
Biodiversity and Bioprospecting of Fungal Endophytes from the Antarctic Plant Colobanthus quitensis


 , ,
, ,  , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Isolation of Culturable Fungal Endophytes
2.3. Identification and Selection of Fungal Endophytes
2.4. Fungal Growth and Samples Preparation
2.5. Antibacterial Activity Evaluation
2.6. Extracellular Enzyme Activities
2.7. Fungal Metabolite Extraction
2.8. Determination of Total Phenolic and Total Flavonoid Content
2.9. Cell Viability Assay
2.10. Expression and Purification of Recombinant Human α-Synuclein
2.11. Aggregation of α-Synuclein In Vitro
2.12. Plant Growth Conditions and Plant-Fungi Co-Cultivation Experiments
2.13. Co-Cultures of Antarctic Fungi and Fungal Phytopathogens
2.14. Statistical Analysis
3. Results and Discussion
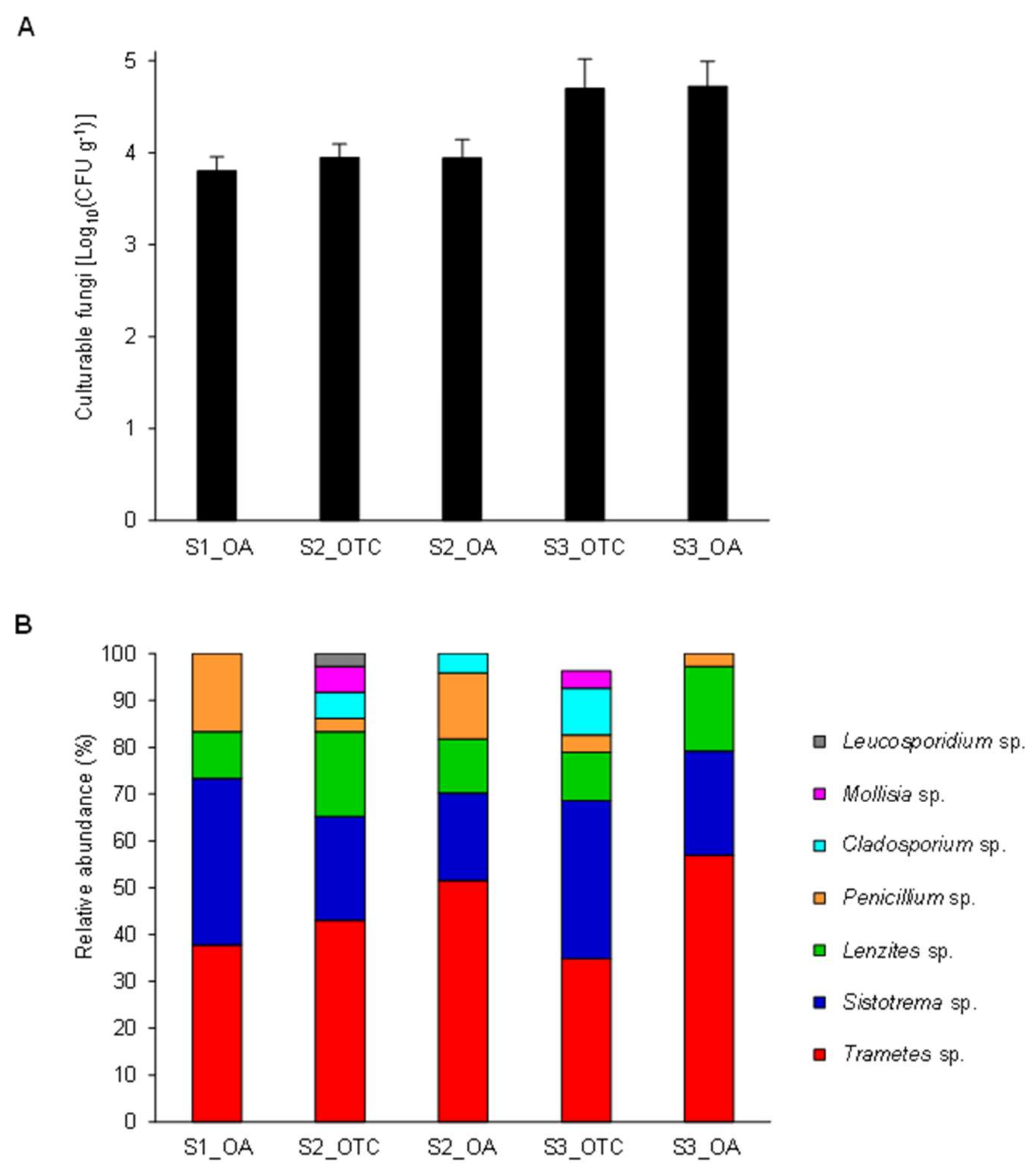
3.1. Isolation and Identification of Endophytic Fungi from the Antarctic Plant Colobanthus quitensis
3.2. Antimicrobial and Enzymatic Activity of Selected Newly Isolated Endophytic Fungi
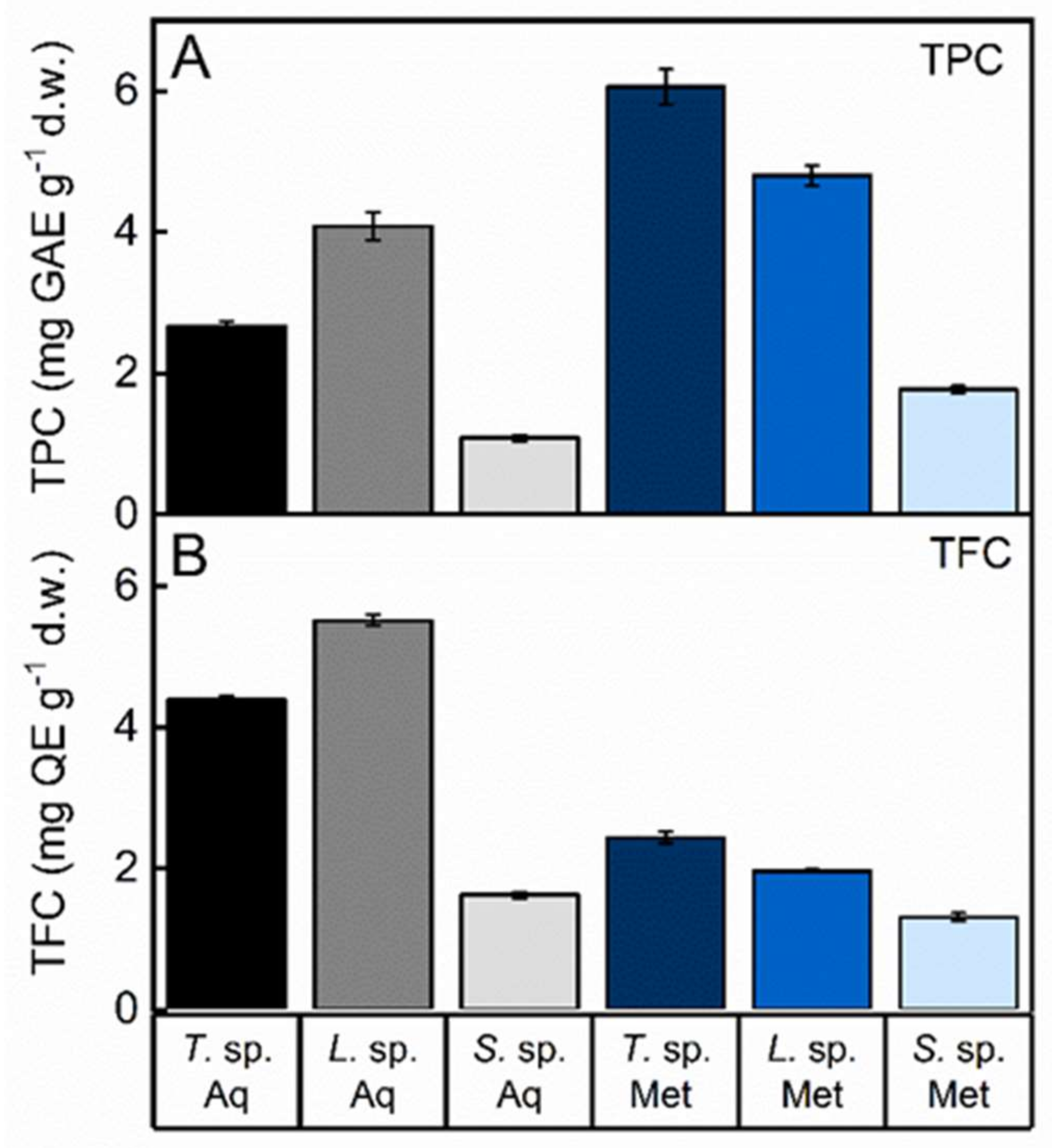
3.3. Total Phenolic and Flavonoid Content of Fungal Extracts
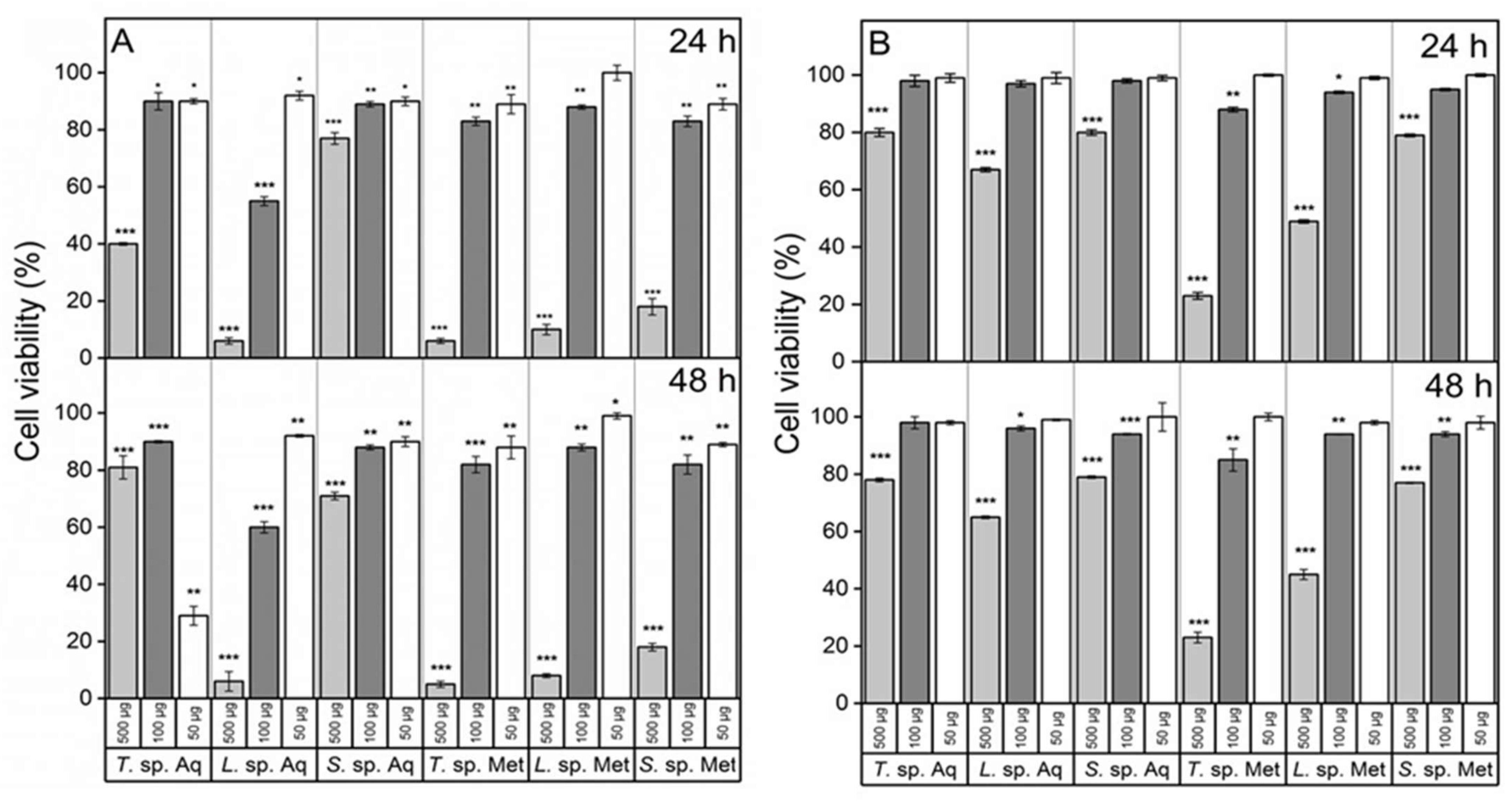
3.4. Cancer Cells Appear to Be Sensitive to Fungal Extracts
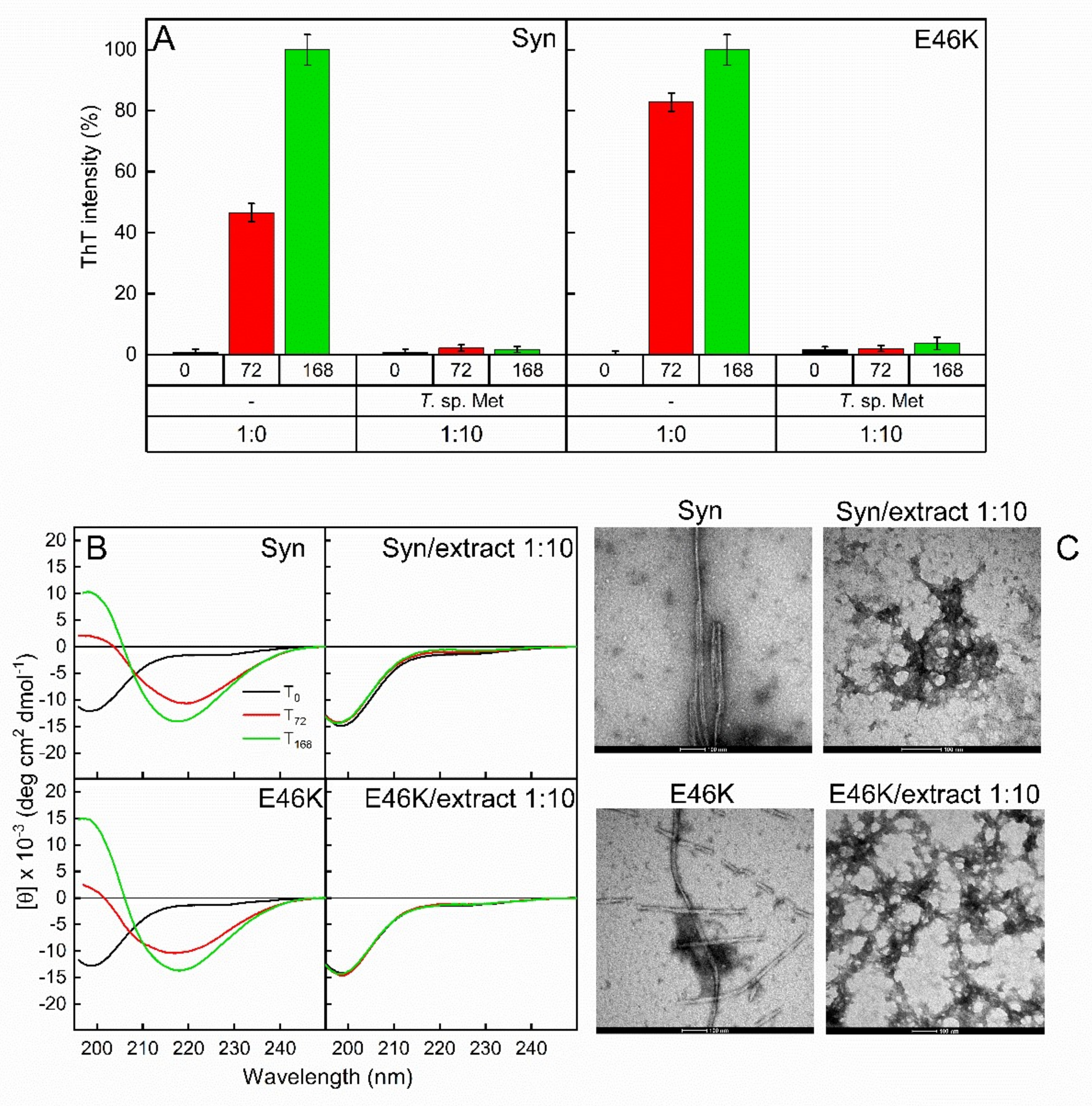
3.5. Fungal Extracts Affect the Formation of Amyloid Fibrils
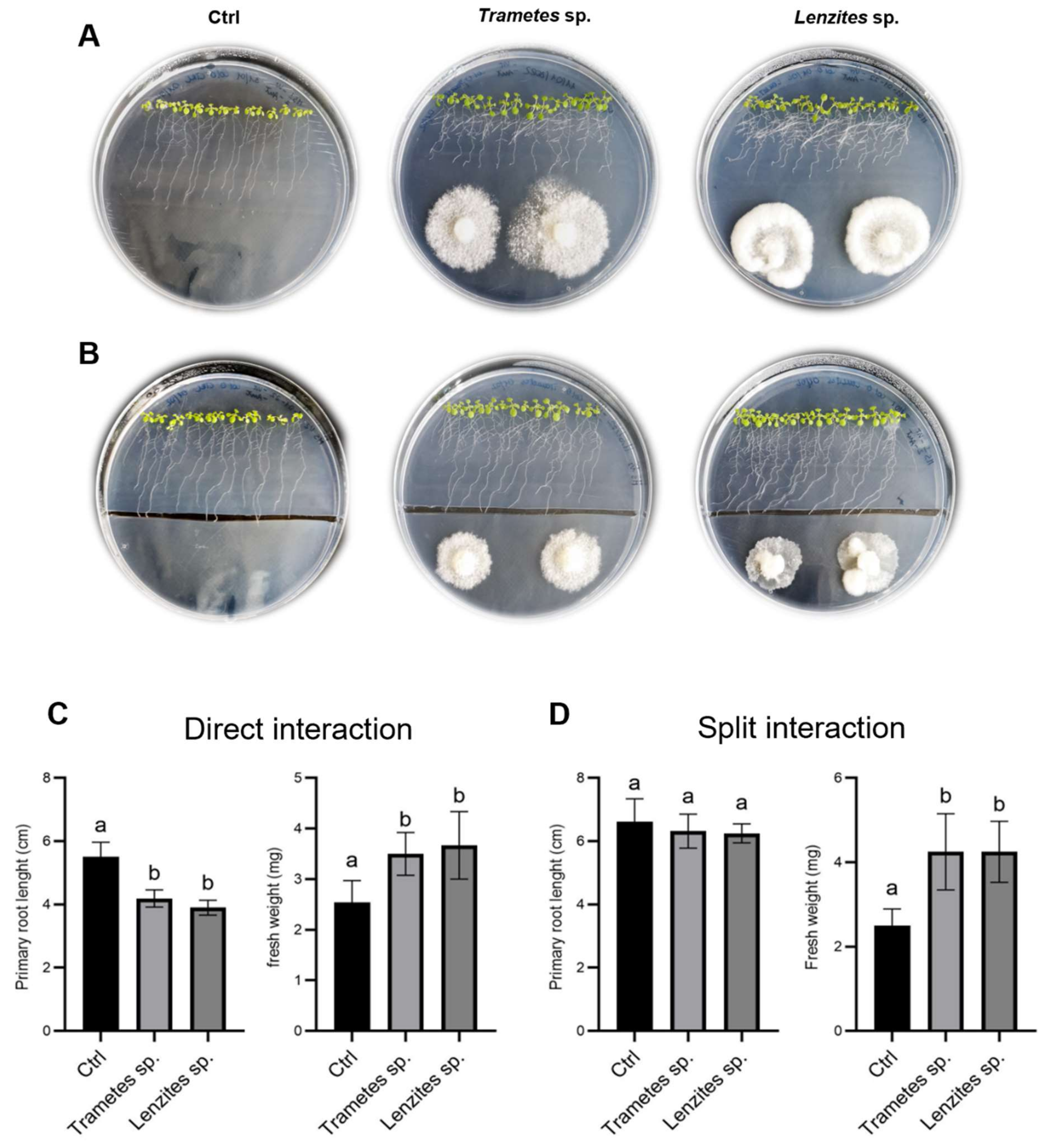
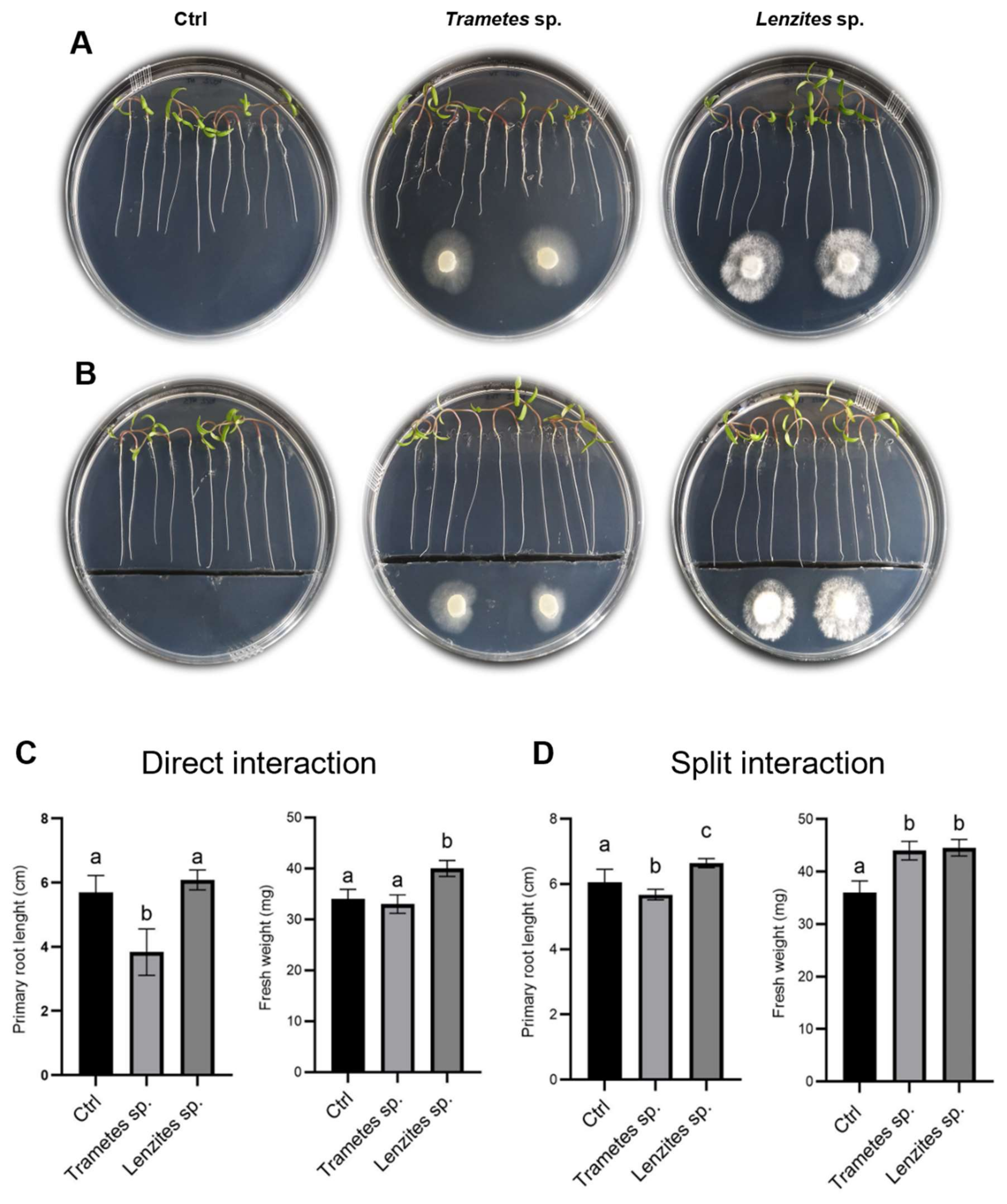
3.6. Antarctic Fungi Affect the Root Architecture in Arabidopsis and Tomato
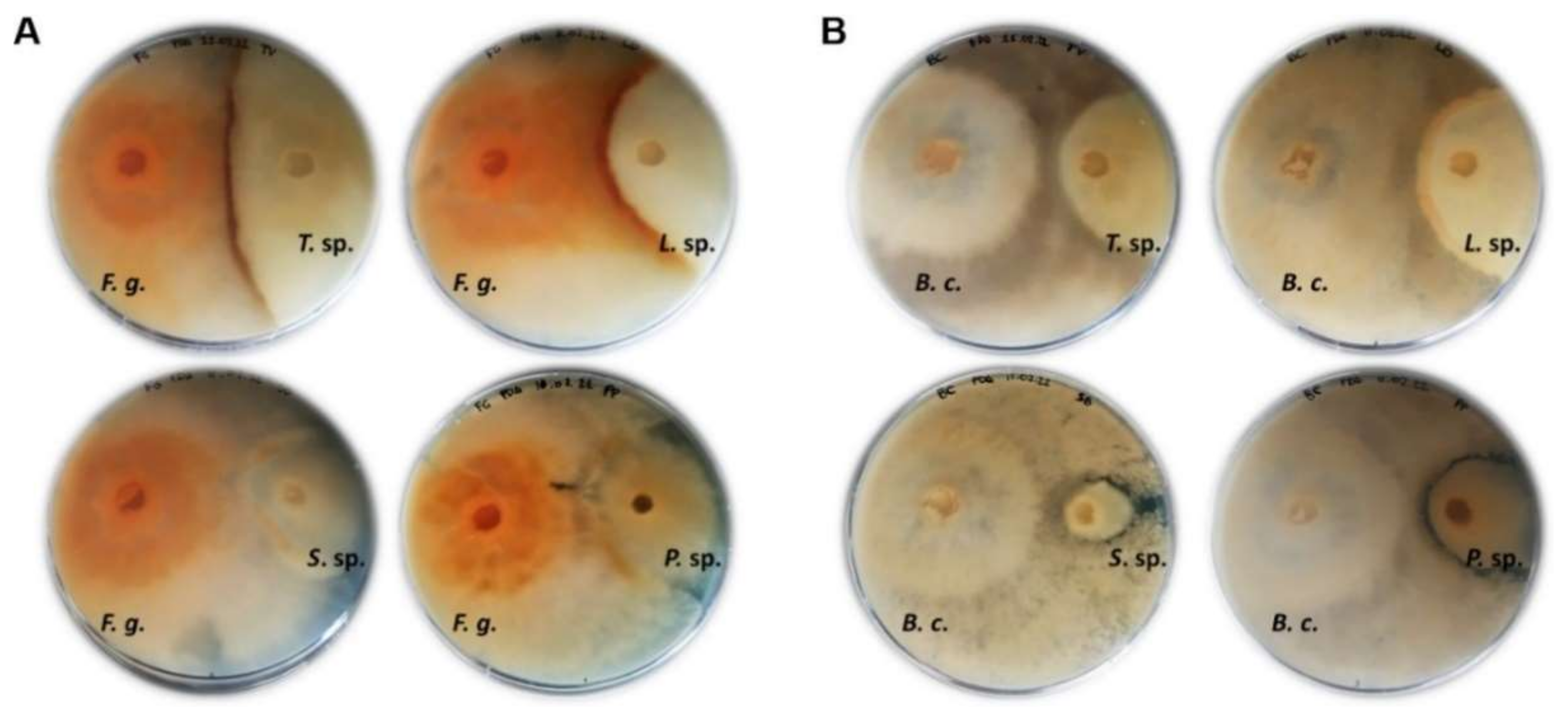
3.7. Antarctic Fungi Antagonize the Growth of Crop Plant Pathogenic Fungi
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Danilovich, M.E.; Sánchez, L.A.; Acosta, F.F.; Delgado, O.D. Antarctic bioprospecting: In pursuit of microorganisms producing new antimicrobials and enzymes. Polar Biol. 2018, 41, 1417–1433. [Google Scholar] [CrossRef]
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biotechnol. 2015, 99, 7907–7913. [Google Scholar] [CrossRef] [PubMed]
- Timling, I.; Taylor, D.L. Peeking through a frosty window: Molecular insights into the ecology of Arctic soil fungi. Fungal Ecol. 2012, 5, 419–429. [Google Scholar] [CrossRef]
- Weinstein, R.N.; Montiel, P.O.; Johnstone, K. Influence of growth temperature on lipid and soluble carbohydrate synthesis by fungi isolated from fellfield soil in the maritime Antarctic. Mycologia 2000, 92, 222–229. [Google Scholar] [CrossRef]
- Coleine, C.; Stajich, J.E.; de los Ríos, A.; Selbmann, L. Beyond the extremes: Rocks as ultimate refuge for fungi in drylands. Mycologia 2021, 113, 108–133. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.H. Cold adaptation in Arctic and Antarctic fungi. New Phytol. 2001, 151, 341–353. [Google Scholar] [CrossRef]
- Zucconi, L.; Canini, F.; Temporiti, M.E.; Tosi, S. Extracellular enzymes and bioactive compounds from Antarctic terrestrial fungi for bioprospecting. Int. J. Environ. Res. Public Health 2020, 17, 6459. [Google Scholar] [CrossRef]
- Krüger, A.; Schäfers, C.; Schröder, C.; Antranikian, G. Towards a sustainable biobased industry—Highlighting the impact of extremophiles. New Biotechnol. 2018, 40, 144–153. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2017, 38, 600–619. [Google Scholar] [CrossRef]
