
Phytophthora × cambivora as a Major Factor Inciting the Decline of European Beech in a Stand within the Southernmost Limit of Its Natural Range in Europe

 ,
,  , ,
, , 
Abstract
1. Introduction
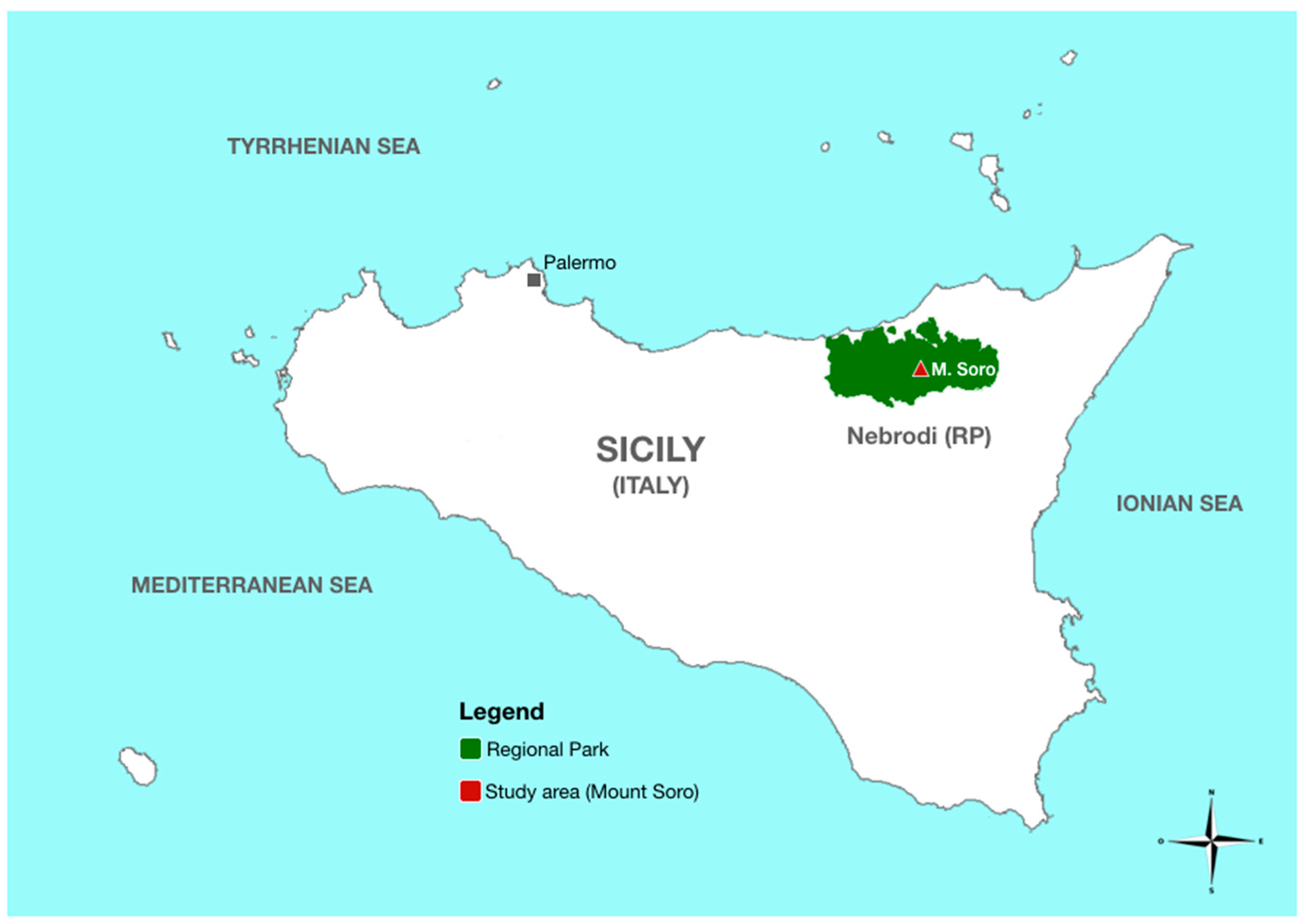
2. Materials and Methods
2.1. Sampling, Isolation, and Quantitative Determination of Phytophthora
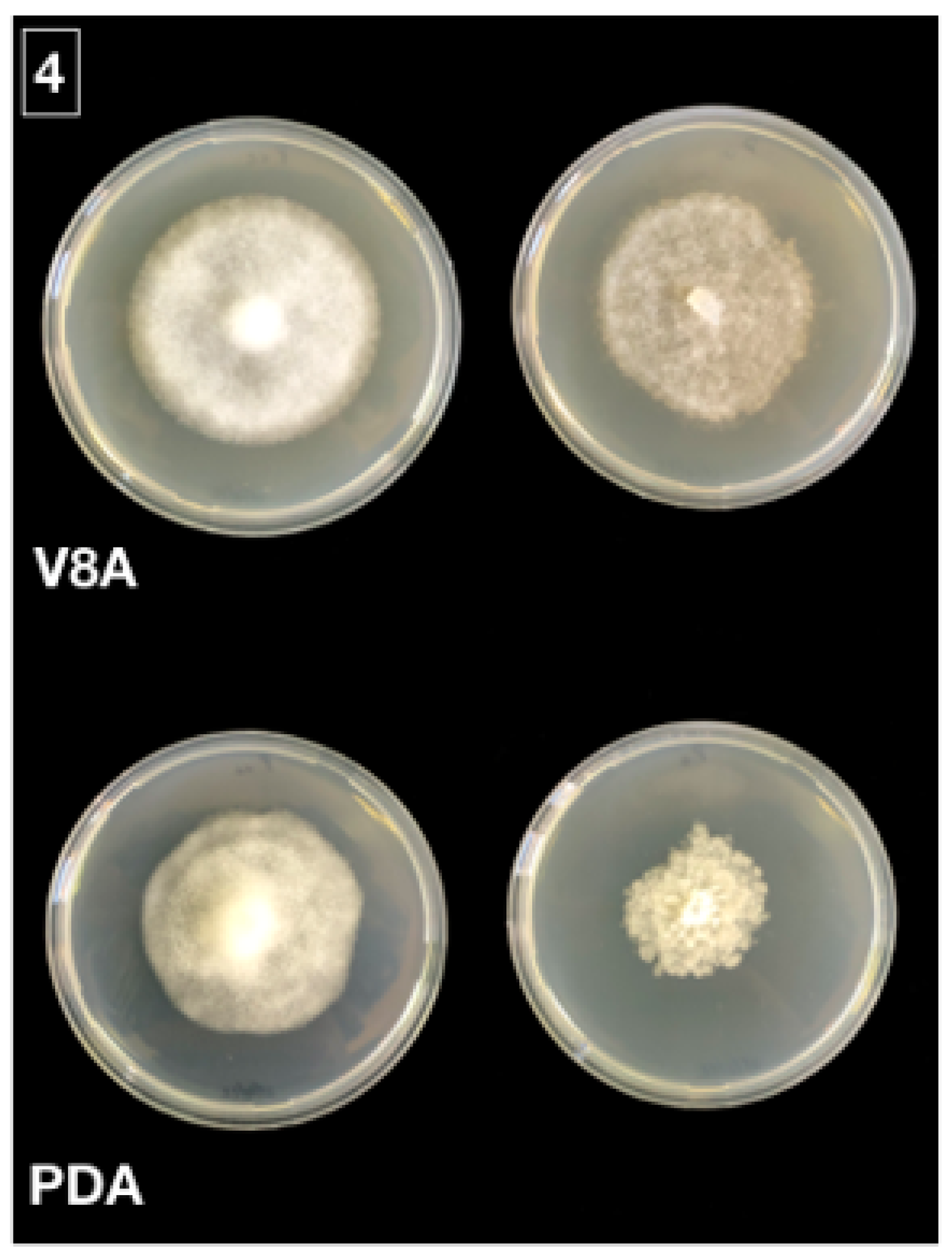
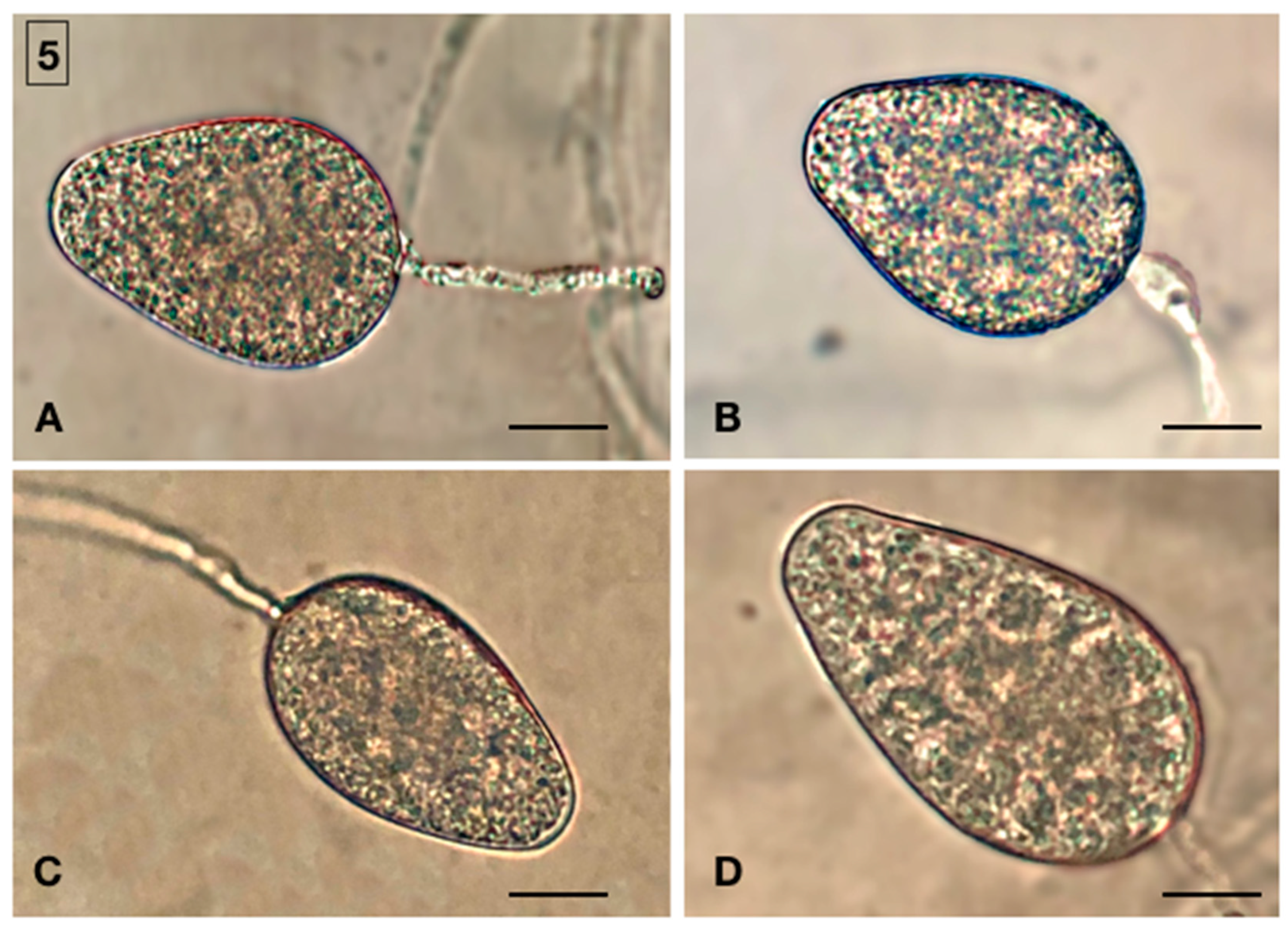
2.2. Morphological Identification of Isolates
2.3. Molecular Identification of Isolates
2.4. Mating Type of Isolates
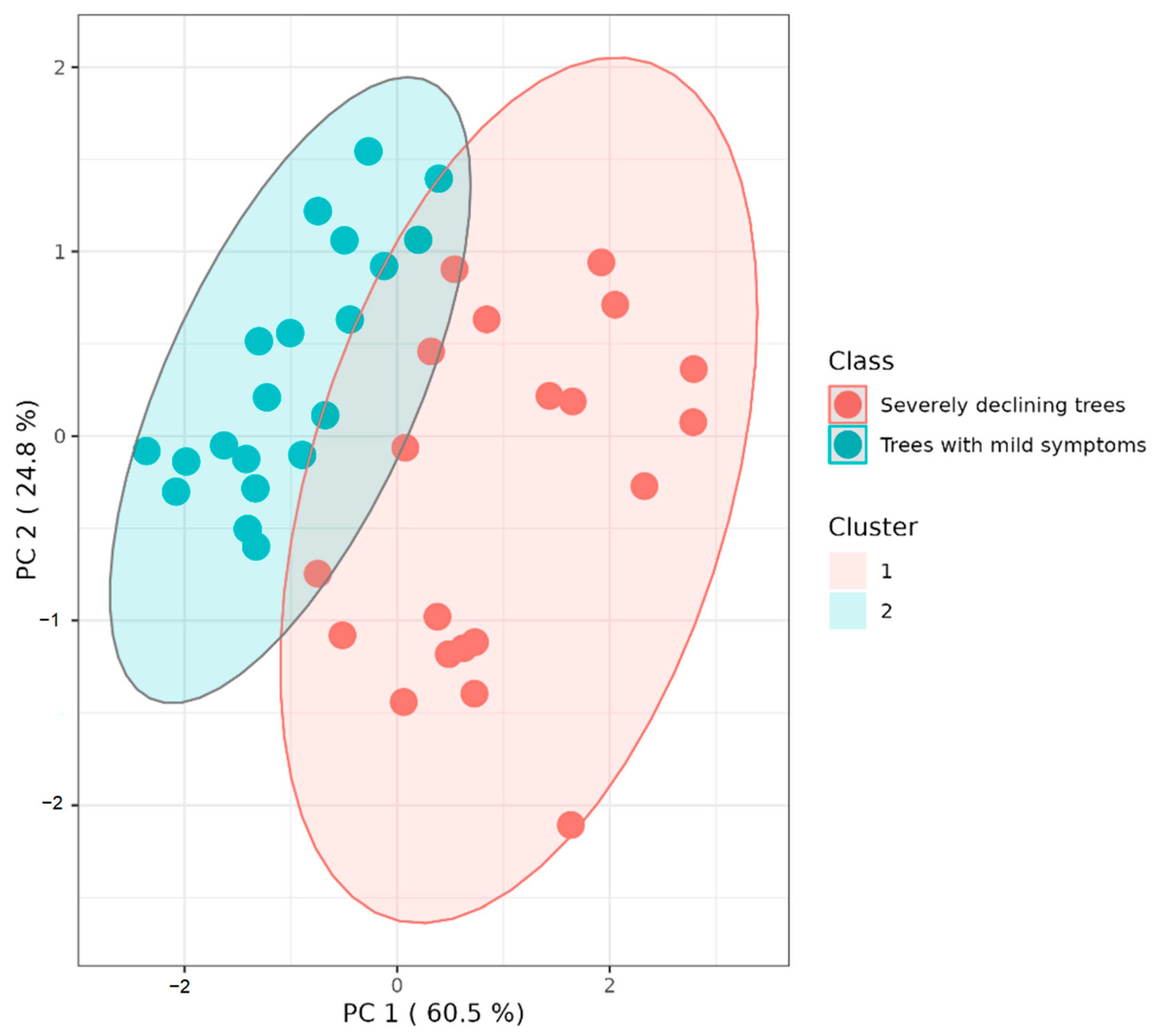
2.5. Statistical Analysis of Results
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudley, N.; Shadie, P.; Stolton, S. Guidelines for Applying Protected Areas Management Categories Including Iucn Wcpa Best Practice Guidance on Recognising Protected Areas and Assigning Management Categories and Governance Type; IUCN: Gland, Switzerland, 2013; p. 31. [Google Scholar]
- Bolte, A.; Czajkowshi, T.; Kompa, T. The north-eastern distribution range of European beech—A review. Forestry 2007, 80, 413–429. [Google Scholar] [CrossRef]
