
Alternaria alternata, the Causal Agent of a New Needle Blight Disease on Pinus bungeana
Abstract
1. Introduction
2. Materials and Methods
2.1. Disease Investigation and Isolate Collection
2.2. Pathogenicity Tests
2.3. Morphological Study
2.4. DNA Extraction and Polymerase Chain Reaction (PCR)
2.5. DNA Sequencing and Phylogenetic Analysis
3. Results
3.1. Symptoms in Nature
3.2. Pathogenicity Tests
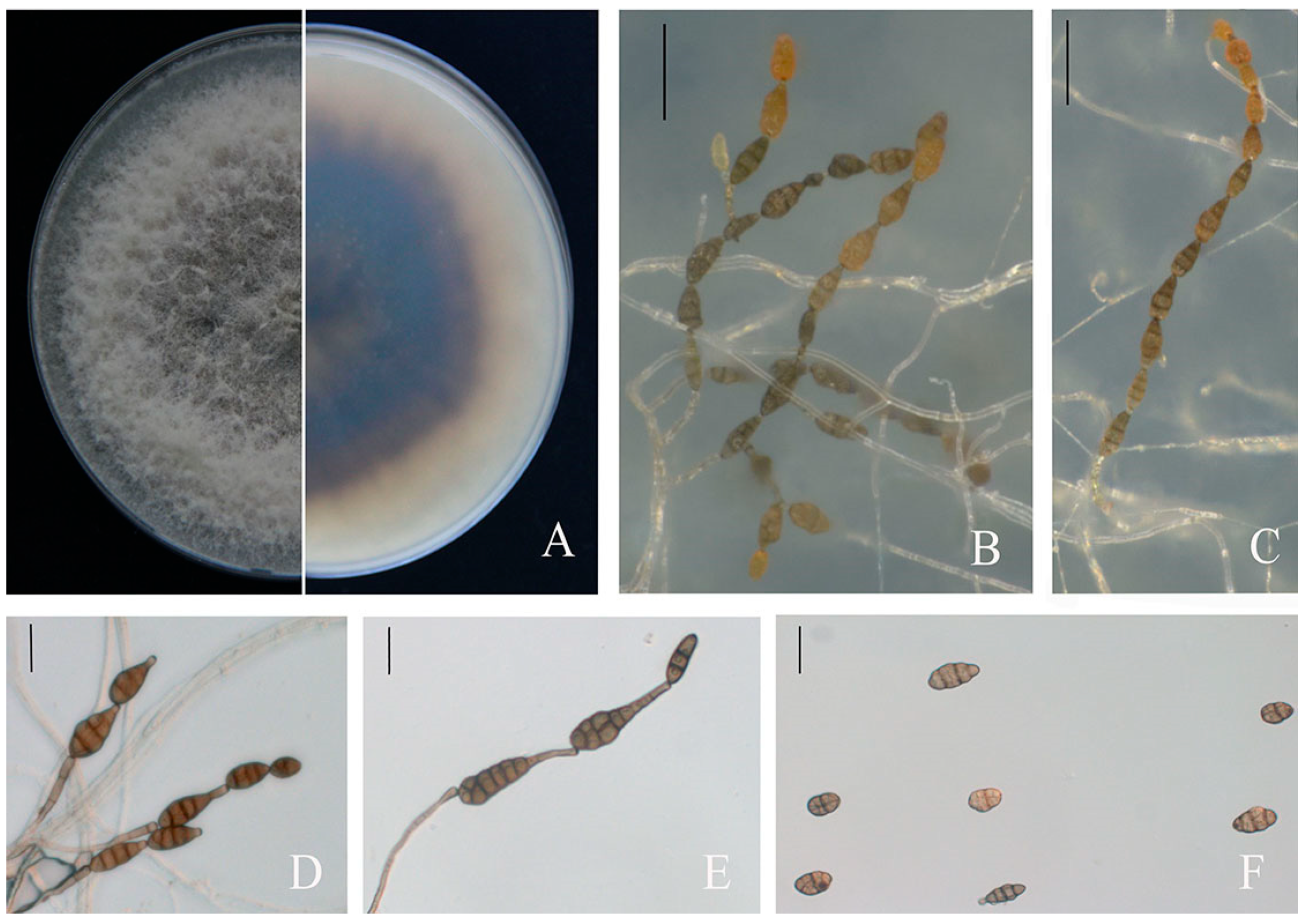
3.3. Morphology of Fungal Isolates
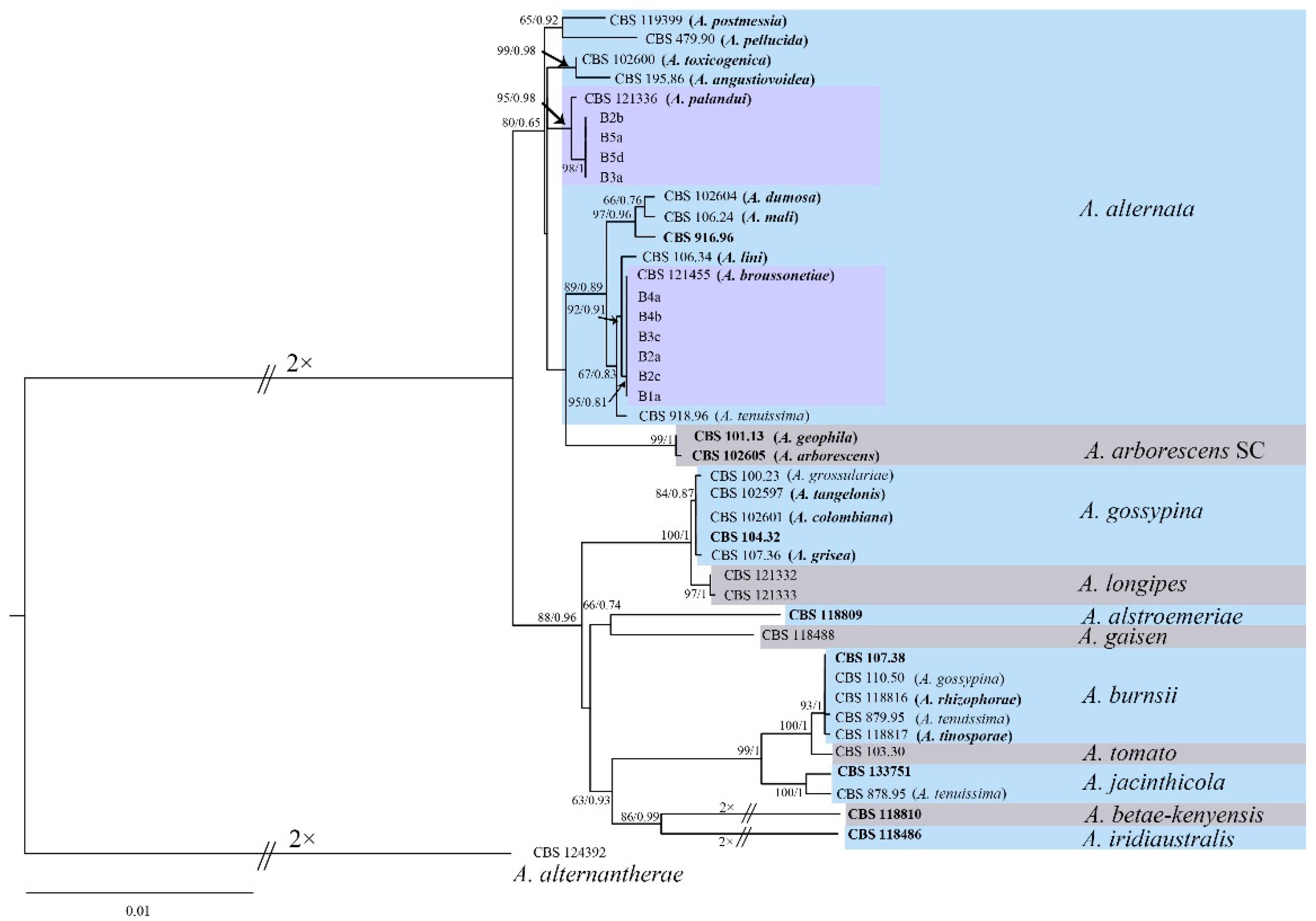
3.4. Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, Y.; Zhang, X.Z.; Wang, X.A. A study on geographical distribution law of Pinus bungeana natural forests in china. Acta Bot. Boreali-Occident. Sin. 1995, 15, 161–166. [Google Scholar]
- Guo, C.C.; Shen, Y.B.; Shi, F.H. Investigating seed dormancy in Pinus bungeana Zucc. ex Endl.: Understanding the contributions of enclosing tissues and temperature on germination. Forests 2018, 9, 401. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, J.; Ji, Z.; Xu, F. Comparison of anatomy and composition distribution between normal and compression wood of Pinus bungeana Zucc. revealed by microscopic imaging techniques. Microsc. Microanal. 2012, 18, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.; Jia, Y.; Yang, J.; Li, Z.H. Comparative transcriptome analysis of male and female conelets and development of microsatellite markers in Pinus bungeana, an endemic conifer in China. Genes 2017, 8, 393. [Google Scholar] [CrossRef]
- Yang, Y.X.; Wang, M.L.; Liu, Z.L.; Zhu, J.; Yan, M.Y.; Li, Z.H. Nucleotide polymorphism and phylogeographic history of an endangered conifer species Pinus bungeana. Biochem. Syst. Ecol. 2016, 64, 89–96. [Google Scholar] [CrossRef]
- Li, Z.H.; Zhu, J.; Yang, Y.X.; Yang, J.; He, J.W.; Zhao, G.F. The complete plastid genome of Bunge’s pine Pinus bungeana (Pinaceae). Mitochondr. DNA 2016, 27, 2971–2972. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The sections of Alternaria: Formalizing species-group concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria. An Identification Manual. CBS Biodiversity Series 6; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Xu, B.; Song, J.; Xi, P.; Li, M.; Hsiang, T.; Jiang, Z. A destructive leaf spot and blight caused by Alternaria kareliniae sp. nov. on a sand-stabilizing plant, caspian sea karelinia. Plant Dis. 2018, 102, 172–178. [Google Scholar] [CrossRef]
- Matic, S.; Tabone, G.; Garibaldi, A.; Gullino, M.L. Alternaria leaf spot caused by Alternaria species: An emerging problem on ornamental plants in Italy. Plant Dis. 2020, 104, 2275–2287. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and taxonomy of the pleomorphic genus Alternaria. Mycol. Prog. 2016, 15, 3. [Google Scholar] [CrossRef]
