
Effects of Five Filamentous Fungi Used in Food Processes on In Vitro and In Vivo Gut Inflammation

 , ,
, ,
Abstract
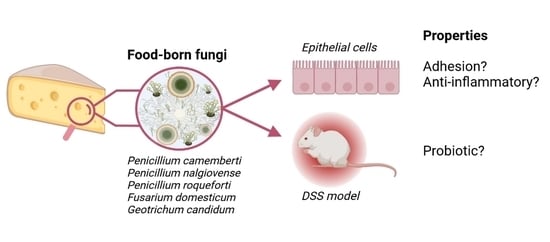
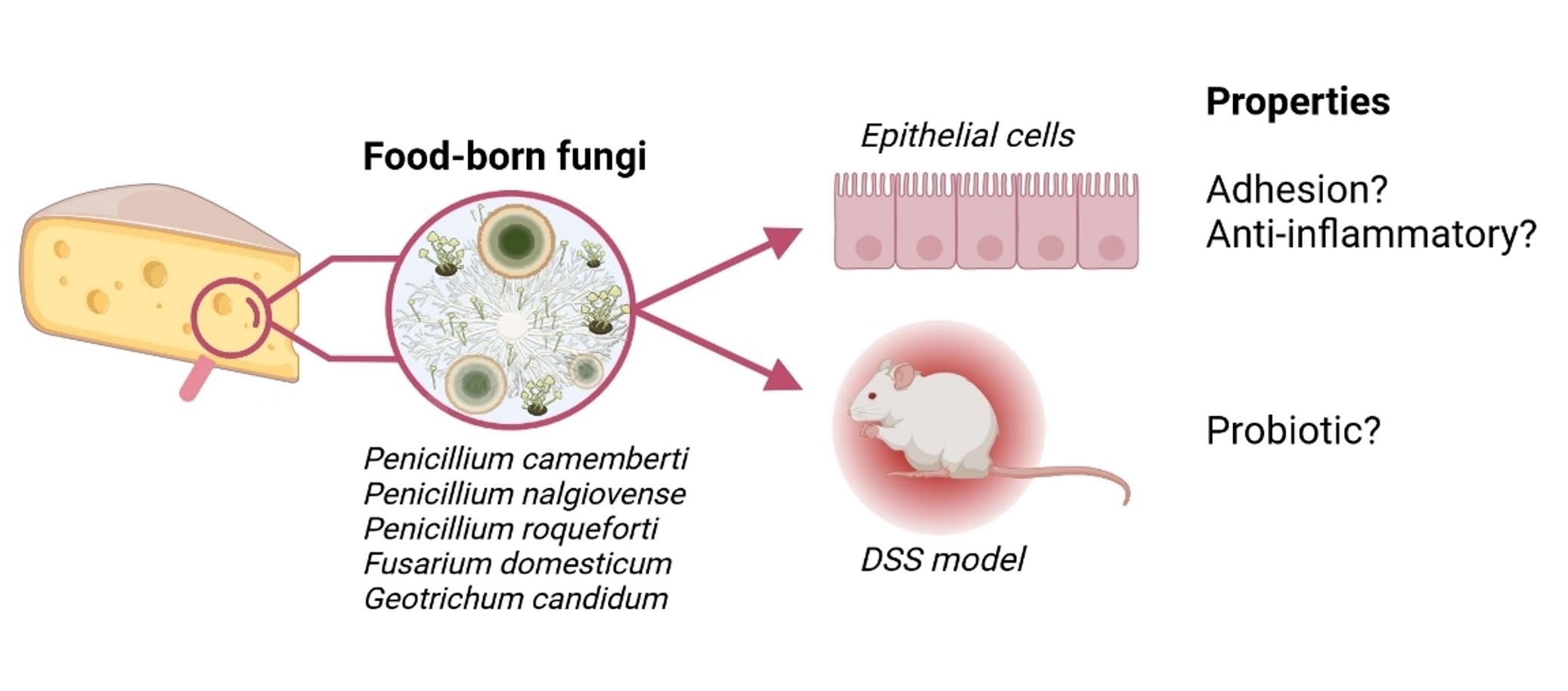
:
1. Introduction
2. Materials and Methods
2.1. Cells Culture
2.2. Preparation of Fungi
2.3. Fungal Adhesion on Caco-2 and HT29-MTX cells
2.3.1. Quantitative Assessment of the Adhesion
2.3.2. DNA Extraction
2.4. IL-8 Production by HT29 after Co-Culture with Fungi
2.5. Human Peripheral Blood Mononuclear Cells (PBMCs) and Fungi Co-Incubation
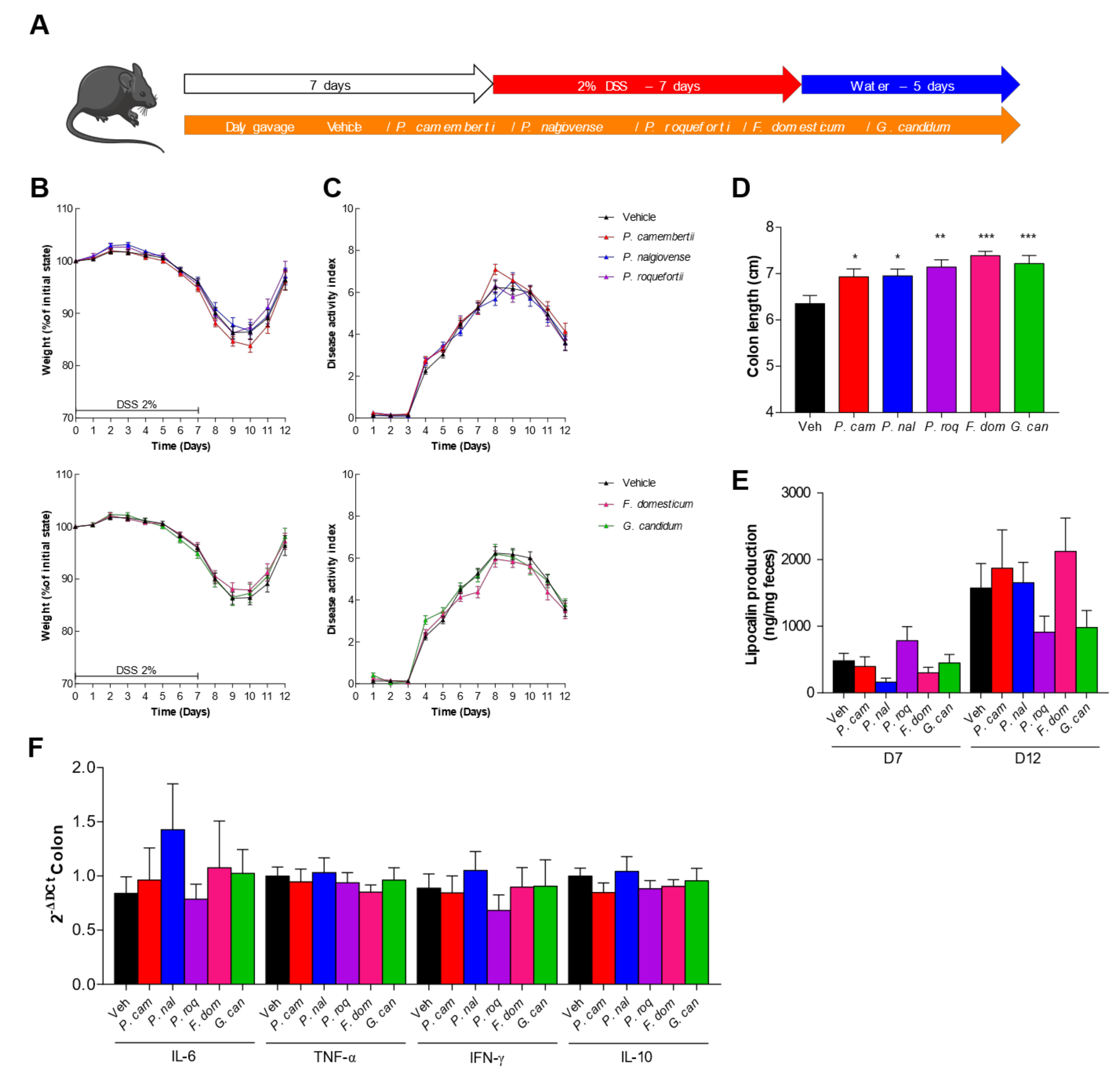
2.6. Colitis Model in the Mice
2.7. Tissues and Samples
2.8. Quantification of Fecal Lipocalin (LCN2) Levels
2.9. RNA Extraction and Gene Expression Analysis Using Quantitative Real-Time PCR (qRT–PCR)
2.10. Statistical Analysis
3. Results
3.1. Adhesion Properties Strongly Vary between Filamentous Strains
3.2. G. candidum Spores Elicite Low Inflammation on HT29 Epithelial Cells
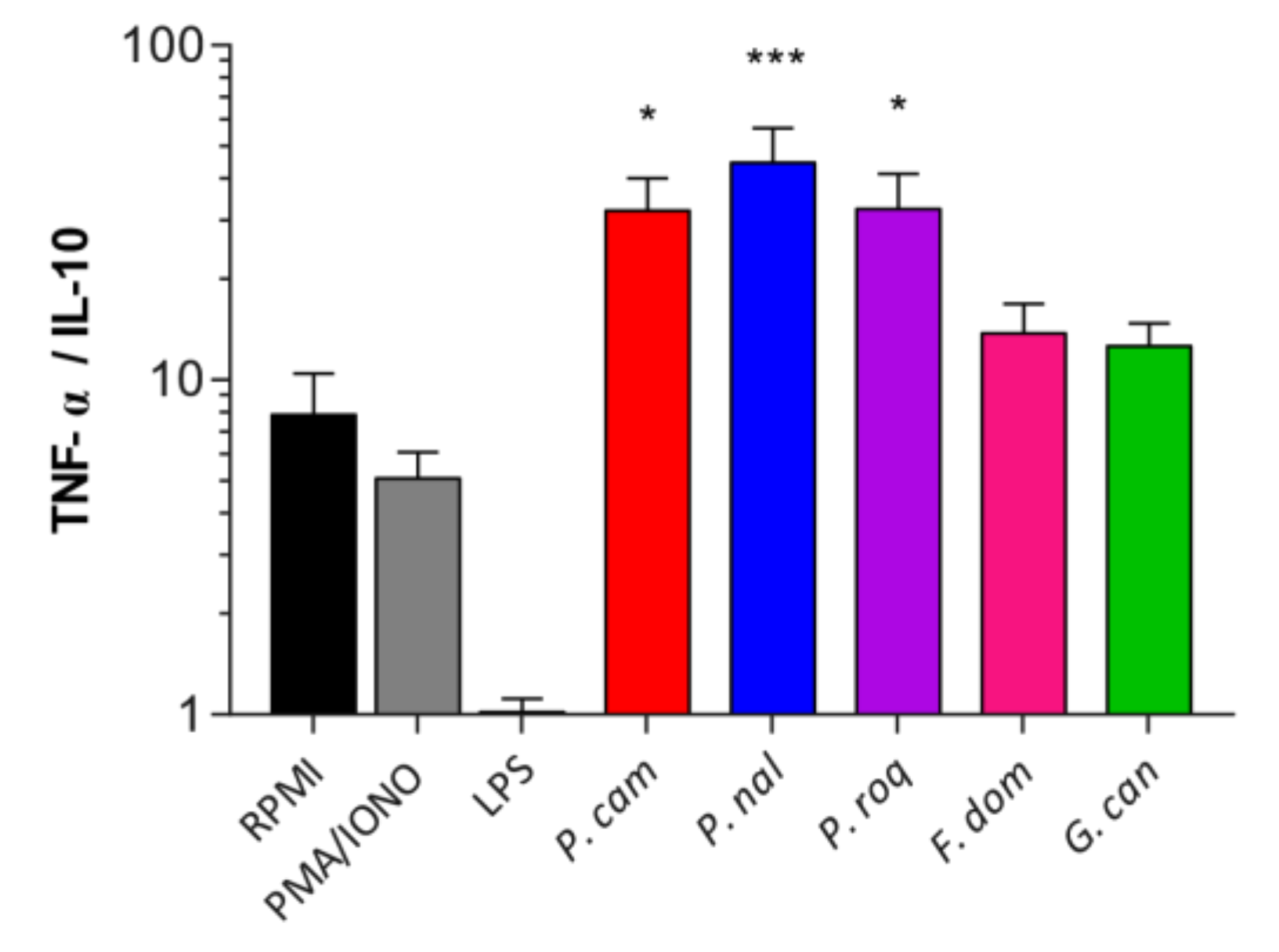
3.3. Human Immune Cells Differentially Recognize Fungal Spores
3.4. DSS-Induced Colitis Is Not Influenced by Fungal Spores’ Oral Administration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, W.; Lv, X.; Tran, V.-T.; Maruyama, J.; Han, K.-H.; Yu, J.-H. Editorial: From Traditional to Modern: Progress of Molds and Yeasts in Fermented-Food Production. Front. Microbiol. 2022, 13, 876872. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.K.; Pandey, A.; Sahoo, D. Biotechnological Potential of Yeasts in Functional Food Industry. Trends Food Sci. Technol. 2019, 83, 129–137. [Google Scholar] [CrossRef]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An Emerging Yeast Cell Factory for Applications in Food and Biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef]