- Loperena, L.; Soria, V.; Varela, H.; Lupo, S.; Bergalli, A.; Guigou, M.; Pellegrino, A.; Bernardo, A.; Calviño, A.; Rivas, F.; et al. Extracellular enzymes produced by microorganisms isolated from maritime Antarctica. World J. Microbiol. Biotechnol. 2012, 28, 2249–2256. [Google Scholar] [CrossRef]
- Gabani, P.; Singh, O.V. Radiation-resistant extremophiles and their potential in biotechnology and therapeutics. Appl. Microbiol. Biotechnol. 2013, 97, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Li, Y.-L.; Zhao, F.-C. Secondary Metabolites from Polar Organisms. Mar. Drugs 2017, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Purić, J.; Vieira, G.; Cavalca, L.B.; Sette, L.D.; Ferreira, H.; Vieira, M.L.C.; Sass, D.C. Activity of Antarctic fungi extracts against phytopathogenic bacteria. Lett. Appl. Microbiol. 2018, 66, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Santiago, I.F.; Alves, T.M.A.; Rabello, A.; Sales Junior, P.A.; Romanha, A.J.; Zani, C.L.; Rosa, C.A.; Rosa, L.H. Leishmanicidal and antitumoral activities of endophytic fungi associated with the Antarctic angiosperms Deschampsia antarctica Desv. and Colobanthus quitensis (Kunth) Bartl. Extremophiles 2012, 16, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Dagan, T.; Deines, P.; Dubilier, N.; Duschl, W.J.; Fraune, S.; Hentschele, U.; Hirtf, H.; Hültera, N.; Lachnitb, T.; et al. Metaorganisms in extreme environments: Do microbes play a role in organismal adaptation? Zoology 2018, 127, 1–19. [Google Scholar] [CrossRef]
- Longton, R.E. Vegetation Ecology and Classification in the Antarctic Zone. Can. J. Bot. 1979, 57, 2264–2278. [Google Scholar] [CrossRef]
- Smith, R.I.L. The enigma of Colobanthus quitensis and Deschampsia antarctica in Antarctica. In Antarctic Biology in a Global Context; Huiskes, A.H.L., Gieskes, W.W.C., Rozema, J., Schorno, R.M.L., van der Vies, S.M., Wolff, W.J., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 234–239. [Google Scholar]
- Arthofer, W.; Bertini, L.; Caruso, C.; Cicconardi, F.; Delph, L.F.; Fields, P.D.; Ikeda, M.; Minegishi, Y.; Proietti, S.; Rotthamer, H.; et al. Genomic Resources Notes accepted 1 February 2015–31 March 2015. Mol. Ecol. Resour. 2015, 15, 1014–1015. [Google Scholar]
- Nibert, M.L.; Manny, A.R.; Debat, H.J.; Firth, A.E.; Bertini, L.; Caruso, C. A barnavirus sequence mined from a transcriptome of the Antarctic pearlwort Colobanthus quitensis. Arch. Virol. 2018, 163, 1921–1926. [Google Scholar] [CrossRef]
- Ballesteros, G.I.; Torres-Díaz, C.; Bravo, L.A.; Balboa, K.; Caruso, C.; Bertini, L.; Proietti, S.; Molina-Montenegro, M.A. In silico analysis of metatranscriptomic data from the Antarctic vascular plant Colobanthus quitensis: Responses to a global warming scenario through changes in fungal gene expression levels. Fungal Ecol. 2020, 43, 100873. [Google Scholar] [CrossRef]
- Torres-Díaz, C.; Gallardo-Cerda, J.; Lavin, P.; Oses, R.; Carrasco-Urra, F.; Atala, C.; Acuña-Rodríguez, I.S.; Convey, P.; Molina-Montenegro, M.A. Biological interactions and simulated climate change modulates the ecophysiological performance of Colobanthus quitensis in the Antarctic ecosystem. PLoS ONE 2016, 11, 10. [Google Scholar] [CrossRef]
