

Morphological and Molecular Characterization of Calonectria foliicola Associated with Leaf Blight on Rubber Tree (Hevea brasiliensis) in Thailand

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Pathogenicity Test
2.3. Molecular Study
2.4. Morphological Study
2.5. Statistical Analysis
3. Results
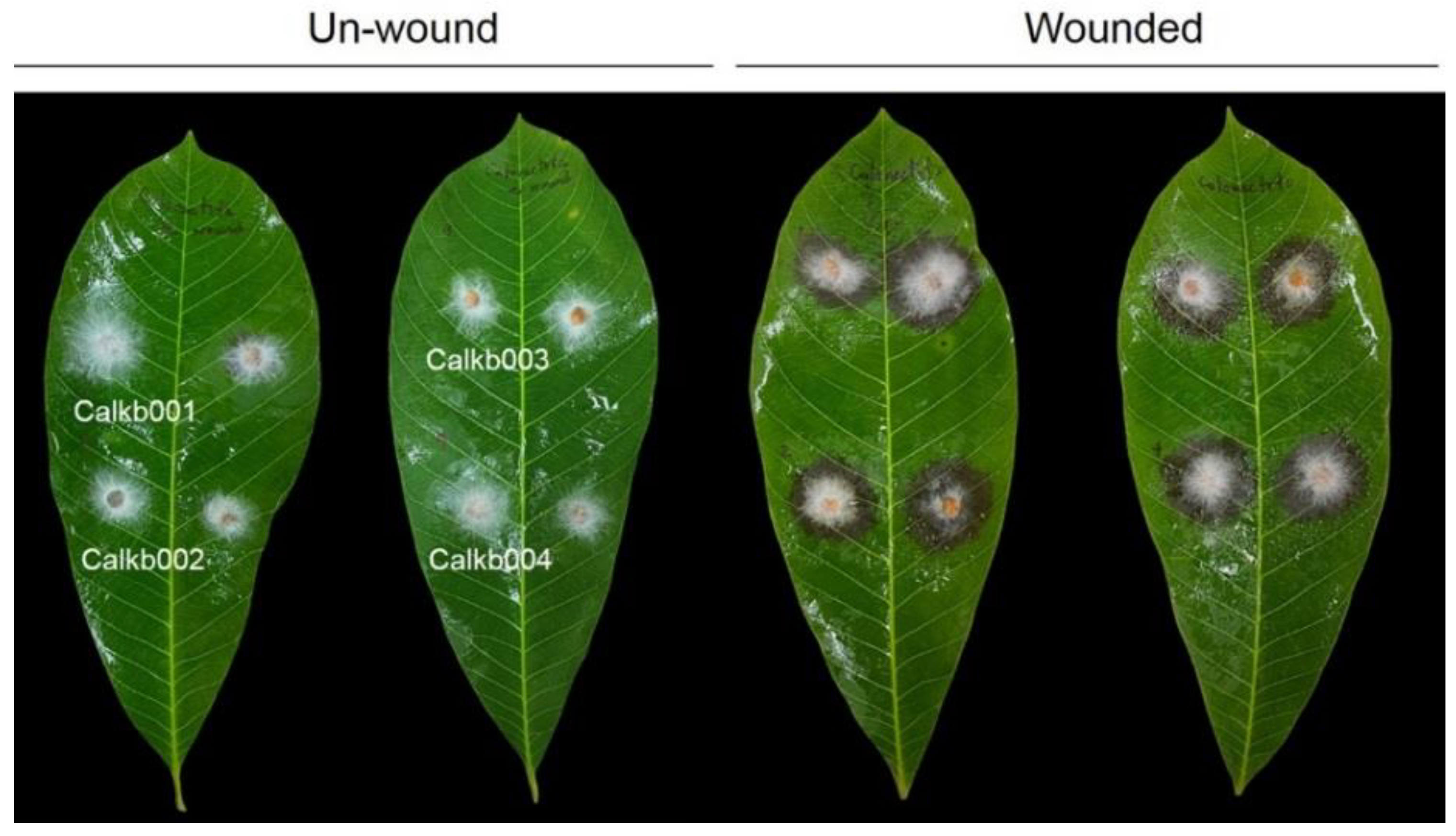
3.1. Symptom Recognition and Fungal Isolates
3.2. Pathogenicity of Fungi
3.3. Molecular Identification
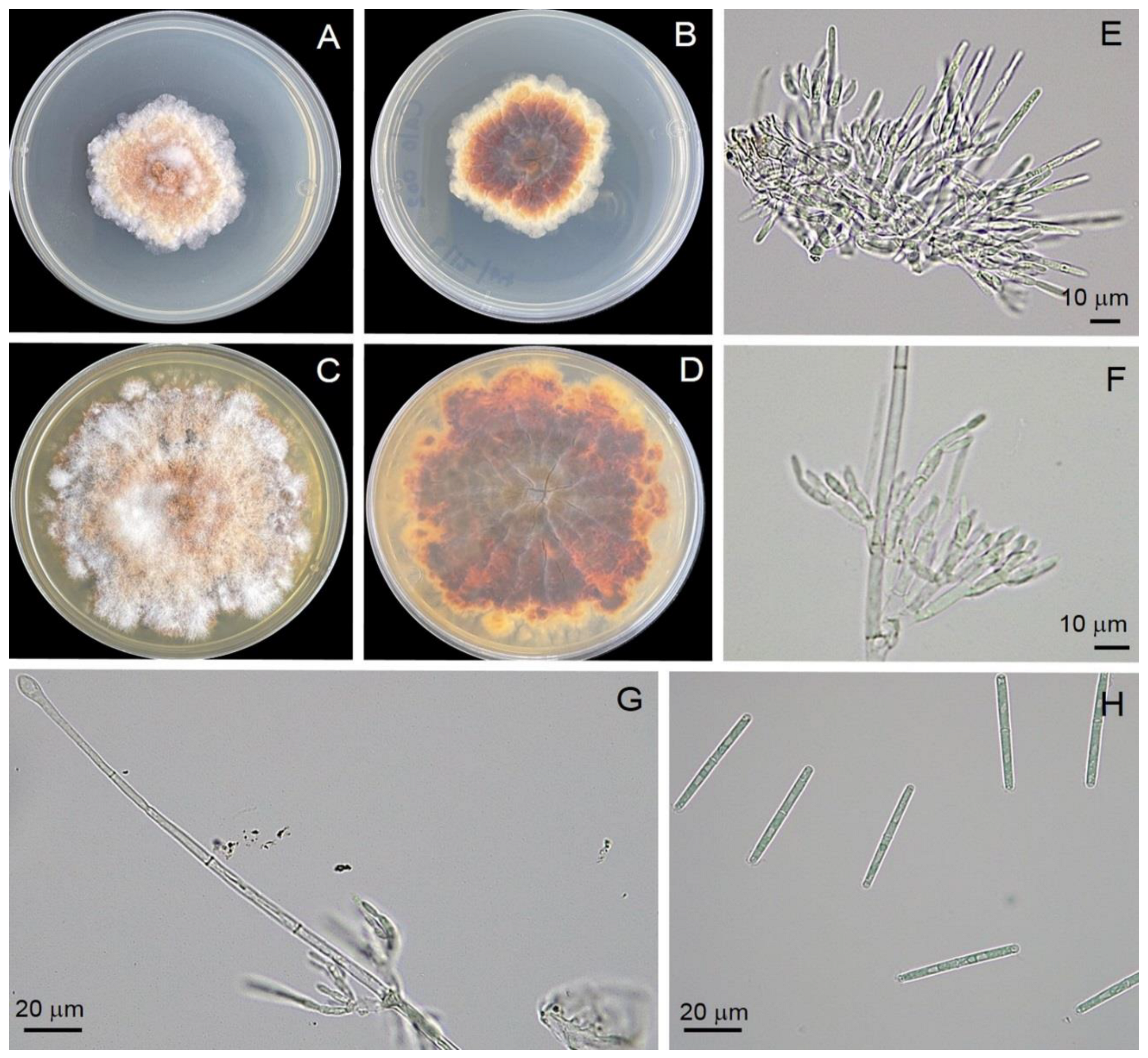
3.4. Morphology Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carr, M.K.V. The water relations of rubber (Hevea brasiliensis): A review. Exp. Agric. 2012, 48, 176–193. [Google Scholar] [CrossRef]
- Chen, J.-W.; Cao, K.-F. A possible link between hydraulic properties and leaf habits in Hevea brasiliensis. Funct. Plant Biol. 2015, 42, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Archer, B.L.; Audeley, B.G. The biosynthesis of rubber. Biochem. J. 1963, 89, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Backhaus, R.A. Rubber formation in plants: A mini-review. Israel J. Bot. 1985, 34, 283–293. [Google Scholar]
- Asawatreratanakul, K.; Zhang, Y.W.; Wititsuwannakul, D.; Wititsuwannakul, R.; Takahashi, S.; Rattanapittayaporn, A.; Koyama, T. Molecular cloning, expression and characterization of cDNA encoding cis-prenyltransferases from Hevea brasiliensis. A key factor participating in natural rubber biosynthesis. Eur. J. Biochem. 2003, 270, 4671–4680. [Google Scholar] [CrossRef]
- Cesar, M.B.; Borges, F.A.; Bilck, A.P.; Yamashita, F.; Paulino, C.G.; Herculano, R.D. Development and characterization of natural rubber latex and polylactic acid membranes for biomedical application. J. Polym. Environ. 2020, 28, 220–230. [Google Scholar] [CrossRef]
- Guerra, N.B.; Sant’Ana Pegorin, G.; Boratto, M.H.; de Barros, N.R.; de Oliveira Graeff, C.F.; Herculano, R.D. Biomedical applications of natural rubber latex from the rubber tree Hevea brasiliensis. Mater. Sci. Eng. C 2021, 126, 112126. [Google Scholar] [CrossRef]
- Limkaisang, S.; Kom-un, S.; Furtado, E.L.; Liew, K.W.; Salleh, B.; Sato, Y. Molecular phylogenetic and morphological analyses of Oidium heveae, a powdery mildew of rubber tree. Mycoscience 2005, 46, 220–226. [Google Scholar] [CrossRef]
- Li, Z.; Fox, J.M. Mapping rubber tree growth in mainland Southeast Asia using time-series MODIS 250 m NDVI and statistical data. Appl. Geogr. 2012, 32, 420–432. [Google Scholar] [CrossRef]