- Khachatourians, G. Fungi in Food Technology; University of Saskatchewan: Saskatoon, SK, Canada, 2003. [Google Scholar] [CrossRef]
- McFarland, L.V. Systematic Review and Meta-Analysis of Saccharomyces boulardii in Adult Patients. World J. Gastroenterol. 2010, 16, 2202–2222. [Google Scholar] [CrossRef]
- Zanello, G.; Berri, M.; Dupont, J.; Sizaret, P.Y.; D’Inca, R.; Salmon, H.; Meurens, F. Saccharomyces cerevisiae Modulates Immune Gene Expressions and Inhibits ETEC-Mediated ERK1/2 and P38 Signaling Pathways in Intestinal Epithelial Cells. PLoS ONE 2011, 6, e18573. [Google Scholar] [CrossRef] [PubMed]
- Pineton de Chambrun, G.; Neut, C.; Chau, A.; Cazaubiel, M.; Pelerin, F.; Justen, P.; Desreumaux, P. A Randomized Clinical Trial of Saccharomyces cerevisiae versus Placebo in the Irritable Bowel Syndrome. Dig. Liver Dis. 2015, 47, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Sugiharto, S. A Review of Filamentous Fungi in Broiler Production. Ann. Agric. Sci. 2019, 64, 1–8. [Google Scholar] [CrossRef]
- Melo-Bolívar, J.F.; Ruiz-Pardo, R.Y.; Hume, M.E.; Sidjabat, H.E.; Villamil-Diaz, L.M. Probiotics for Cultured Freshwater Fish. Microbiol. Aust. 2020, 41, 105–108. [Google Scholar] [CrossRef]
- Sugiharto, S.; Yudiarti, T.; Isroli, I. Functional Properties of Filamentous Fungi Isolated from the Indonesian Fermented Dried Cassava, with Particular Application on Poultry. Mycobiology 2015, 43, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Karimi, S.; Soofiani, N.M.; Lundh, T.; Mahboubi, A.; Kiessling, A.; Taherzadeh, M.J. Evaluation of Filamentous Fungal Biomass Cultivated on Vinasse as an Alternative Nutrient Source of Fish Feed: Protein, Lipid, and Mineral Composition. Fermentation 2019, 5, 99. [Google Scholar] [CrossRef] [Green Version]