- Jung, T. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. For. Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Jung, T.; Hudler, G.W.; Griffiths, H.M.; Fleischmann, F.; Oßwald, W. Involvement of Phytophthora spp. in the decline of European beech in Europe and the USA. Mycologist 2005, 19, 159–166. [Google Scholar]
- Nelson, A.H.; Weiland, J.E.; Hudler, G.W. Prevalence, distribution and identification of Phytophthora species from bleeding canker on European beech. J. Environ. Hort. 2010, 28, 150–158. [Google Scholar] [CrossRef]
- Stepniewska, H.; Dłuszyňski, J. Incidence of Phytophthora cambivora in bleeding lesions on beech stems in selected forest stands in south-eastern Poland. Phytopathologia 2010, 56, 39–51. [Google Scholar]
- Milenković, I.; Keca, N.; Karadžić, D.; Nowakowska, J.A.; Borys, M.; Sikora, K.; Oszako, T. Incidence of Phytophthora species in beech stands in Serbia. Folia For. Pol. 2012, 54, 223–232. [Google Scholar]
- Telfer, K.H.; Brurberg, M.B.; Herrero, M.L.; Stensvand, A.; Talgø, V. Phytophthora cambivora found on beech in Norway. For. Pathol. 2015, 45, 415–425. [Google Scholar] [CrossRef]