- Gallardo-Cerda, J.; Levihuan, J.; Lavín, P.; Oses, R.; Atala, C.; Torres-Díaz, C.; Cuba-Díaz, M.; Barrera, A.; Molina-Montenegro, M.A. Antarctic rhizobacteria improve salt tolerance and physiological performance of the Antarctic vascular plants. Polar Biol. 2018, 41, 1973–1982. [Google Scholar] [CrossRef]
- Ramos, P.; Rivas, N.; Pollmann, S.; Casati, P.; Molina-Montenegro, M.A. Hormonal and physiological changes driven by fungal endophytes increase Antarctic plant performance under UV-B radiation. Fungal Ecol. 2018, 34, 76–82. [Google Scholar] [CrossRef]
- Barrera, A.; Hereme, R.; Ruiz-Lara, S.; Larrondo, L.F.; Gundel, P.E.; Pollmann, S.; Molina-Montenegro, M.A.; Ramos, P. Fungal endophytes enhance the photoprotective mechanisms and photochemical efficiency in the Antarctic Colobanthus quitensis (Kunth) Bartl. exposed to UV-B radiation. Front. Ecol. Evol. 2020, 8, 122. [Google Scholar] [CrossRef]
- Hereme, R.; Morales-Navarro, S.; Ballesteros, G.; Barrera, A.; Ramos, P.; Gundel, P.E.; Molina-Montenegro, M.A. Fungal endophytes exert positive effects on Colobanthus quitensis under water stress but neutral under a projected climate change scenario in Antarctica. Front. Microbiol. 2020, 11, 264. [Google Scholar] [CrossRef] [PubMed]
- Oses-Pedraza, R.; Torres-Díaz, C.; Lavín, P.; Retamales-Molina, P.; Atala, C.; Gallardo-Cerda, J.; Acuña-Rodríguez, I.S.; Molina-Montenegro, M.A. Root endophytic Penicillium promotes growth of Antarctic vascular plants by enhancing nitrogen mineralization. Extremophiles 2020, 24, 721–732. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Oses, R.; Torres-Diaz, C.; Atala, C.; Zurita-Silva, A.; Ruiz-Lara, S. Root-endophytes improve the ecophysiological performance and production of an agricultural species under drought condition. AoB Plants 2016, 8, plw062. [Google Scholar] [CrossRef]
- Acuña-Rodríguez, I.S.; Hansen, H.; Gallardo-Cerda, J.; Atala, C.; Molina-Montenegro, M.A. Antarctic extremophiles: Biotechnological alternative to crop productivity in saline soils. Front. Bioeng. Biotechnol. 2019, 7, 22. [Google Scholar] [CrossRef]
- Yadav, A.N. Beneficial plant-microbe interactions for agricultural sustainability. J. Appl. Biol. Biotechnol. 2021, 9, 1–4. [Google Scholar] [CrossRef]
- Chávez, R.; Fierro, F.; García-Rico, R.O.; Vaca, I. Filamentous fungi from extreme environments as a promising source of novel bioactive secondary metabolites. Front. Microbiol. 2015, 6, 903. [Google Scholar] [CrossRef]
- Galambos, N.; Compant, S.; Moretto, M.; Sicher, C.; Puopolo, G.; Wäckers, F.; Sessitsch, A.; Pertot, I.; Perazzolli, M. Humic Acid Enhances the Growth of Tomato Promoted by Endophytic Bacterial Strains through the Activation of Hormone-, Growth-, and Transcription-Related Processes. Front. Plant Sci. 2020, 11, 82267. [Google Scholar] [CrossRef]
- Arrigoni, E.; Albanese, D.; Longa, C.M.; Angeli, D.; Donati, C.; Ioriatti, C.; Pertot, I.; Perazzolli, M. Tissue age, orchard location and disease management influence the composition of fungal and bacterial communities present on the bark of apple trees. Environ. Microbiol. 2020, 22, 2080–2093. [Google Scholar] [CrossRef] [PubMed]