- Thomma, B. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Kahl, S.M.; Ulrich, A.; Kirichenko, A.A.; Muller, M.E.H. Phenotypic and phylogenetic segregation of Alternaria infectoria from small-spored Alternaria species isolated from wheat in Germany and Russia. J. Appl. Microbiol. 2015, 119, 1637–1650. [Google Scholar] [CrossRef] [PubMed]
- Al-Nadabi, H.H.; Maharachchikumbura, S.S.N.; Agrama, H.; Al-Azri, M.; Nasehi, A.; Al-Sadi, A.M. Molecular characterization and pathogenicity of Alternaria species on wheat and date palms in Oman. Eur. J. Plant Pathol. 2018, 152, 577–588. [Google Scholar] [CrossRef]
- Elfar, K.; Zoffoli, J.P.; Latorre, B.A. Identification and characterization of Alternaria species associated with moldy core of apple in Chile. Plant Dis. 2018, 102, 2158–2169. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, M.; Venkataravanappa, V.; Saha, S.; Rai, A.B.; Tripathi, S.; Rai, R.K.; Pandey, A.K. Morphological, pathogenic and molecular characterizations of Alternaria species causing early blight of tomato in Northern India. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 86, 325–330. [Google Scholar] [CrossRef]
- Zhu, Y.; Lujan, P.; Dura, S.; Steiner, R.; Zhang, J.; Sanogo, S. Etiology of Alternaria leaf spot of cotton in Southern New Mexico. Plant Dis. 2019, 103, 1595–1604. [Google Scholar] [CrossRef]
- Andrew, M.; Peever, T.L.; Pryor, B.M. An expanded multilocus phylogeny does not resolve morphological species within the small-spored Alternaria species complex. Mycologia 2009, 101, 95–109. [Google Scholar] [CrossRef]
- Pryor, B.M.; Bigelow, D.M. Molecular characterization of Embellisia and Nimbya species and their relationship to Alternaria, Ulocladium and Stemphylium. Mycologia 2003, 95, 1141–1154. [Google Scholar] [CrossRef]
- Woudenberg, J.H.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef]
- Luo, Y.; Hou, L.; Forster, H.; Pryor, B.; Adaskaveg, J.E. Identification of Alternaria species causing heart rot of pomegranates in California. Plant Dis. 2017, 101, 421–427. [Google Scholar] [CrossRef]
- Zhu, X.Q.; Xiao, C.L. Phylogenetic, morphological and pathogenic characterization of Alternaria species Associated with fruit rot of blueberry in California. Phytopathology 2015, 105, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Pryor, B.M.; Gilbertson, R.L. Molecular phylogenetic relationships amongst Alternaria species and related fungi based upon analysis of nuclear ITS and mt SSU rDNA sequences. Mycol. Res. 2000, 104, 1312–1321. [Google Scholar] [CrossRef]