- Jung, T.; Jung, M.H.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef]
- Corcobado, T.; Cech, T.L.; Brandstetter, M.; Daxer, A.; Hüttler, C.; Kudláček, T.; Jung, M.H.; Jung, T. Decline of European Beech in Austria: Involvement of Phytophthora spp. and contributing biotic and abiotic factors. Forests 2020, 11, 895. [Google Scholar]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Horta Jung, M.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and distribution of Phytophthora species in protected natural areas in Sicily. Forests 2019, 10, 259. [Google Scholar]
- Cacciola, S.O.; Gullino, M.L. Emerging and re-emerging fungus and oomycete soil-borne plant diseases in Italy. Phytopathol. Mediterr. 2019, 58, 451–472. [Google Scholar]
- Vettraino, A.M.; Morel, O.; Perlerou, C.; Robin, C.; Diamandis, S.; Vannini, A. Occurrence and distribution of Phytophthora species associated with ink disease of chestnut in Europe. Eur. J. Plant Pathol. 2005, 111, 169–180. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Natili, G.; Anselmi, N.; Vannini, A. Recovery and pathogenicity of Phytophthora species associated with a resurgence of ink disease in Castanea sativa in Italy. Plant Path. 2001, 50, 90–96. [Google Scholar] [CrossRef]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [PubMed]
- Seddaiu, S.; Brandano, A.; Ruiu, P.A.; Sechi, C.; Scanu, B. An overview of Phytophthora species inhabiting declining Quercus suber stands in Sardinia (Italy). Forests 2020, 11, 971. [Google Scholar] [CrossRef]
- Schomacher, M.E.; Zarnoch, S.J.; Bechtold, W.A.; Latelle, D.J.; Burkman, W.G.; Cox, S.M. Crown-Condition Classification: A Guide to Data Collection and Analysis; General Technical Report SRS 102; United States Department of Agriculture—Forest Service, Southern Research Station: Asheville, NC, USA, 2007; p. 78.
- Eichhorn, J.; Roskams, P.; Potočić, N.; Timmermann, V.; Ferretti, M.; Mues, V.; Szepesi, A.; Durrant, D.; Seletković, I.; Schröck, H.-W.; et al. Part IV: Visual assessment of crown condition and damaging agents. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 50, + Annex; Available online: http://www.icp-forests.org/manual.htm (accessed on 20 May 2021).
- Riolo, M.; Aloi, F.; La Spada, F.; Sciandrello, S.; Moricca, S.; Santilli, E.; Pane, A.; Cacciola, S.O. Diversity of Phytophthora communities across different types of Mediterranean vegetation in a nature reserve area. Forests 2020, 11, 853. [Google Scholar] [CrossRef]
- Jung, T.; Horta Jung, M.; Scanu, B.; Seress, D.; Kovács, D.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.T.; Chandelier, A.; Heungens, A.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia 2017, 38, 100–135. [Google Scholar] [CrossRef] [PubMed]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First report of root rot caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef] [PubMed]
- Cooke, D.E.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Kroon, L.P.N.M.; Bakker, F.T.; van den Bosch, G.B.M.; Bonants, P.J.M.; Flier, W.G. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef] [PubMed]
- FinchTV v.1.4.0. Available online: https://digitalworldbiology.com/FinchTV (accessed on 20 May 2021).