- Elazegui, F.A.; Castilla, N.P.; Dalisay, T.U.; Mew, T.W. Causal agent of red stripe disease of rice. Plant Dis. 2004, 88, 1310–1317. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Orpin, J.B.; Mzungu, I.; Usman-Sani, H. Fungal Infestation of Garri Sold around Dutsinma Metropolis. J. Proteom. Bioinform. 2020, 13, 1–4. [Google Scholar]
- Rosa, L.H.; Almeida Vieira Mde, L.; Santiago, I.F.; Rosa, C.A. Endophytic fungi community associated with the dicotyledonous plant Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae) in Antarctica. FEMS Microb. Ecol. 2010, 73, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.B. Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Egham, UK, 1971; p. 608. [Google Scholar]
- Watanabe, T. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and Key to Species, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 426. [Google Scholar]
- Hofstetter, V.; Buyck, B.; Eyssartier, G.; Schnee, S.; Gindro, K. The unbearable lightness of sequenced-based identification. Fungal Divers. 2019, 96, 243–284. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Favaro, L.; Corich, V.; Giacomini, A.; Basaglia, M.; Casella, S. Grape marcs as unexplored source of new yeasts for future biotechnological applications. World J. Microbiol. Biotechnol. 2013, 29, 1551–1562. [Google Scholar]
- Favaro, L.; Jooste, T.; Basaglia, M.; Rose, S.H.; Saayman, M.; Görgens, J.F.; Casella, S.; van Zyl, W.H. Codon-optimized glucoamylase sGAI of Aspergillus awamori improves starch utilization in an industrial yeast. Appl. Microbiol. Biotechnol. 2012, 95, 957–968. [Google Scholar] [CrossRef]
- Lim, C.K.; Ahmad, R.; Marzuki, N.F.; Goh, Y.K.; Azizan, K.A.; Goh, Y.K.; Goh, K.J. Optimization of metabolite extraction protocols for untargeted metabolite profiling of mycoparasitic Scytalidium parasiticum using LC-TOF-MS. Sains Malays. 2018, 47, 3061–3068. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Knežević, P.; Šibul, F.; Kaišarević, S.; Karaman, M. Bioactive phenolic compounds of two medicinal mushroom species Trametes versicolor and Stereum subtomentosum as antioxidant and antiproliferative agents. Chem. Biodivers. 2020, 17, 12. [Google Scholar] [CrossRef]
- Singleton, V.L. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Way, M.L. A comparison of laboratory analysis methods for total phenolic content of cider. Beverages 2020, 6, 55. [Google Scholar] [CrossRef]
- Shirazi, O.U. Determination of total phenolic, flavonoid content and free radical scavenging activities of common herbs and spices. J. Pharmacogn. Phytochem. 2014, 3, 104–108. [Google Scholar]
- Siddiqui, N. Spectrophotometric determination of the total phenolic content, spectral and fluorescence study of the herbal Unani drug Gul-e-Zoofa (Nepeta bracteata Benth). J. Taibah Univ. Med. Sci. 2017, 12, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Yang, M.H.; Wen, H.J.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Ghosh, M.; Sinha, B.N.; Seijas, J.A.; Vázquez-Tato, M.P.; Feás, X. Flavonoids and phenolic compounds from Litsea polyantha Juss. bark. In Medicinal and Natural Products Chemistry, Proceedings of the 18th International Electronic Conference On Synthetic Organic Chemistry (ECSOC-18), Lugo, Spain, 1–30 November 2014; Seijas, J.A., Vázquez-Tato, M.P., Lin, S.K., Eds.; MDPI: Basel, Switzerland, 2015. [Google Scholar]