- Jinji, P.; Xin, Z.; Yangxian, Q.; Yixian, X.; Huiqiang, Z.; He, Z. First record of Corynespora leaf fall disease of Hevea rubber tree in China. Australas. Plant Dis. Notes 2007, 2, 35–36. [Google Scholar] [CrossRef]
- Li, B.; Liu, X.; Cai, J.; Feng, Y.; Huang, G. First report on Neopestalotiopsis aotearoa of rubber tree in China. Plant Dis. 2021, 105, 1223. [Google Scholar] [CrossRef]
- Liu, X.; Li, B.; Cai, J.; Zheng, X.; Feng, Y.; Huang, G. Colletotrichum species causing anthracnose of rubber trees in China. Sci. Rep. 2018, 8, 10435. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J.; Bai, Y.H.; Xu, K.C.; Zhang, R.Q.; Huang, Q. First report of wilt of rubber tree caused by Chalaropsis thielavioides in China. Plant Dis. 2021, 105, 1221. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Peng, Y.; Liu, Y.; Wang, M.; Yang, Y.; Zhang, Y. First report of Bipolaris bicolor causing a leaf spot disease on rubber tree. J. Phytopath. 2019, 167, 553–557. [Google Scholar] [CrossRef]
- Wonglom, P.; Thithuan, N.; Bunjongsiri, P.; Sunpapao, A. Plant-parasitic algae (Cephaleuros spp.) in Thailand, including four new records. Pac. Sci. 2018, 72, 363–371. [Google Scholar] [CrossRef]
- Thithuan, N.; Bunjonsiri, P.; Sunpapao, A. Morphology and behavior of gametes and zoospores from the plant-parasitic green algae, Cephaleuros (Chlorophyta, Ulvophyceae). Pac. Sci. 2019, 73, 403–410. [Google Scholar] [CrossRef]
- Pitaloka, M.K.; Petcharat, V.; Arikit, S.; Sunpapao, A. Cephaleuros virescens, the cause of an algal leaf spot on Para rubber in Thailand. Australas. Plant Dis. Notes 2015, 10, 4. [Google Scholar] [CrossRef]
- Laohasakul, B.; Boonyapipat, P.; Plodpai, P. First report of Phytophthora citrophthora causing leaf fall of Para rubber tree (Hevea brasiliensis) in Thailand. Plant Dis. 2017, 101, 1057. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Chairin, T.; Thaochan, N.; Sunpapao, A. Identification and characterization of Neopestalotiopsis fungi associated with a novel leaf fall disease of rubber trees (Hevea brasiliensis) in Thailand. J. Phytopathol. 2020, 168, 416–427. [Google Scholar] [CrossRef]
- Wu, R.H.; Zhnag, Y.; Li, Z.P. First report of leaf spot on rubber tree caused by Lasiodiplodia pseudotheobromae in China. Plant Dis. 2019, 103, 766. [Google Scholar] [CrossRef]
- Narayanan, C.; Reju, M.J. Variation in susceptibility to Phyllosticta capitalensis-associated leaf disease among inter-specific hybrids, half-sibs and high-yielding clones of Para rubber tree (Hevea). Plant Pathol. Quar. 2020, 10, 100–110. [Google Scholar] [CrossRef]