- BLAST Searches. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 20 May 2021).
- Phytophthora Database. Available online: http://www.phytophthoradb.org/ (accessed on 20 May 2021).
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 20 May 2021).
- Mammella, M.A.; Cacciola, S.O.; Martin, F.; Schena, L. Genetic characterization of Phytophthora nicotianae by the analysis of polymorphic regions of the mitochondrial DNA. Fungal Biol. 2011, 115, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Mammella, M.A.; Martin, F.N.; Cacciola, S.O.; Coffey, M.D.; Faedda, R.; Schena, L. Analyses of the population structure in a global collection of Phytophthora nicotianae isolates inferred from mitochondrial and nuclear DNA sequences. Phytopathology 2013, 103, 610–622. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.H.; Chang, H.S.; Su, H.J. Isolates of Phytophthora cinnamomi from Taiwan as evidence for an Asian origin of the species. Trans. Br. Mycol. Soc. 1978, 71, 3. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster Analysis Basics and Extensions, R Package Version 20.6. Available online: https://CRAN.R-project.org/package=cluster (accessed on 20 May 2021).
- Grünwald, N.J.; Martin, F.N.; Larsen, M.M.; Sullivan, C.M.; Press, C.M.; Coffey, M.D.; Hansen, E.M.; Parke, J.L. Phytophthora-ID.org: A sequence-based Phytophthora identification tool. Plant Dis. 2011, 95, 337–342. [Google Scholar] [CrossRef]
- Corcobado, T.; Vivas, M.; Moreno, G.; Solla, A. Ectomycorrhizal symbiosis in declining and non-declining Quercus ilex trees infected with or free of Phytophthora cinnamomi. For. Ecol. Manag. 2014, 324, 72–80. [Google Scholar] [CrossRef]
- Day, W.R. Root-rot of sweet chestnut and beech caused by species of Phytophthora. I. Cause and symptoms of disease: Its relation to soil conditions. Forestry 1938, 12, 101–116. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Motta, E.; Raudino, F.; Chimento, A.; Pane, A.; Magnano di San Lio, G. Phytophthora pseudosyringae the causal agent of bleeding cankers of beech in central Italy. J. Plant Pathol. 2005, 87, 289. [Google Scholar]
- Motta, E.; Annesi, T.; Pane, A.; Cooke, D.E.L.; Cacciola, S.O. A new Phytophthora sp. causing a basal canker on beech in Italy. Plant Dis. 2003, 87, 1005. [Google Scholar]
- Diana, G.; Pane, A.; Raudino, F.; Cooke, D.E.L.; Cacciola, S.O.; Magnano di San Lio, G. 2006:A decline of beech trees caused by Phytophthora pseudosyringae in central Italy. In Progress in Research on Phytophthora Diseases of Forest Trees, Proceedings of the 3rd International IUFRO Working Party 7.02.09 Meeting, Freising, Germany, 11–17 September 2004; Brasier, C., Jung, T., Osswald, W., Eds.; Forest Research: Farnham, UK, 2006; pp. 142–144. ISBN 0-85538-721-1. [Google Scholar]
- Jung, T.; Nechwatal, J.; Cooke, D.E.L.; Hartmann, G.; Blaschke, M.; Oßwald, W.F.; Duncan, J.M.; Delatour, C. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol. Res. 2003, 107, 772–789. [Google Scholar] [CrossRef]
- Linzer, R.E.; Rizzo, D.M.; Cacciola, S.O.; Garbelotto, M. AFLPs detect low genetic diversity for Phytophthora nemorosa and P. pseudosyringae in the US and Europe. Mycol. Res. 2009, 113, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Osswald, W. Involvement of Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Benson, D.M. Residual activity of metalaxyl and population dynamic of Phytophthora cinnamomi in landscape beds of azalea. Plant Dis. 1987, 71, 886–891. [Google Scholar] [CrossRef]