- De Franceschi, G.; Frare, E.; Bubacco, L.; Mammi, S.; Fontana, A.; Polverino de Laureto, P. Molecular insights into the interaction between alpha-synuclein and docosahexaenoic acid. J. Mol. Biol. 2009, 394, 94–107. [Google Scholar] [CrossRef] [PubMed]
- LeVine, H., 3rd. Thioflavine T interaction with synthetic Alzheimer’s disease beta-amyloid peptides: Detection of amyloid aggregation in solution. Protein Sci. 1993, 2, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Santiago, I.F.; Rosa, C.A.; Rosa, L.H. Endophytic symbiont yeasts associated with the Antarctic angiosperms Deschampsia antarctica and Colobanthus quitensis. Polar Biol. 2017, 40, 177–183. [Google Scholar] [CrossRef]
- Coelho, L.d.C.; de Carvalho, C.R.; Rosa, C.A.; Rosa, L.H. Diversity, distribution, and xerophilic tolerance of cultivable fungi associated with the Antarctic angiosperms. Polar Biol. 2021, 44, 379–388. [Google Scholar] [CrossRef]
- Rosa, L.H.; Zani, C.L.; Cantrell, C.L.; Duke, S.O.; Dijck, P.V.; Desideri, A.; Rosa, C.A. Fungi in Antarctica: Diversity, Ecology, Effects of Climate Change, and Bioprospection for Bioactive Compounds. In Fungi of Antarctica, 1st ed.; Rosa, L., Ed.; Springer: Cham, Switzerland, 2019; pp. 1–17. [Google Scholar]
- Arenz, B.E.; Blanchette, R.A.; Farrell, R.L. Fungal diversity in Antarctic soils. In Antarctic Terrestrial Microbiology, 1st ed.; Cowan, D.A., Ed.; Springer: Berlin, Germany, 2014; pp. 35–53. [Google Scholar]
- Canini, F.; Geml, J.; D’Acqui, L.P.; Buzzini, P.; Turchetti, B.; Onofri, S.; Ventura, S.; Zucconi, L. Fungal diversity and functionality are driven by soil texture in Taylor Valley, Antarctica. Fungal Ecol. 2021, 50, 101041. [Google Scholar] [CrossRef]
- Botnen, S.; Vik, U.; Carlsen, T.; Eidesen, P.B.; Davey, M.L.; Kauserud, H. Low host specificity of root-associated fungi at an Arctic site. Mol. Ecol. 2014, 23, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Gazis, R.; Skaltsas, D.; Chaverri, P.; Hibbett, D. Unexpected diversity of basidiomycetous endophytes in sapwood and leaves of Hevea. Mycologia 2015, 107, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Pegler, D.N.; Spooner, B.M.; Lewis Smith, R.I. Higher Fungi of Antarctica, the Subantarctic Zone and Falkland Islands. Kew Bull. 1980, 35, 499–562. [Google Scholar] [CrossRef]
- Held, B.W.; Blanchette, R.A. Deception Island, Antarctica, harbors a diverse assemblage of wood decay fungi. Fungal Biol. 2017, 121, 145–157. [Google Scholar] [CrossRef]
- De Giori, G.S.; de Valdez, G.F.; de Ruiz Holgado, A.P.; Oliver, G. Effect of pH and temperature on the proteolytic activity of lactic acid bacteria. J. Dairy Sci. 1985, 68, 2160–2164. [Google Scholar] [CrossRef]
- Pansare, A.C.; Venugopal, V.; Lewis, N.F. A note on nutritional influence on extracellular protease synthesis in Aeromonas hydrophila. J. Appl. Bacteriol. 1985, 58, 101–104. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Rhoden, S.A.; Mota, T.R.; Azevedo, J.L.; Pamphile, J.A.; de Souza, C.G.M.; Polizeli, M.L.T.M.; Bracht, A.; Peralta, R.M. Endophytic fungi: Expanding the arsenal of industrial enzyme producers. J. Ind. Microbiol. Biotechnol. 2014, 41, 1467–1478. [Google Scholar] [CrossRef]