- Amir, I.; Zuberi, A.; Kamran, M.; Imran, M. Evaluation of Commercial Application of Dietary Encapsulated Probiotic (Geotrichum candidum QAUGC01): Effect on Growth and Immunological Indices of Rohu (Labeo Rohita, Hamilton 1822) in Semi-Intensive Culture System. Fish Shellfish Immunol. 2019, 95, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Noor-Ul, H.; Haokun, L.; Junyan, J.; Xiaoming, Z.; Dong, H.; Yunxia, Y.; Shouqi, X. Dietary Supplementation of Geotrichum candidum Improves Growth, Gut Microbiota, Immune-Related Gene Expression and Disease Resistance in Gibel Carp CAS Ⅲ (Carassius Auratus Gibelio). Fish Shellfish Immunol. 2020, 99, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Fierro, F.; Laich, F.; García-Rico, R.O.; Martín, J.F. High Efficiency Transformation of Penicillium nalgiovense with Integrative and Autonomously Replicating Plasmids. Int. J. Food Microbiol. 2004, 90, 237–248. [Google Scholar] [CrossRef]
- Dupont, J.; Dequin, S.; Giraud, T.; Le Tacon, F.; Marsit, S.; Ropars, J.; Richard, F.; Selosse, M.A. Fungi as a Source of Food. Microbiol. Spectr. 2017, 5, 1063–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, M.; Andrade, M.J.; García, C.; Rondán, J.J.; Núñez, F. Effects of Preservative Agents on Quality Attributes of Dry-Cured Fermented Sausages. Foods 2020, 9, 1505. [Google Scholar] [CrossRef] [PubMed]
- Querol, A.; Barrio, E.; Ramón, D. A Comparative Study of Different Methods of Yeast Strain Characterization. Syst. Appl. Microbiol. 1992, 15, 439–446. [Google Scholar] [CrossRef]
