
Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records


Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Voucher | Locality | GenBank Accession No. | Reference | ||
|---|---|---|---|---|---|---|
| LSU | tef1 | rpb2 | ||||
| Cantharellus afrocibarius | BB 96.236 | Zambia | KF294669 | JX192994 | KF294747 | [3] |
| C. afrocibarius | BB 96.235 (HT) | Zambia | KF294668 | JX192993 | KF294746 | [3] |
| C. albovenosus | 1690 (HT) | South Korean | – | KY271942 | – | [11] |
| C. albovenosus | 1713 | South Korean | – | MW124387 | – | [11] |
| C. albovenosus | GDGM85853 | China | OM978952 | ON119062 | ON119006 | Present study |
| C. albovenosus | GDGM85846 | China | OM978950 | ON119060 | ON119004 | Present study |
| C. albovenosus | GDGM85142 | China | OM978949 | ON119059 | ON229082 | Present study |
| C. albovenosus | HMAS279296 | China | OM978948 | ON119066 | ON119010 | Present study |
| C. albovenosus | HMAS279284 | China | ON212414 | ON119064 | ON119008 | Present study |
| C. albovenosus | HMAS279292 | China | ON212412 | ON119065 | ON119009 | Present study |
| C. albovenosus | HMAS279262 | China | OM978947 | ON119063 | ON119007 | Present study |
| C. albovenosus | GDGM85852 | China | OM978951 | ON119061 | ON119005 | Present study |
| C. albus | HKAS107045 (HT) | China | MT782540 | MT776015 | MT776012 | [12] |
| C. albus | GDGM81399 | China | MZ605074 | MZ613977 | MZ614022 | [13] |
| C. albus | GDGM81064 | China | MZ605073 | MZ613976 | MZ614021 | [13] |
| C. appalachiensis | GRSM77088 | USA | DQ898690 | – | DQ898748 | [31] |
| C. appalachiensis | BB 07.123 | USA | KF294635 | GQ914979 | KF294711 | [3] |
| C. aurantinus | GDGM46278 (HT) | China | MZ766517 | MZ766560 | [13] | |
| C. aurantinus | GDGM46279 | China | MZ766518 | MZ766561 | MZ766571 | [13] |
| C. aurantinus | GDGM81899 | China | MZ766520 | MZ766563 | MZ766573 | [13] |
| C. aurantinus | GDGM84974 | China | MZ766521 | MZ766564 | MZ766572 | [13] |
| C. austrosinensis | GDGM81303 | China | MZ605084 | MZ613986 | MZ614029 | [13] |
| C. austrosinensis | GDGM81249 (HT) | China | MZ605082 | MZ613983 | MZ614027 | [13] |
| C. austrosinensis | GDGM80616 | China | MZ605081 | MZ613982 | MZ614026 | [13] |
| C. austrosinensis | GDGM81381 | China | MZ605086 | MZ613988 | MZ614031 | [13] |