- Sandler, H.A.; Timmer, L.W.; Graham, J.H.; Zitko, S.E. Effect of fungicide applications on populations of Phytophthora parasitica and on feeder root densities and fruit yields of citrus trees. Plant Dis. 1989, 73, 902–906. [Google Scholar] [CrossRef]
- Timmer, L.W.; Sandler, H.A.; Graham, J.H.; Zitko, S.E. Sampling citrus orchards in Florida to estimate populations of Phytophthora parasitica. Phytopathol. 1988, 78, 940–944. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Magnano di San Lio, G. Management of citrus diseases caused by Phytophthora spp. In Integrated Management of Diseases Caused by Fungi, Phytoplasma and Bacteria; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 61–84. ISBN 978-1-4020-8571-0. [Google Scholar]
- Nagle, A.M.; Long, R.P.; Madden, L.V.; Bonello, P. Association of Phytophthora cinnamomi with white oak decline in Southern Ohio. Plant Dis. 2010, 94, 1026–1034. [Google Scholar] [CrossRef]
- Balci, Y.; Long, R.P.; Mansfield, M.; Balser, D.; MacDonald, W.L. Involvement of Phytophthora species in white oak (Quercus alba) decline in southern Ohio. For. Pathol. 2010, 40, 430–442. [Google Scholar] [CrossRef]
- McConnel, M.E.; Balci, Y. Phytophthora cinnamomi as a contributor to white oak decline in mid-atlantic United States forests. Plant Dis. 2014, 98, 319–327. [Google Scholar] [CrossRef]
- Riolo, M.; La Spada, F.; Aloi, F.; Giusso del Galdo, G.; Santilli, E.; Pane, A.; Cacciola, S.O. Phytophthora Diversity in a sentinel arboretum and in a nature reserve area. Biol. Life Sci. Forum 2021, 4, 51. [Google Scholar]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe Environmental constraints including climate change. Ann. Sci. For. 1996, 53, 347–358. [Google Scholar]
- Moreira, A.C.; Martins, J.M.S. Influence of site factors on the impact of Phytophthora cinnamomi in cork oak stands in Portugal. Forest Pathol. 2005, 35, 145–162. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Jung, T.; Cooke, D.E.L.; Blaschke, H.; Duncan, J.M.; Oßwald, W. Phytophthora quercina sp. nov., causing root rot of European oaks. Mycol. Res. 1999, 103, 785–798. [Google Scholar] [CrossRef]
- Dunstan, W.A.; Rudman, T.; Shearer, B.L.; Moore, N.A.; Paap, T.; Calver, M.C.; Dell, B.; Hardy, G.E.S.J. Containment and spot eradication of a highly destructive, invasive plant pathogen (Phytophthora cinnamomi) in natural ecosystems. Biol. Invasions 2010, 12, 913–925. [Google Scholar] [CrossRef]
- Serrano, M.S.; Rios, P.; Gonzales, M.; Sanchez, M.E. Experimental minimum threshold for Phytophthora cinnamomi root disease expression on Quercus suber. Phytopath. Medit. 2015, 54, 461–464. [Google Scholar]
- Frisullo, S.; Lima, G.; Magnano di San Lio, G.; Camele, I.; Melissano, L.; Puglisi, I.; Pane, A.; Agosteo, G.E.; Prudente, L.; Cacciola, S.O. Phytophthora cinnamomi involved in the decline of holm oak (Quercus ilex) stands in southern Italy. For. Sci. 2018, 64, 290–298. [Google Scholar] [CrossRef]
- Brasier, C.M.; Beales, P.A.; Denman, S.; Rose, J. Phytophthora kernoviae sp. nov., an invasive pathogen causing bleeding stem lesions on forest trees and foliar necrosis of ornamentals in the UK. Mycol. Res. 2005, 109, 853–859. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press—The American Phytopathological Society: St. Paul, MN, USA, 1996; pp. 258–261; 334–335. [Google Scholar]
- Cleary, M.; Blomquist, M.; Ghasemkhani, M.; Witzell, J. First report of Phytophthora gonapodyides causing stem canker on European beech (Fagus sylvatica) in southern Sweden. Plant Dis. 2016, 100, 2174. [Google Scholar] [CrossRef]