- Rang, J.C.; Crous, P.W.; Mchau, G.R.A.; Serdani, M.; Song, S.M. Phylogenetic analysis of Alternaria spp. associated with apple core rot and citrus black rot in South Africa. Mycol. Res. 2002, 106, 1151–1162. [Google Scholar] [CrossRef]
- Sun, X.M.; Huang, J.G. First report of Alternaria tenuissima causing red leaf spot disease on Paeonia lactiflora in China. Plant Dis. 2017, 101, 1322. [Google Scholar] [CrossRef]
- Wang, Q.; Luan, Q.S.; Wang, J.J.; Jiang, X.; Chen, G. First report of Alternaria alternata causing leaf blight on Actinidia arguta in Liaoning, China. Plant Dis. 2020, 104, 2725–2726. [Google Scholar] [CrossRef]
- Mmbaga, M.T.; Shi, A.N.; Kim, M.S. Identification of Alternaria alternata as a causal agent for leaf blight in Syringa species. Plant Pathol. J. 2011, 27, 120–127. [Google Scholar] [CrossRef]
- Kim, H.Y.; Shin, J.H.; Lee, N.H.; Kim, S.; Kim, K. Isolation and evaluation of fungicides for control of Alternaria alternata causing Alternaria leaf spot disease on lettuce. J. Agric. Life Sci. 2021, 55, 89–97. [Google Scholar] [CrossRef]
- Zhang, J.X.; Kang, Y.; Xu, T.T.; Xu, X.Y.; Li, C.X.; Chen, K.F.; Cao, H.Q. First report of leaf spot caused by Alternaria alternata on Sonchus asper in China. Plant Dis. 2021, 105, 503. [Google Scholar] [CrossRef]
- Sun, H.F.; Li, N.; Yan, Y.; Wei, M.Y.; Wang, H.; Yang, H.Y. First report of Alternaria alternata causing leaf blight on Clematis terniflora var. mandshurica in China. Plant Dis. 2022, 106, 2264. [Google Scholar] [CrossRef]
- Long, H.J.; Yang, S.; Yin, X.H.; Zhao, Z.B.; Long, Y.H.; Fan, J.; Shu, R.; Gu, G.F. First report of leaf spot on Prunus salicina caused by Alternaria alternata in China. J. Plant Pathol. 2021, 103, 1021. [Google Scholar] [CrossRef]
- Qi, M.; Xie, C.X.; Chen, Q.W.; Yu, Z.D. Pestalotiopsis trachicarpicola, a novel pathogen causes twig blight of Pinus bungeana (Pinaceae: Pinoideae) in China. Antonie Van Leeuwenhoek. Int. J. Gen. Mol. Microbiol. 2021, 114, 1–9. [Google Scholar] [CrossRef]
- Lin, Y.R.; Tang, Y.P. Seven species of Lophodermium on pinus. Acta Mycol. Sin. 1988, 7, 129–137. [Google Scholar]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.S.; Waller, J.M.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Pryor, B.M.; Michailides, T.J. Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with Alternaria late blight of pistachio. Phytopathology 2002, 92, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H.I. Untersuchungen uber die morphologische und biologische differenzierung in der Fusaium-Sektion Liseola. In Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft Berlin-Dahlem; Kommissionsverlag Paul Parey: Berlin, Germany, 1976; Volume 169, pp. 1–117. [Google Scholar]