| C. austrosinensis | GDGM81379 | China | MZ605085 | MZ613987 | MZ614030 | [13] |
| C. austrosinensis | GDGM81985 | China | MZ605087 | MZ613989 | MZ614032 | [13] |
| C. chrysanthus | GDGM45166 | China | OM978959 | ON119074 | ON119011 | Present study |
| C. chrysanthus | GDGM45937 | China | OM978960 | ON119075 | ON119012 | Present study |
| C. chrysanthus | GDGM85298 | China | OM978975 | ON119089 | ON119025 | Present study |
| C. chrysanthus | GDGM85305 | China | OM978976 | ON119090 | ON119026 | Present study |
| C. chrysanthus | GDGM53485 | China | OM978962 | ON119077 | ON119014 | Present study |
| C. chrysanthus | GDGM80220 (HT) | China | OM978970 | ON119083 | ON119019 | Present study |
| C. chrysanthus | GDGM82511 | China | OM978973 | ON119087 | ON119023 | Present study |
| C. chrysanthus | GDGM82516 | China | OM978974 | ON119088 | ON119024 | Present study |
| C. chrysanthus | GDGM80436 | China | OM978971 | ON119084 | ON119020 | Present study |
| C. chrysanthus | GDGM80202 | China | OM978965 | ON119080 | ON119016 | Present study |
| C. chrysanthus | GDGM80204 | China | OM978966 | ON119081 | ON119017 | Present study |
| C. chrysanthus | HMAS279434 | China | ON212413 | ON119091 | ON229079 | Present study |
| C. chrysanthus | GDGM80438 | China | – | ON119085 | ON119021 | Present study |
| C. chrysanthus | GDGM82473 | China | OM978972 | ON119086 | ON119022 | Present study |
| C. chrysanthus | GDGM77035 | China | OM978964 | ON119079 | ON229081 | Present study |
| C. chrysanthus | GDGM60524 | China | OM978963 | ON119078 | ON119015 | Present study |
| C. chrysanthus | GDGM80217 | China | OM978969 | ON119082 | ON119018 | Present study |
| C. chrysanthus | GDGM49628 | China | OM978961 | ON119076 | ON119013 | Present study |
| C. chrysanthus | GDGM87950 | China | OM978968 | – | ON119027 | Present study |
| C. chrysanthus | GDGM87951 | China | OM978967 | – | ON119028 | Present study |
| C. cibarius | GE 07.025 | France | KF294658 | GQ914949 | KF294736 | [3] |
| C. cibarius | BB 07.300 | Slovakia | KF294641 | GQ914950 | KF294718 | [3] |
| C. cinnabarinus | BB 04.263 (NT) | USA | – | GQ914983 | – | [32] |
| C. cinnabarinus | BB 07.053 | USA | KF294630 | GQ914984 | KF294705 | [32] |
| C. cinnabarinus | BB 07.001 | USA | KF294624 | GQ914985 | KF294698 | [32] |