- Liu, Q.L.; Chen, S.F. Two novel species of Calonectria isolated from soil in a natural forest in China. MycoKeys 2017, 26, 25–60. [Google Scholar] [CrossRef]
- Liu, L.; Wu, W.; Chen, S. Species diversity and distribution characteristics of Calonectria in five soil layers in a Eucalyptus plantation. J. Fungi 2021, 7, 857. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Lombard, L.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Novel species of Calonectria associated with Eucalyptus leaf blight in southeast China. Persoonia 2011, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.C.; Chen, S.F. Calonectria pentaseptata causes severe leaf disease of cultivated Eucalyptus on the Leizhou Peninsula of southern China. Plant Dis. 2019, 104, 493–509. [Google Scholar] [CrossRef]
- Miranda, I.D.S.; Auer, C.G.; dos Santos, Á.F.; Ferreira, M.A.; Tambarussi, E.V.; da Silva, R.A.F.; Rezende, E.H. Occurrence of Calonectria leaf blight in Eucalyptus benthamii progenies and potential for disease resistance. Trop. Plant Pathol. 2021, 46, 254–264. [Google Scholar] [CrossRef]
- Daengsuwan, W.; Wonglom, P.; Sunpapao, A. First report of Lasiodiplodia theobromae causing spadix rot in Anthurium andraeanum. J. Phytopathol. 2020, 168, 129–133. [Google Scholar] [CrossRef]
- Dy, K.S.; Wonglom, P.; Pornsuriya, C.; Sunpapao, A. Morphological, molecular identification and pathogenicity of Neoscytalidium dimidiatum causing stem canker of Hylocereus polyrhizus in southern Thailand. Plants 2022, 11, 504. [Google Scholar] [CrossRef]
- Daengsuwan, W.; Wonglom, P.; Arikit, S.; Sunpapao, A. Morphological and molecular identification of Neopestalotiopsis clavispora causing flower blight on Anthurium andraeanum in Thailand. Hortic. Plant J. 2021, 7, 573–578. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Kunasakdakul, K.; Wonglom, P.; Dy, K.S.; Sunpapao, A. Morphological and molecular studies of a rare mucoralean species causing flower rot in Hylocereus polyrhizus. J. Phytopathol. 2022, 170, 217–220. [Google Scholar] [CrossRef]
- Sunpapao, A.; Suwannarach, N.; Kumla, J.; Dumhai, R.; Riangwong, K.; Sanguansub, S.; Wanchana, S.; Arikit, S. Morphological and molecular identification of plant pathogenic fungi associated with dirty panicle disease in coconuts (Cocos nucifera) in Thailand. J. Fungi 2022, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, K.; Togashi, K.; Arie, T.; Teraoka, T. A simple method for a mini-preparation of fungal DNA. J. Gen. Plant Pathol. 2006, 72, 348–350. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.H.; Taylor, J.W. PCR Protocols: A Guide to Methods and Application; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MRBAYES 3.2: Efficient Bayesian phylogenetic inference and model selection across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Lombard, L.; Chen, S.F.; Mou, X.; Zhou, X.D.; Crous, P.W.; Wingfield, M.J. New species, hyper-diversity and potential importance of Calonectria spp. from Eucalyptus in South China. Stud. Mycol. 2015, 80, 151–188. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Edwards, J.; Taylor, P.W.J.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Wingfield, M.J.; Alfenas, A.C.; Crous, P.W. The forgotten Calonectria collection: Pouring old wine into new bags. Stud. Mycol. 2016, 85, 159–198. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Sunpapao, A. Fusarium incarnatum is associated with postharvest fruit rot of muskmelon (Cucumis melo). J. Phytopathol. 2020, 168, 204–210. [Google Scholar] [CrossRef]
- Shirata, A.; Tomiyama, K.; Doke, N.; Takahashi, K. Increases in peroxidase- and polyphenoloxidase- activities and production of antifungal substances in mulberry shoots following wounding or infection by pathogenic fungi. J.-Phytopathol. Soc. Japan 1978, 44, 127–136. [Google Scholar] [CrossRef][Green Version]
- Pornsuriya, C.; Ito, S.; Sunpapao, A. First report of leaf spot on lettuce caused by Curvularia aeria. J. Gen. Plant Pathol. 2018, 84, 296–299. [Google Scholar] [CrossRef]
- Bose, R.; Pandey, S.; Joshi, P.; Banerjee, S.; Pandey, A.; Bhandari, M.S. First report of Calonectria cerciana causing leaf blight of Eucalyptus in northern India. Forest Pathol. 2020, 54, e12658. [Google Scholar] [CrossRef]
- Stępniewska, H.; Jankowiak, R.; Boroń, P.; Woźniak, K. First report of Calonectria montana causing damping-off disease on pine and spruce seedlings in Europe. Forest Pathol. 2020, 50, e12595. [Google Scholar] [CrossRef]
- Crous, P.W. Taxonomy and Pathology of Cylindrocladium (Calonectria) and Allied Genera; The American Phytopathological Society: St. Paul, MN, USA, 2002. [Google Scholar]
- Alfenas, R.F.; Lombard, L.; Pereira, O.L.; Alfenas, A.C.; Crous, P.W. Diversity and potential impact of Calonectria species in Eucalyptus plantations in Brazil. Stud. Mycol. 2015, 80, 89–130. [Google Scholar] [CrossRef]
- Lombard, L.; Rodas, C.A.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Calonectria (Cylindrocladium) species associated with dying Pinus cuttings. Persoonia 2009, 23, 41–47. [Google Scholar] [CrossRef]
- Wu, W.; Chen, S. Species diversity, mating strategy and pathogenicity of Calonectria species from diseased leaves and soils in the Eucalyptus plantation in southern China. J. Fungi 2021, 7, 73. [Google Scholar] [CrossRef]