- Damm, U.; Mostert, L.; Crous, P.W.; Fourie, P.H. Novel Phaeoacremonium species associated with necrotic wood of Prunus trees. Persoonia 2008, 20, 87–102. [Google Scholar] [CrossRef] [PubMed]
- de Hoog, G.S.; Gerrits van den Ende, A.H. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.B.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef]
- Crous, P.W.; Schoch, C.L.; Hyde, K.D.; Wood, A.R.; Gueidan, C.; Hoog, G.D.; Groenewald, J.Z. Phylogenetic lineages in the Capnodiales. Stud. Mycol. 2009, 64, 17–47. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.R.; Zhang, M.J.; Shang, X.L.; Fang, S.Z.; Chen, F.M. Stem canker on Cyclocarya paliurus is caused by Botryosphaeria dothidea. Plant Dis. 2020, 104, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Moumni, M.; Allagui, M.B.; Mancini, V.; Murolo, S.; Tarchoun, N.; Romanazzi, G. Morphological and molecular identification of seedborne fungi in Squash (Cucurbita maxima, Cucurbita moschata). Plant Dis. 2020, 104, 1335–1350. [Google Scholar] [CrossRef]
- Gao, J.; Yang, M.J.; Xie, Z.; Lu, B.H.; Hsiang, T.; Liu, L.P. Morphological and molecular identification and pathogenicity of Alternaria spp. associated with ginseng in Jilin province, China. Can. J. Plant Pathol. 2021, 43, 537–550. [Google Scholar] [CrossRef]
- Woudenberg, J.H.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Stewart, J.E.; Timmer, L.W.; Lawrence, C.B.; Pryor, B.M.; Peever, T.L. Discord between morphological and phylogenetic species boundaries: Incomplete lineage sorting and recombination results in fuzzy species boundaries in an asexual fungal pathogen. BMC Evol. Biol. 2014, 14, 38. [Google Scholar] [CrossRef]
- Al-Lami, H.F.D.; You, M.P.; Barbetti, M.J. Incidence, pathogenicity and diversity of Alternaria spp. associated with alternaria leaf spot of canola (Brassica napus) in Australia. Plant Pathol. 2019, 68, 492–503. [Google Scholar] [CrossRef]
- Somma, S.; Amatulli, M.T.; Masiello, M.; Moretti, A.; Logrieco, A.F. Alternaria species associated to wheat black point identified through a multilocus sequence approach. Int. J. Food Microbiol. 2019, 293, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Kant, R.; Joshi, P.; Bhandari, M.S.; Pandey, A.; Pandey, S. Identification and pathogenicity of Alternaria alternata causing leaf spot and blight disease of Ailanthus excelsa in India. Forest Pathol. 2020, 50, e12584. [Google Scholar] [CrossRef]