- Chassaing, B.; Srinivasan, G.; Delgado, M.A.; Young, A.N.; Gewirtz, A.T.; Vijay-Kumar, M. Fecal Lipocalin 2, a Sensitive and Broadly Dynamic Non-Invasive Biomarker for Intestinal Inflammation. PLoS ONE 2012, 7, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Meneses, A.M.C.; Schneeberger, K.; Kruitwagen, H.S.; Penning, L.C.; van Steenbeek, F.G.; Burgener, I.A.; Spee, B. Intestinal Organoids-Current and Future Applications. Vet. Sci. 2016, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, M.; Zihler Berner, A.; Chervet, N.; Chassard, C.; Lacroix, C. Comparison of the Caco-2, HT-29 and the Mucus-Secreting HT29-MTX Intestinal Cell Models to Investigate Salmonella Adhesion and Invasion. J. Microbiol. Methods 2013, 94, 274–279. [Google Scholar] [CrossRef]
- Yoshimoto, A.N.; Bernardazzi, C.; Carneiro, A.J.V.; Elia, C.C.S.; Martinusso, C.A.; Ventura, G.M.; Castelo-Branco, M.T.L.; de Souza, H.S.P. Hedgehog Pathway Signaling Regulates Human Colon Carcinoma HT-29 Epithelial Cell Line Apoptosis and Cytokine Secretion. PLoS ONE 2012, 7, e45332. [Google Scholar] [CrossRef] [Green Version]
- Lammers, K.M.; Helwig, U.; Swennen, E.; Rizzello, F.; Venturi, A.; Caramelli, E.; Kamm, M.A.; Brigidi, P.; Gionchetti, P.; Campieri, M. Effect of Probiotic Strains on Interleukin 8 Production by HT29/19A Cells. Am. J. Gastroenterol. 2002, 97, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, K.E.; Cutilli, J.; Piliero, L.M.; Ghavimi-Alagha, D.; Starr, S.E.; Campbell, D.E.; Douglas, S.D. Measurement of Cytokine Secretion, Intracellular Protein Expression, and MRNA in Resting and Stimulated Peripheral Blood Mononuclear Cells. Clin. Diagn. Lab. Immunol. 2000, 7, 920–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strong, P.J.; Self, R.; Allikian, K.; Szewczyk, E.; Speight, R.; O’Hara, I.; Harrison, M.D. Filamentous Fungi for Future Functional Food and Feed. Curr. Opin. Biotechnol. 2022, 76, 102729. [Google Scholar] [CrossRef] [PubMed]