- Lombard, L.; Crous, P.W.; Wingfield, D.B.; Wingfield, M.J. Species concepts in Calonectria (Cylindrocladium). Stud. Mycol. 2010, 66, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zheng, L.; Wang, C.; Xie, C.P. The first report of Calonectria pteridis causing a leaf spot disease on Serenoa repens in China. Plant Dis. 2014, 98, 854. [Google Scholar] [CrossRef] [PubMed]
- Soares, I.D.; Auer, C.G.; dos Santos, A.F.; da Silva, R.A.F.; Soares, T.P.F.; Ferreira, M.A. First report of Calonectria leaf blight caused by Calonectria metrosideri on Eucalyptus benthamii in Brazil. Plant Dis. 2019, 103, 2477. [Google Scholar] [CrossRef]
- Aiello, D.; Fiorenza, A.; Gusella, G.; Polizzi, G. First report of Calonectria tunisiana causing crown and root rot on Eucalyptus globulus. J. Plant Pathol. 2020, 102, 1353. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calonectria Isolates | 3 dpi. 1 | 5 dpi. | ||
|---|---|---|---|---|
| Unwounded 2 | Wounded | Unwounded | Wounded | |
| Calkb001 | - | 24.8 ± 2.3 | 17.0 ± 9.8 | 54.3 ± 1.5 |
| Calkb002 | - | 24.3 ± 0.3 | - | 57.5 ± 0.3 |
| Calkb003 | - | 21.5 ± 1.2 | 11.0 ± 8.5 | 51.0 ± 2.3 |
| Calkb004 | - | 23.5 ± 1.3 | - | 56.3 ± 3.0 |
| ns 3 | ns | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thaochan, N.; Pornsuriya, C.; Chairin, T.; Chomnunti, P.; Sunpapao, A. Morphological and Molecular Characterization of Calonectria foliicola Associated with Leaf Blight on Rubber Tree (Hevea brasiliensis) in Thailand. J. Fungi 2022, 8, 986. https://doi.org/10.3390/jof8100986
Thaochan N, Pornsuriya C, Chairin T, Chomnunti P, Sunpapao A. Morphological and Molecular Characterization of Calonectria foliicola Associated with Leaf Blight on Rubber Tree (Hevea brasiliensis) in Thailand. Journal of Fungi. 2022; 8(10):986. https://doi.org/10.3390/jof8100986
Chicago/Turabian StyleThaochan, Narit, Chaninun Pornsuriya, Thanunchanok Chairin, Putarak Chomnunti, and Anurag Sunpapao. 2022. "Morphological and Molecular Characterization of Calonectria foliicola Associated with Leaf Blight on Rubber Tree (Hevea brasiliensis) in Thailand" Journal of Fungi 8, no. 10: 986. https://doi.org/10.3390/jof8100986
APA StyleThaochan, N., Pornsuriya, C., Chairin, T., Chomnunti, P., & Sunpapao, A. (2022). Morphological and Molecular Characterization of Calonectria foliicola Associated with Leaf Blight on Rubber Tree (Hevea brasiliensis) in Thailand. Journal of Fungi, 8(10), 986. https://doi.org/10.3390/jof8100986