- Tian, Y.; Qiu, C.D.; Zhang, Y.Y.; Liu, Z.Y. First report of Alternaria alternata causing leaf spot on Chaenomeles cathayensis in Anhui province of China. Plant Dis. 2020, 104, 279–280. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: An overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- O’Donnell, J.; Dickinson, C.H. Pathogenicity of Alternaria and Cladosporium isolates on Phaseolus. Trans. Br. Mycol. Soc. 1980, 74, 335–342. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, L.D. A comparative study of endophytic fungi in needles, bark, and xylem of Pinus tabulaeformis. Can. J. Bot. 2007, 85, 911–917. [Google Scholar] [CrossRef]
- Nguyen, M.H.; Yong, J.H.; Sung, H.J.; Lee, J.K. Screening of endophytic fungal isolates dgainst Raffaelea quercus-mongolicae causing oak wilt disease in Korea. Mycobiology 2020, 48, 484–494. [Google Scholar] [CrossRef]
- Behnke-Borowczyk, J.; Kwaśna, H.; Kulawinek, B. Fungi associated with Cyclaneusma needle cast in Scots pine in the west of Poland. Forest Pathol. 2019, 49, e12487. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primer | Sequence (5′-3′) | PCR Conditions | Reference |
|---|---|---|---|---|
| Internal transcribed spacer (ITS) | V9G | TTACGTCCCTGCCCTTTGTA | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 48 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [38] |
| ITS4 | TCCTCCGCTTATTGATATGC | [39] | ||
| Elongation factor 1-alpha (tef1) | EF1-728F | CATCGAGAAGTTCGAGAAGG | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 55 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [40] |
| EF1-986R | TACTTGAAGGAACCCTTACC | |||
| Endopolygalacturonase (endoPG) | PG3 | TACCATGGTTCTTTCCGA | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 50 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [18] |
| PG2b | GAGAATTCRCARTCRTCYTGRTT | |||
| Anonymous gene region (OPA 10-2) | OPA 10-2R | GATTCGCAGCAGGGAAACTA | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 58 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [18] |
| OPA 10-2L | TCGCAGTAAGACACA TTCTACG | |||
| Alternaria major allergen gene (Alt a 1) | Alt-for | ATGCAGTTCACCACCATCGC | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 60 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [41] |
| Alt-rev | ACGAGGGTGAY GTAGGCGTC | |||
| 18S nrDNA (SSU) | NS1 | GTAGTCATATGCTTGTCTC | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 55 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [39] |
| NS4 | CTTCCGTCAATTCCTTTAAG | |||