- Shankar Naik, B.; Abrar, S.; Krishnappa, M. Industrially Important Enzymes from Fungal Endophytes. In Recent Advancement in White Biotechnology through Fungi; Yadav, A., Mishra, S., Singh, S., Gupta, A., Eds.; Springer: Cham, Switzerland, 2019; Volume 1, pp. 263–280. [Google Scholar]
- Kogel, K.-H.; Franken, P.; Hückelhoven, R. Endophyte or parasite—What decides? Curr. Opin. Plant Pathol. 2006, 9, 358–363. [Google Scholar] [CrossRef]
- Robl, D.; da Silva Delabona, P.; Mergel, C.M.; Rojas, J.D.; dos Santos Costa, P.; Pimentel, I.C.; Vicente, V.A.; da Cruz Pradella, J.G.; Padilla, G. The capability of endophytic fungi for production of hemicellulases and related enzymes. BMC Biotechnol. 2013, 13, 1–12. [Google Scholar] [CrossRef]
- Queiroz, C.B.D.; Santana, M.F. Prediction of the secretomes of endophytic and non-endophytic fungi reveals similarities in host plant infection and colonization strategies. Mycologia 2020, 112, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Cai, X.; Shao, C.; She, Z.; Xia, X.; Chen, Y.; Yang, J.; Zhou, S.; Lin, Y. Chemistry and weak antimicrobial activities of phomopsins produced by mangrove endophytic fungus Phomopsis sp. ZSU-H76. Phytochemistry 2008, 69, 1604–1608. [Google Scholar] [CrossRef] [PubMed]
- Zabalgogeazcoa, I.; Oleaga, A.; Pérez-Sánchez, R. Pathogenicity of endophytic entomopathogenic fungi to Ornithodoros erraticus and Ornithodoros moubata (acari: Argasidae). Vet. Parasitol. 2008, 158, 336–343. [Google Scholar] [CrossRef]
- Demain, A.L.; Velasco, J.; Adrio, J.L. Industrial mycology: Past, present, and future. In Handbook of Industrial Mycology, 1st ed.; An, Z., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 1–25. [Google Scholar]
- Bérdy, J. Bioactive Microbial Metabolites. A Personal View. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. 2012, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Flores-Vallejo, R.D.C.; Folch-Mallol, J.L.; Sharma, A.; Cardoso-Taketa, A.; Alvarez-Berber, L.; Villarreal, M.L. ITS2 ribotyping, in vitro anti-inflammatory screening, and metabolic profiling of fungal endophytes from the Mexican species Crescentia alata Kunth. S. Afr. J. Bot. 2020, 134, 213–224. [Google Scholar] [CrossRef]
- Oyetayo, O.V.; Nieto-Camacho, A.; Ramırez-Apana, T.M.; Baldomero, R.E.; Jimenez, M. Total Phenol, Antioxidant and Cytotoxic Properties of Wild Macrofungi Collected from Akure Southwest Nigeria. Jordan J. Biol. Sci. 2012, 6, 105–110. [Google Scholar] [CrossRef]
- Dixon, R.A.; Pasinetti, G.M. Flavonoids and Isoflavonoids: From Plant Biology to Agriculture and Neuroscience. Plant Physiol. 2010, 154, 453–457. [Google Scholar] [CrossRef]
- Hodgson, J.M.; Croft, K.D.; Puddey, I.B.; Mori, T.A.; Beilin, L.J. Soybean isoflavonoids and their metabolic products inhibit in vitro lipoprotein oxidation in serum. J. Nutr. Biochem. 1996, 7, 664–669. [Google Scholar] [CrossRef]