- Venturini Copetti, M. Yeasts and Molds in Fermented Food Production: An Ancient Bioprocess. Curr. Opin. Food Sci. 2019, 25, 57–61. [Google Scholar] [CrossRef]
- Richard, M.L.; Sokol, H. The Gut Mycobiota: Insights into Analysis, Environmental Interactions and Role in Gastrointestinal Diseases. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 331–345. [Google Scholar] [CrossRef]
- Marcellino, N.; Beuvier, E.; Grappin, R.; Guéguen, M.; Benson, D.R. Diversity of Geotrichum candidum Strains Isolated from Traditional Cheesemaking Fabrications in France. Appl. Environ. Microbiol. 2001, 67, 4752–4759. [Google Scholar] [CrossRef] [Green Version]
- Ropars, J.; Cruaud, C.; Lacoste, S.; Dupont, J. A Taxonomic and Ecological Overview of Cheese Fungi. Int. J. Food Microbiol. 2012, 155, 199–210. [Google Scholar] [CrossRef]
- Ropars, J.; Didiot, E.; Rodríguez de la Vega, R.C.; Bennetot, B.; Coton, M.; Poirier, E.; Coton, E.; Snirc, A.; Le Prieur, S.; Giraud, T. Domestication of the Emblematic White Cheese-Making Fungus Penicillium camemberti and Its Diversification into Two Varieties. Curr. Biol. 2020, 30, 4441–4453.e4. [Google Scholar] [CrossRef]
- Coton, E.; Coton, M.; Hymery, N.; Mounier, J.; Jany, J.L. Penicillium Roqueforti: An Overview of Its Genetics, Physiology, Metabolism and Biotechnological Applications. Fungal Biol. Rev. 2020, 34, 59–73. [Google Scholar] [CrossRef]
- Perkins, V.; Vignola, S.; Lessard, M.H.; Plante, P.L.; Corbeil, J.; Dugat-Bony, E.; Frenette, M.; Labrie, S. Phenotypic and Genetic Characterization of the Cheese Ripening Yeast Geotrichum candidum. Front. Microbiol. 2020, 11, 737. [Google Scholar] [CrossRef]