| 28S nrDNA (LSU) | LSU1Fd | GRATCAGGTAGG RATACCCG | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 51 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [42] |
| LR5 | ATCCTGAGGGAAACTTC | [43] | ||
| glyceraldehyde-3-phosphate dehydrogenase (gapdh) | gpd1 | CAACGGCTTCGGTCG CATTG | Denaturation for 3 min at 94 °C, followed by 30 cycles; 30 s at 94 °C, 30 s at 57 °C, 30 s at 72 °C, and 10 min of a final extension at 72 °C | [44] |
| gpd2 | GCCAAGCAGTTGGTTGTGC |
| Species Name and Strain Number 1,2 | Locality, Host/Substrate | GenBank Accession Numbers 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| SSU | LSU | ITS | gapdh | tef1 | Alt a 1 | endoPG | OPA10-2 | ||
| Alternaria alstroemeriae | |||||||||
| CBS 118809 T | Australia, Alstroemeria sp. | KP124918 | KP124448 | KP124297 | KP124154 | KP125072 | np | KP123994 | KP124602 |
| Alternaria alternantherae | |||||||||
| CBS 124392 | China, Solanum melongena | KC584506 | KC584251 | KC584179 | KC584096 | KC584633 | KP123846 | np | np |
| Alternaria alternata | |||||||||
| B4b | China, Pinus bungeana | MZ835355 | MZ835345 | MZ823461 | MZ835385 | MZ835395 | MZ802959 | MZ835375 | MZ835365 |
| B2c | China, Pinus bungeana | MZ835356 | MZ835346 | MZ823462 | MZ835386 | MZ835396 | MZ802960 | MZ835376 | MZ835366 |
| B4a | China, Pinus bungeana | MZ835357 | MZ835347 | MZ823463 | MZ835387 | MZ835397 | MZ802961 | MZ835377 | MZ835367 |
| B1a | China, Pinus bungeana | MZ835358 | MZ835348 | MZ823464 | MZ835388 | MZ835398 | MZ802962 | MZ835378 | MZ835368 |
| B2a | China, Pinus bungeana | MZ835359 | MZ835349 | MZ823465 | MZ835389 | MZ835399 | MZ802963 | MZ835379 | MZ835369 |
| B3c | China, Pinus bungeana | MZ835360 | MZ835350 | MZ823466 | MZ835390 | MZ835400 | MZ802964 | MZ835380 | MZ835370 |
| B2b | China, Pinus bungeana | MZ835361 | MZ835351 | MZ823467 | MZ835391 | MZ835401 | MZ802965 | MZ835381 | MZ835371 |
| B5d | China, Pinus bungeana | MZ835362 | MZ835352 | MZ823468 | MZ835392 | MZ835402 | MZ802966 | MZ835382 | MZ835372 |
| B5a | China, Pinus bungeana | MZ835363 | MZ835353 | MZ823469 | MZ835393 | MZ835403 | MZ802967 | MZ835383 | MZ835373 |
| B3a | China, Pinus bungeana | MZ835364 | MZ835354 | MZ823470 | MZ835394 | MZ835404 | MZ802968 | MZ835384 | MZ835374 |
| CBS 916.96 T | India, Arachis hypogaea | KC584507 | DQ678082 | AF347031 | AY278808 | KC584634 | AY563301 | JQ811978 | KP124632 |
| CBS 195.86 (A. angustiovoidea T) | Canada, Euphorbia esula | KP124939 | KP124469 | KP124317 | KP124173 | KP125093 | JQ646398 | KP124017 | KP124624 |
| CBS 106.24 (A. mali T) | USA, Malus sylvestris | KP124919 | KP124449 | KP124298 | KP124155 | KP125073 | KP123847 | AY295020 | JQ800620 |