- Andersen, M.; Markham, K.R. Flavonoids, Chemistry, Biochemistry and Applications; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 1–1239. [Google Scholar]
- Abdel-Lateif, K.; Bogusz, D.; Hocher, V. The role of flavonoids in the establishment of plant roots endosymbioses with arbuscular mycorrhiza fungi, rhizobia and Frankia bacteria. Plant Signal. Behav. 2012, 7, 636–641. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Selassie, C.D.; Kapur, S.; Verma, R.P.; Rosario, M. Cellular Apoptosis and Cytotoxicity of Phenolic Compounds: A Quantitative Structure-Activity Relationship Study. J. Med. Chem. 2005, 48, 7234–7242. [Google Scholar] [CrossRef] [PubMed]
- Tapojyoti, S.; Swapan, K.G. Anti-cancer property of Lenzites betulina (L) Fr. on cervical cancer cell lines and its anti-tumor effect on HeLa-implanted mice. bioRxiv 2019. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, E.A.; Menchinskaya, E.S.; Pislyagin, E.A.; Trinh, P.T.H.; Ivanets, E.V.; Smetanina, O.F.; Yurchenko, A.N. Neuroprotective Activity of Some Marine Fungal Metabolites in the 6-Hydroxydopamin- and Paraquat-Induced Parkinson’s Disease Models. Mar. Drugs 2018, 16, 457. [Google Scholar] [CrossRef]
- Macedo, D.; Tavares, L.; McDougall, G.J.; Vicente Miranda, H.; Stewart, D.; Ferreira, R.B.; Tenreiro, S.; Outeiro, T.F.; Santos, C.N. (Poly)phenols protect from α-synuclein toxicity by reducing oxidative stress and promoting autophagy. Hum. Mol. Genet. 2015, 24, 1717–1732. [Google Scholar] [CrossRef]
- Polverino de Laureto, P.; Acquasaliente, L.; Palazzi, L. Polyphenols as potential therapeutic drugs in neurodegeneration. In Neuroprotection—New Approaches and Prospects, 1st ed.; Otero-Losada, M., Capani, F., Lloret, S.P., Eds.; IntechOpen: London, UK, 2019; Volume 3, pp. 37–67. [Google Scholar]
- Palazzi, L.; Leri, M.; Cesaro, S.; Stefani, M.; Bucciantini, M.; Polverino de Laureto, P. Insight into the molecular mechanism underlying the inhibition of α-synuclein aggregation by hydroxytyrosol. Biochem. Pharmacol. 2020, 173, 113722. [Google Scholar] [CrossRef]
- Weinreb, P.H.; Zhen, W.; Poon, A.W.; Conway, K.A.; Lansbury, P.T., Jr. NACP, a protein implicated in Alzheimer’s disease and learning, is natively unfolded. Biochemistry 1996, 35, 13709–13715. [Google Scholar] [CrossRef]
- Fongaro, B.; Cappelletto, E.; Sosic, A.; Spolaore, B.; Polverino de Laureto, P. 3,4-Dihydroxyphenylethanol and 3,4-dihydroxyphenylacetic acid affect the aggregation process of E46K variant of α-synuclein at different extent: Insights into the interplay between protein dynamics and catechol effect. Protein Sci. 2022, 31, e4356. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart, A.; et al. Environmental growth conditions of Trichoderma spp. affects indole acetic acid derivatives, volatile organic compounds, and plant growth promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, E.; Ortega-Amaro, M.A.; Salazar-Badillo, F.B.; Bautista, E.; Douterlungne, D.; Jiménez-Bremont, J.F. The Arabidopsis-Trichoderma interaction reveals that the fungal growth medium is an important factor in plant growth induction. Sci. Rep. 2018, 8, 16427. [Google Scholar] [CrossRef] [PubMed]