- Metin, B. Filamentous Fungi in Cheese Production; IGI Global: Hershey, PA, USA, 2018; pp. 257–275. [Google Scholar] [CrossRef] [Green Version]
- Bernáldez, V.; Córdoba, J.J.; Rodríguez, M.; Cordero, M.; Polo, L.; Rodríguez, A. Effect of Penicillium nalgiovense as Protective Culture in Processing of Dry-Fermented Sausage “Salchichón”. Food Control. 2013, 32, 69–76. [Google Scholar] [CrossRef]
- Ludemann, V.; Greco, M.; Rodríguez, M.P.; Basílico, J.C.; Pardo, A.G. Conidial Production by Penicillium nalgiovense for Use as Starter Cultures in Dry Fermented Sausages by Solid State Fermentation. LWT—Food Sci. Technol. 2010, 43, 315–318. [Google Scholar] [CrossRef]
- Ryu, S.; Park, M.R.; Maburutse, B.E.; Lee, W.J.; Park, D.J.; Cho, S.; Hwang, I.; Oh, S.; Kim, Y. Diversity and Characteristics of the Meat Microbiological Community on Dry Aged Beef. J. Microbiol. Biotechnol. 2018, 28, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.E.; Lee, H.J.; Kim, C.H.; Ryu, S.; Kim, Y.; Jo, C. Effect of Penicillium candidum and Penicillium calgiovense and Their Combination on the Physicochemical and Sensory Quality of Dry-Aged Beef. Food Microbiol. 2022, 107, 104083. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Yudiarti, T.; Isroli, I. Assay of Antioxidant Potential of Two Filamentous Fungi Isolated from the Indonesian Fermented Dried Cassava. Antioxidants 2016, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Yun, H.M.; Kim, I.H. The Effect of Feeding Rhizopus oligosporus on Growth Performance, Nutrient Digestibility, Blood Profile, Fecal Microbiota, and Fecal Score in Weanling Pigs. Turk. J. Vet. Anim. Sci. 2016, 40, 700–706. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | 5′–Forward–3′ | 5′–Reverse–3′ |
|---|---|---|
| GAPDH | AACTTTGGCATTGTGGAAGG | ACACATTGGGGGTAGGAACA |