| CBS 102604 (A. Dumosa T) | Israel, Minneola tangelo | KP124956 | KP124486 | KP124334 | AY562410 | KP125110 | AY563305 | KP124035 | KP124643 |
| CBS 106.34 (A. lini T) | Unknown, Linum usitatissimum | KP124924 | KP124454 | Y17071 | JQ646308 | KP125078 | KP123853 | KP124000 | KP124608 |
| CBS 918.96 (A. tenuissima R) | UK, Dianthus chinensis | KC584567 | KC584311 | AF347032 | AY278809 | KC584693 | AY563302 | KP124026 | KP124633 |
| CBS 479.90 (A. pellucida T) | Japan, Citrus unshiu | KP124941 | KP124471 | KP124319 | KP124174 | KP125095 | KP123870 | KP124019 | KP124626 |
| CBS 102600 (A. toxicogenica T) | USA, Citrus reticulata | KP124953 | KP124483 | KP124331 | KP124186 | KP125107 | KP123880 | KP124033 | KP124640 |
| CBS 119399 (A. postmessia T) | USA, Minneola tangelo | KP124983 | KP124513 | KP124361 | JQ646328 | KP125137 | KP123910 | KP124063 | KP124672 |
| CBS 121336 (A. palandui T) | USA, Allium sp. | KP124987 | KP124517 | KJ862254 | KJ862255 | KP125141 | KJ862259 | KP124067 | KP124676 |
| CBS 121455 (A. broussonetiae T) | China, Broussonetia papyrifera | KP124992 | KP124522 | KP124368 | KP124220 | KP125146 | KP123916 | KP124072 | KP124681 |
| Alternaria arborescens species complex (AASC) | |||||||||
| CBS 101.13 (A. geophila T) | Switzerland, peat soil | KP125016 | KP124546 | KP124392 | KP124244 | KP125170 | KP123940 | KP124096 | KP124705 |
| CBS 102605 (A. arborescens T) | USA, Solanum lycopersicum | KC584509 | KC584253 | AF347033 | AY278810 | KC584636 | AY563303 | AY295028 | KP124712 |
| Alternaria betae-kenyensis | |||||||||
| CBS 118810 T | Kenya, Beta vulgaris var. cicla | KP125042 | KP124572 | KP124419 | KP124270 | KP125197 | KP123966 | KP124123 | KP124733 |
| Alternaria burnsii | |||||||||
| CBS 107.38 T | India, Cuminum cyminum | KP125043 | KP124573 | KP124420 | JQ646305 | KP125198 | KP123967 | KP124124 | KP124734 |
| CBS 110.50 (A. gossypina) | Mozambique, Gossypium sp. | KP125044 | KP124574 | KP124421 | KP124271 | KP125199 | KP123968 | KP124125 | KP124735 |
| CBS 879.95 (A. tenuissima) | UK, Sorghum sp. | KP125045 | KP124575 | KP124422 | KP124272 | KP125200 | KP123969 | KP124126 | KP124736 |
| CBS 118816 (A. rhizophorae T) | India, Rhizophora mucronata | KP125046 | KP124576 | KP124423 | KP124273 | KP125201 | KP123970 | KP124127 | KP124737 |
| CBS 118817 (A. tinosporae T) | India, Tinospora cordifolia | KP125047 | KP124577 | KP124424 | KP124274 | KP125202 | KP123971 | KP124128 | KP124738 |
| Alternaria gaisen | |||||||||
| CBS 118488 R | Japan, Pyrus pyrifolia | KP125051 | KP124581 | KP124427 | KP124278 | KP125206 | KP123975 | KP124132 | KP124743 |