- Pelagio-Flores, R.; Esparza-Reynoso, S.; Garnica-Vergara, A.; López-Bucio, J.; Herrera-Estrella, A. Trichoderma-induced acidification Is an early trigger for changes in Arabidopsis root growth and determines fungal phytostimulation. Front. Plant Sci. 2017, 8, 822. [Google Scholar] [CrossRef] [PubMed]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299. [Google Scholar] [CrossRef]
- Bertrand, S.; Bohni, N.; Schnee, S.; Schumpp, O.; Gindro, K.; Wolfender, J.-L. Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol. Adv. 2014, 32, 1180–1204. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Haas, S.; Junker, C.; Andrée, N.; Schobert, M. Fungal endophytes are involved in multiple balanced antagonisms. Curr. Sci. 2015, 109, 39–45. [Google Scholar]
- Masachis, S.; Segorbe, D.; Turrà, D.; Leon-Ruiz, M.; Fürst, U.; El Ghalid, M.; Leonard, G.; López-Berges, M.S.; Richards, T.A.; Felix, G.; et al. A fungal pathogen secretes plant alkalinizing peptides to increase infection. Nat. Microbiol. 2016, 1, 16043. [Google Scholar] [CrossRef]
- Palmieri, D.; Vitale, S.; Lima, G.; Di Pietro, A.; Turrà, D. A bacterial endophyte exploits chemotropism of a fungal pathogen for plant colonization. Nat. Commun. 2020, 11, 5264. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Isolate | Activity | |||||
|---|---|---|---|---|---|---|
| Amylolytic | Lipolytic | Cellulolytic | Pectinolytic | Proteolytic (Skimmed Milk) | Proteolytic (Gelatin) | |
| Trametes sp. S1.OA.A_F2 | + | + | +++ | + | + | + |
| Trametes sp. S2.OTC.C_F5 | + | + | +++ | + | + | − |
| Trametes sp. S2.OA.A_F5 | +++ | + | + | + | + | − |
| Trametes sp. S2.OA.C_F6 | + | + | − | + | + | − |
| Lenzites sp. S2.OTC.C_F8 | − | + | − | + | + | − |
| Lenzites sp. S3.OA.B_F6 | +++ | + | + | − | + | − |
| Sistotrema sp. S1.OA.C_F2 | + | − | + | − | − | − |
| Peniophora sp. S3.OTC.C_F1 | − | − | + | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertini, L.; Perazzolli, M.; Proietti, S.; Capaldi, G.; Savatin, D.V.; Bigini, V.; Longa, C.M.O.; Basaglia, M.; Favaro, L.; Casella, S.; et al. Biodiversity and Bioprospecting of Fungal Endophytes from the Antarctic Plant Colobanthus quitensis. J. Fungi 2022, 8, 979. https://doi.org/10.3390/jof8090979
Bertini L, Perazzolli M, Proietti S, Capaldi G, Savatin DV, Bigini V, Longa CMO, Basaglia M, Favaro L, Casella S, et al. Biodiversity and Bioprospecting of Fungal Endophytes from the Antarctic Plant Colobanthus quitensis. Journal of Fungi. 2022; 8(9):979. https://doi.org/10.3390/jof8090979
Chicago/Turabian StyleBertini, Laura, Michele Perazzolli, Silvia Proietti, Gloria Capaldi, Daniel V. Savatin, Valentina Bigini, Claudia Maria Oliveira Longa, Marina Basaglia, Lorenzo Favaro, Sergio Casella, and et al. 2022. "Biodiversity and Bioprospecting of Fungal Endophytes from the Antarctic Plant Colobanthus quitensis" Journal of Fungi 8, no. 9: 979. https://doi.org/10.3390/jof8090979
APA StyleBertini, L., Perazzolli, M., Proietti, S., Capaldi, G., Savatin, D. V., Bigini, V., Longa, C. M. O., Basaglia, M., Favaro, L., Casella, S., Fongaro, B., Polverino de Laureto, P., & Caruso, C. (2022). Biodiversity and Bioprospecting of Fungal Endophytes from the Antarctic Plant Colobanthus quitensis. Journal of Fungi, 8(9), 979. https://doi.org/10.3390/jof8090979