| IL-10 | AGAAGCATGGCCCAGAAATCA | GGCCTTGTAGACACCTTGGT |
| IL-6 | GTAGCTATGGTACTCCAGAAGAC | ACGATGATGCACTTGCAGAA |
| INF-γ | CCATCCTTTTGCCAGTTCCTC | ATGAACGCTACACACTGCATC |
| TNF-α | GACCCTCACACTCAGATCATCTTCT | CCACTTGGTGGTTTGCTACGA |
| TEF1a | GATTTCATCAAGAACATGAT | GACGTTGAAACCGACGTTGTC |
| Score | Consistency Stool | Blood |
|---|---|---|
| 0 | Normal | Negative (−) |
| 2 | Loose stools | + |
| 4 | Diarrhea | Bleeding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poirier, M.; Hugot, C.; Spatz, M.; Da Costa, G.; Lapiere, A.; Michaudel, C.; Danne, C.; Martin, V.; Langella, P.; Michel, M.-L.; et al. Effects of Five Filamentous Fungi Used in Food Processes on In Vitro and In Vivo Gut Inflammation. J. Fungi 2022, 8, 893. https://doi.org/10.3390/jof8090893
Poirier M, Hugot C, Spatz M, Da Costa G, Lapiere A, Michaudel C, Danne C, Martin V, Langella P, Michel M-L, et al. Effects of Five Filamentous Fungi Used in Food Processes on In Vitro and In Vivo Gut Inflammation. Journal of Fungi. 2022; 8(9):893. https://doi.org/10.3390/jof8090893
Chicago/Turabian StylePoirier, Maxime, Cindy Hugot, Madeleine Spatz, Gregory Da Costa, Alexia Lapiere, Chloé Michaudel, Camille Danne, Valérie Martin, Philippe Langella, Marie-Laure Michel, and et al. 2022. "Effects of Five Filamentous Fungi Used in Food Processes on In Vitro and In Vivo Gut Inflammation" Journal of Fungi 8, no. 9: 893. https://doi.org/10.3390/jof8090893
APA StylePoirier, M., Hugot, C., Spatz, M., Da Costa, G., Lapiere, A., Michaudel, C., Danne, C., Martin, V., Langella, P., Michel, M.-L., Sokol, H., Boyaval, P., & Richard, M. L. (2022). Effects of Five Filamentous Fungi Used in Food Processes on In Vitro and In Vivo Gut Inflammation. Journal of Fungi, 8(9), 893. https://doi.org/10.3390/jof8090893