| Alternaria gossypina | |||||||||
| CBS 100.23 (A. grossulariae) | Unknown, Malus domestica | KP125053 | KP124583 | KP124429 | KP124280 | KP125208 | KP123977 | KP124134 | KP124745 |
| CBS 104.32 T | Zimbabwe, Gossypium sp. | KP125054 | KP124584 | KP124430 | JQ646312 | KP125209 | JQ646395 | KP124135 | KP124746 |
| CBS 107.36 (A. grisea T) | Indonesia, soil | KP125055 | KP124585 | KP124431 | JQ646310 | KP125210 | JQ646393 | KP124136 | KP124747 |
| CBS 102597 (A. tangelonis T) | USA, Minneola tangelo | KP125056 | KP124586 | KP124432 | KP124281 | KP125211 | KP123978 | KP124137 | KP124748 |
| CBS 102601 (A. colombiana T) | Colombia, Minneola tangelo | KP125057 | KP124587 | KP124433 | KP124282 | KP125212 | KP123979 | KP124138 | KP124749 |
| Alternaria iridiaustralis | |||||||||
| CBS 118486 T | Australia, Iris sp. | KP125059 | KP124589 | KP124435 | KP124284 | KP125214 | KP123981 | KP124140 | KP124751 |
| Alternaria jacinthicola | |||||||||
| CBS 878.95 (A. tenuissima) | Mauritius, Arachis hypogaea | KP125061 | KP124591 | KP124437 | KP124286 | KP125216 | KP123983 | KP124142 | KP124753 |
| CBS 133751 T | Mali, Eichhornia crassipes | KP125062 | KP124592 | KP124438 | KP124287 | KP125217 | KP123984 | KP124143 | KP124754 |
| Alternaria longipes | |||||||||
| CBS 121333 R | USA, Nicotiana tabacum | KP125068 | KP124598 | KP124444 | KP124293 | KP125223 | KP123990 | KP124150 | KP124761 |
| CBS 12133 | USA, Nicotiana tabacum | KP125067 | KP124597 | KP124443 | KP124292 | KP125222 | KP123989 | KP124149 | KP124760 |
| Alternaria tomato | |||||||||
| CBS 103.30 | Unknown, Solanum lycopersicum | KP125069 | KP124599 | KP124445 | KP124294 | KP125224 | KP123991 | KP124151 | KP124762 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.-J.; Zheng, X.-R.; Li, H.; Chen, F.-M. Alternaria alternata, the Causal Agent of a New Needle Blight Disease on Pinus bungeana. J. Fungi 2023, 9, 71. https://doi.org/10.3390/jof9010071
Zhang M-J, Zheng X-R, Li H, Chen F-M. Alternaria alternata, the Causal Agent of a New Needle Blight Disease on Pinus bungeana. Journal of Fungi. 2023; 9(1):71. https://doi.org/10.3390/jof9010071
Chicago/Turabian StyleZhang, Mao-Jiao, Xiang-Rong Zheng, Huan Li, and Feng-Mao Chen. 2023. "Alternaria alternata, the Causal Agent of a New Needle Blight Disease on Pinus bungeana" Journal of Fungi 9, no. 1: 71. https://doi.org/10.3390/jof9010071
APA StyleZhang, M.-J., Zheng, X.-R., Li, H., & Chen, F.-M. (2023). Alternaria alternata, the Causal Agent of a New Needle Blight Disease on Pinus bungeana. Journal of Fungi, 9(1), 71. https://doi.org/10.3390/jof9010071