| C. citrinus | 1691 (HT) | South Korean | – | MW124385 | – | [16] |
| C. citrinus | 1715 | South Korean | – | MW124388 | – | [16] |
| C. citrinus | 1710 | South Korean | – | MW124386 | – | [16] |
| C. citrinus | 1711 | South Korean | – | MW124384 | – | [16] |
| C. citrinus | GDGM86140 | China | OM978955 | ON119070 | ON119032 | Present study |
| C. citrinus | GDGM86141 | China | OM978956 | ON119071 | ON119033 | Present study |
| C. citrinus | GDGM80825 | China | – | ON119069 | ON119031 | Present study |
| C. citrinus | GDGM86142 | China | OM978957 | ON119072 | ON119034 | Present study |
| C. citrinus | GDGM80724 | China | OM978954 | ON119068 | ON119030 | Present study |
| C. citrinus | GDGM86143 | China | OM978958 | ON119073 | ON119035 | Present study |
| C. citrinus | GDGM80723 | China | OM978953 | ON119067 | ON119029 | Present study |
| C. coccolobae | 1064_RC. 14_24 | Guadeloupe | KX857088 | KX857020 | KX856992 | [33] |
| C. coccolobae | 1065_RC. 11_25 (HT) | Guadeloupe | KX857089 | KX857021 | KX856993 | [33] |
| C. congolensis | 1645/BB16.044 | Saharan Africa | KX857102 | KX857075 | KX857006 | [33] |
| C. congolensis | 1676/BB16.123 | Saharan Africa | KX857106 | KX857078 | KX857010 | [33] |
| C. aff. congolensis | BB 06.176 | Madagascar | KF294606 | – | KF294680 | [3] |
| C. aff. congolensis | BB 06.197 | Madagascar | KF294608 | – | KF294683 | [3] |
| C. convexus | GDGM54841 | China | OM978940 | ON119052 | ON119036 | Present study |
| C. convexus | GDGM70307 (HT) | China | OM978941 | ON119053 | ON119037 | Present study |
| C. corallinus | 1083_JJ_MO_CANT_2 | USA | – | KX857031 | – | [34] |
| C. corallinus | 1086_JJ_MO_CANT_5 | USA | – | KX857034 | – | [34] |
| C. corallinus | FLAS_F_61106 | USA | – | MK045368 | – | [34] |
| C. curvatus | BRNM:825749 (HT) | South Korea | MW124390 | [16] | ||
| C. cyphelloides | TNS F-61721 (HT) | Japan | NG059027 | – | – | [35] |
| C. decolorans | BB 08.278 (HT) | Madagascar | KF294654 | GQ914968 | KF294731 | [3] |
| C. fistulosus | DT_43 | Tanzania | JQ976965 | JX192997 | – | [3] |
| C. friesii | AH44798 | Spain | KR677522 | KX828831 | KX828752 | [36] |
| C. friesii | VDKO 1165 | Africa | – | KX834408 | KX881922 | [5] |
| C. galbanus | GDGM86249 (HT) | China | ZM766516 | MZ766568 | MZ766577 | [13] |
