505–512. [Google Scholar] [CrossRef]
- Bagagli, E.; Theodoro, R.C.; Bosco, S.M.; McEwen, J.G. Paracoccidioides brasiliensis: Phylogenetic and ecological aspects. Mycopathologia 2008, 165, 197–207. [Google Scholar] [CrossRef][Green Version]
- Richini-Pereira, V.B.; Bosco Sde, M.; Theodoro, R.C.; Macoris, S.A.; Bagagli, E. Molecular approaches for eco-epidemiological studies of Paracoccidioides brasiliensis. Mem. Inst. Oswaldo Cruz 2009, 104, 636–643. [Google Scholar] [CrossRef]
- Silva-Vergara, M.L.; Martinez, R.; Camargo, Z.P.; Malta, M.H.; Maffei, C.M.; Chadu, J.B. Isolation of Paracoccidioides brasiliensis from armadillos (Dasypus novemcinctus) in an area where the fungus was recently isolated from soil. Med. Mycol. 2000, 38, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Silva-Vergara, M.L.; Martinez, R.; Chadu, A.; Madeira, M.; Freitas-Silva, G.; Leite Maffei, C.M. Isolation of a Paracoccidioides brasiliensis strain from the soil of a coffee plantation in Ibia, State of Minas Gerais, Brazil. Med. Mycol. 1998, 36, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Arantes, T.D.; Theodoro, R.C.; Teixeira, M.D.M.; Bosco, S.D.M.G.; Bagagli, E. Environmental mapping of Paracoccidioides spp. in Brazil reveals new clues into genetic diversity, biogeography and wild host association. PLoS Negl. Trop. Dis. 2016, 10, e0004606. [Google Scholar] [CrossRef]
- Ferreira, M.S.; Freitas, L.H.; Lacaz Cda, S.; del Negro, G.M.; de Melo, N.T.; Garcia, N.M.; de Assis, C.M.; Salebian, A.; Heins-Vaccari, E.M. Isolation and characterization of a Paracoccidioides brasiliensis strain from a dogfood probably contaminated with soil in Uberlandia, Brazil. J. Med. Vet. Mycol. 1990, 28, 253–256. [Google Scholar] [CrossRef]
- De Albornoz, M.B. Isolation of Paracoccidioides brasiliensis from rural soil in Venezuela. Sabouraudia 1971, 9, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef]
- Guarro, J.; Gene, J.; Stchigel, A.M.; Figueras, M.J. Atlas of Soil Ascomycetes; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012; p. 486. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).