- Solís-García, I.A.; Ceballos-Luna, O.; Cortazar-Murillo, E.M.; Desgarennes, D.; Garay-Serrano, E.; Patiño-Conde, V.; Guevara-Avendaño, E.; Méndez-Bravo, A.; Reverchon, F. Phytophthora root rot modifies the composition of the avocado rhizosphere microbiome and increases the abundance of opportunistic fungal pathogens. Front. Microbiol. 2021, 11, 574110. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Phytophthora Species | Source | Accession Numbers | |
|---|---|---|---|---|
| ITS | Cox I | |||
| MSK1A | Phytophthora × cambivora | Soil and roots a | ON186623 | ON227269 |
| MSK6C | P.× cambivora | Soil and roots | ON186624 | ON227270 |
| MSQ9B | P. × cambivora | Soil and roots | ON186625 | ON227271 |
| MSK3C | P. × cambivora | Soil and roots | ON186626 | ON227272 |
| MSK11A | P. × cambivora | Soil and roots | ON186627 | ON227273 |
| MSK12A | P. × cambivora | Soil and roots | ON186628 | ON227274 |
| MSQ4A | P. × cambivora | Soil and roots | ON186629 | ON227275 |
| MSK6B | P. × cambivora | Soil and roots | ON186630 | ON227276 |
| MSK14C | P. × cambivora | Soil and roots | ON186631 | ON227277 |
| MSQ2C | P. × cambivora | Soil and roots | ON186632 | ON227278 |
| MSC1A | P gonapodyides | Stem bark | ON186633 | ON227279 |
| MSC2B | P. gonapodyides | Stem bark | ON186634 | ON227280 |
| MSC1D | P. gonapodyides | Stem bark | ON186635 | ON227281 |
| MSC5E | P. gonapodyides | Stem bark | ON186636 | ON227282 |
| MSC4D | P. gonapodyides | Stem bark | ON186637 | ON227283 |
| MSC2C | P. gonapodyides | Stem bark | ON186638 | ON227284 |
| MSC7E | P. gonapodyides | Stem bark | ON186639 | ON227285 |
| MSC3A | P. gonapodyides | Stem bark | ON186640 | ON227286 |
| MSC9B | P. gonapodyides | Stem bark | ON186641 | ON227287 |
| MSC8E | P. gonapodyides | Stem bark | ON186642 | ON227288 |
| Foliage Transparency | Inoculum Density (cfu/g of Soil) | Proportion of Infected Fibrous Roots (%) | |
|---|---|---|---|
| Proportion of infected fibrous roots (%) | 0.466 ** | 0.508 ** | 1 |
| Foliage transparency | 1 | 0.548 ** | 0.466 ** |
| Inoculum density (cfu/g of soil) | 0.548 ** | 1 | 0.508 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riolo, M.; Aloi, F.; Conti Taguali, S.; Pane, A.; Franco, M.; Cacciola, S.O. Phytophthora × cambivora as a Major Factor Inciting the Decline of European Beech in a Stand within the Southernmost Limit of Its Natural Range in Europe. J. Fungi 2022, 8, 973. https://doi.org/10.3390/jof8090973
Riolo M, Aloi F, Conti Taguali S, Pane A, Franco M, Cacciola SO. Phytophthora × cambivora as a Major Factor Inciting the Decline of European Beech in a Stand within the Southernmost Limit of Its Natural Range in Europe. Journal of Fungi. 2022; 8(9):973. https://doi.org/10.3390/jof8090973
Chicago/Turabian StyleRiolo, Mario, Francesco Aloi, Sebastiano Conti Taguali, Antonella Pane, Massimo Franco, and Santa Olga Cacciola. 2022. "Phytophthora × cambivora as a Major Factor Inciting the Decline of European Beech in a Stand within the Southernmost Limit of Its Natural Range in Europe" Journal of Fungi 8, no. 9: 973. https://doi.org/10.3390/jof8090973
APA StyleRiolo, M., Aloi, F., Conti Taguali, S., Pane, A., Franco, M., & Cacciola, S. O. (2022). Phytophthora × cambivora as a Major Factor Inciting the Decline of European Beech in a Stand within the Southernmost Limit of Its Natural Range in Europe. Journal of Fungi, 8(9), 973. https://doi.org/10.3390/jof8090973