| C. garnierii | BB 09.024 | New Caledonia | KX857085 | KX857017 | KX856989 | [34] |
| C. garnierii | BB 09.283 | New Caledonia | KX857087 | KX857019 | KX856991 | [34] |
| C. garnierii | BB 09.033 | New Caledonia | KX857086 | KX857018 | KX856990 | [34] |
| C. garnierii | RF33 | New Caledonia | AY392768 | – | [37] | |
| C. garnierii | RF32 | New Caledonia | AY392767 | – | [37] | |
| C. koreanus | 1697 | South Korea | – | KY271940 | – | [11] |
| C. koreanus | 1689 (HT) | South Korea | – | KY271941 | – | [11] |
| C. koreanus | GDGM85306 | China | OM978978 | ON119093 | ON229077 | Present study |
| C. koreanus | GDGM79233 | China | OM978977 | ON119092 | ON229078 | Present study |
| C. koreanus | 1693 | South Korea | – | – | Unpublished | |
| C. koreanus | 1694 | South Korea | – | – | Unpublished | |
| C. koreanus | 1696 | South Korea | – | – | Unpublished | |
| C. luteolus | GDGM60393 (HT) | China | ZM766515 | MZ766566 | MZ766575 | [13] |
| C. luteolus | GDGM86247 | China | MZ766513 | MZ766567 | MZ766576 | [13] |
| C. luteolus | GDGM44258 | China | ZM766514 | MZ766566 | MZ766570 | [13] |
| C. luteovirens | GDGM81079 | China | MZ605092 | MZ613994 | MZ614036 | [13] |
| C. luteovirens | GDGM80672 (HT) | China | MZ605090 | MZ613992 | MZ614035 | [13] |
| C. luteovirens | GDGM80680 | China | MZ605091 | MZ613993 | – | [13] |
| C. minioalbus | GDGM78910 | China | MZ605098 | MZ613999 | MZ614043 | [13] |
| C. minioalbus | GDGM78901 (HT) | China | MZ605097 | MZ613998 | MZ614042 | [13] |
| C. minioalbus | GDGM78916 | China | MZ605100 | MZ614001 | MZ614045 | [13] |
| C. minor | BB 07.057 | USA | KF294632 | JX192979 | KF294707 | [3] |
| C. minor | BB 07.002 | USA | KF294625 | JX192978 | KF294699 | [3] |
| C. neopersicinus | GDGM85145-1 | China | OM978942 | ON119054 | ON119039 | Present study |
| C. neopersicinus | GDGM85145-2 | China | OM978945 | ON119055 | ON119040 | Present study |
| C. neopersicinus | GDGM85145-3 | China | OM978946 | ON119056 | ON119041 | Present study |
| C. neopersicinus | GDGM87366-1 (HT) | China | OM978943 | ON119057 | ON119042 | Present study |
| C. neopersicinus | GDGM87366-2 | China | OM978944 | ON119058 | ON119043 | Present study |
| C. phloginus | GDGM79007-1 | China | OM978979 | ON119094 | ON119044 | Present study |
| C. phloginus | GDGM79007-2 | China | OM978980 | ON119095 | ON119045 | Present study |
| C. phloginus | SSC99 (HT) | China | – | KF801096 | – | [22] |
| C. phloginus | SSC98 | China | – | KF801095 | – | [22] |
| C. phloginus | Yuan14468 | China | – | MW999424. | – | [7] |
| C. phloginus | Yuan14490 | China | – | MW999425 | – | [7] |
| C. phloginus | GDGM82514 | China | – | ON119096 | – | Present study |
| C. pseudominimus | JV 00.663 | Portugal | KF294657 | JX192991 | KF294735 | [3,10] |
| C. romagnesianus | AH44218 | Spain | KX828807 | KX828836 | KX828757 | [36] |
| C. roseofagetorum | AH44789 | Georgia | KX828812 | KX828839 | KX828760 | [36] |
| C. sinocinnabarinus | GDGM83229 | China | OM978983 | ON119098 | ON119047 | Present study |
| C. sinocinnabarinus | GDGM83238 | China | OM978985 | ON119101 | ON119051 | Present study |
| C. sinocinnabarinus | GDGM83023 | China | OM978981 | ON119097 | ON119050 | Present study |
| C. sinocinnabarinus | GDGM83232 | China | – | ON119100 | ON119049 | Present study |
| C. sinocinnabarinus | GDGM83027 | China | OM978982 | – | ON119046 | Present study |
| C. sinocinnabarinus | GDGM83230 (HT) | China | OM978984 | ON119099 | ON119048 | Present study |
| C. sinocinnabarinus | HKAS58243 | China | JF906727 | – | – | [20] |
| C. sinominor | GDGM80788 | China | MZ605105 | MZ614004 | MZ614048 | [13] |
| C. sinominor | GDGM80842 (HT) | China | MZ605107 | MZ614006 | MZ614050 | [13] |
| C. sinominor | GDGM80885 | China | MZ605108 | MZ614007 | MZ614051 | [13] |
| C. aff. subcyanoxanthus | BB 98.014 | Tanzania | KF294615 | JX192973 | KF294689 | [3] |
| C. tabernensis | BB 07.119 | USA | KF294634 | GQ914976 | KF294709 | [3] |
| C. tabernensis | BB 07.056 (ET) | USA | KF294631 | GQ914974 | KF294706 | [3,38] |
| C. texensis | 341/O7.120 | USA | JN940601 | GQ914987 | KF294710 | [3] |
| C. texensis | BB 07.018 | USA | KF294626 | GQ914988 | KF294701 | [3] |
| C. xanthocyaneus | 1751 | Congo | MT006309 | MT002277 | – | [39] |
| C. xanthocyaneus | Congo | MT006310 | MT002278 | – | [39] | |
| C. zangii | GDGM82389 | China | MZ605110 | MZ614009 | MZ614053 | [13] |
| C. zangii | GDGM82393 | China | MZ605111 | MZ614010 | MZ614054 | [13] |
| C. zangii | GDGM82374 | China | MZ605109 | MZ614008 | MZ614052 | [13] |
3. Results
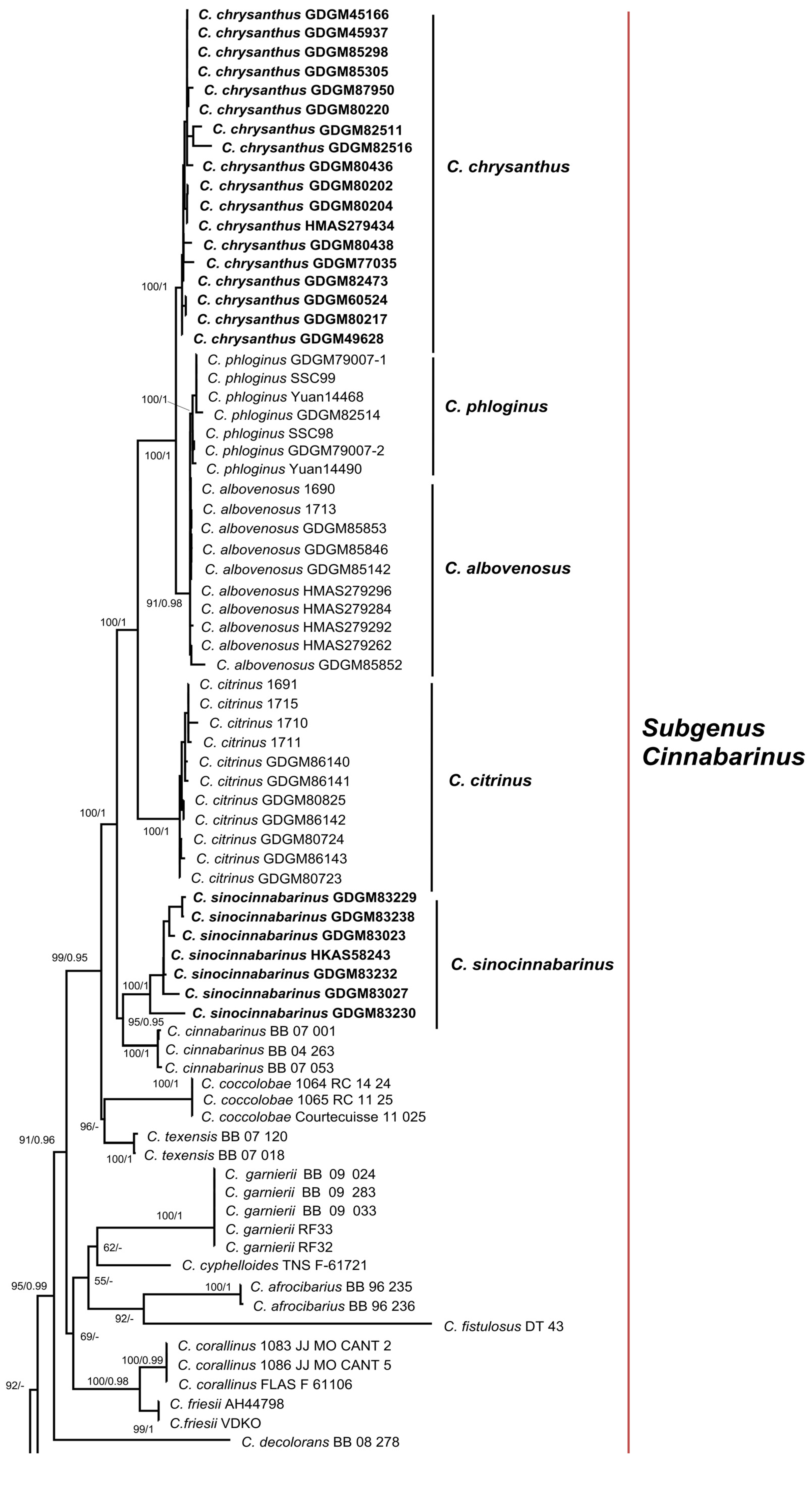
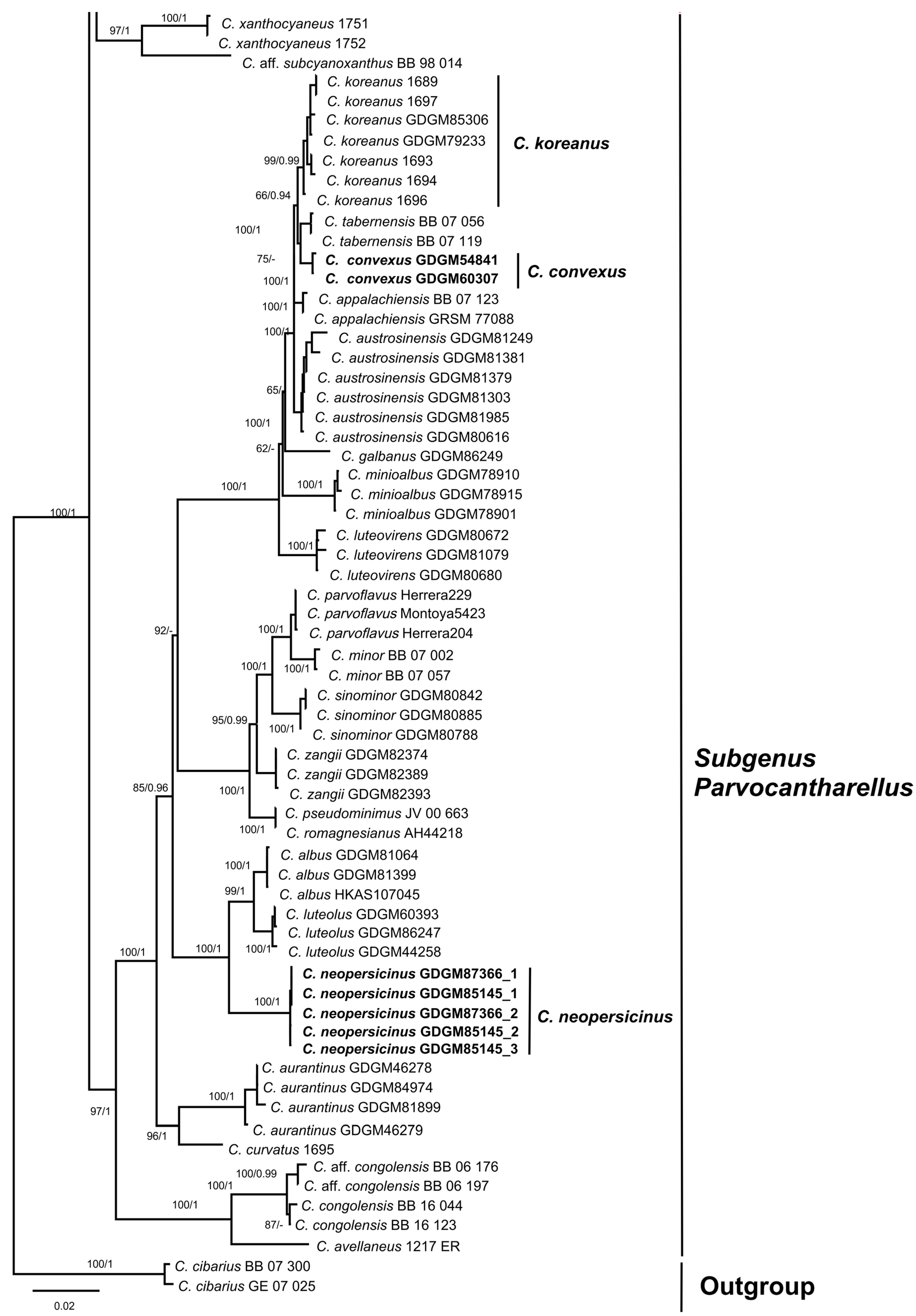
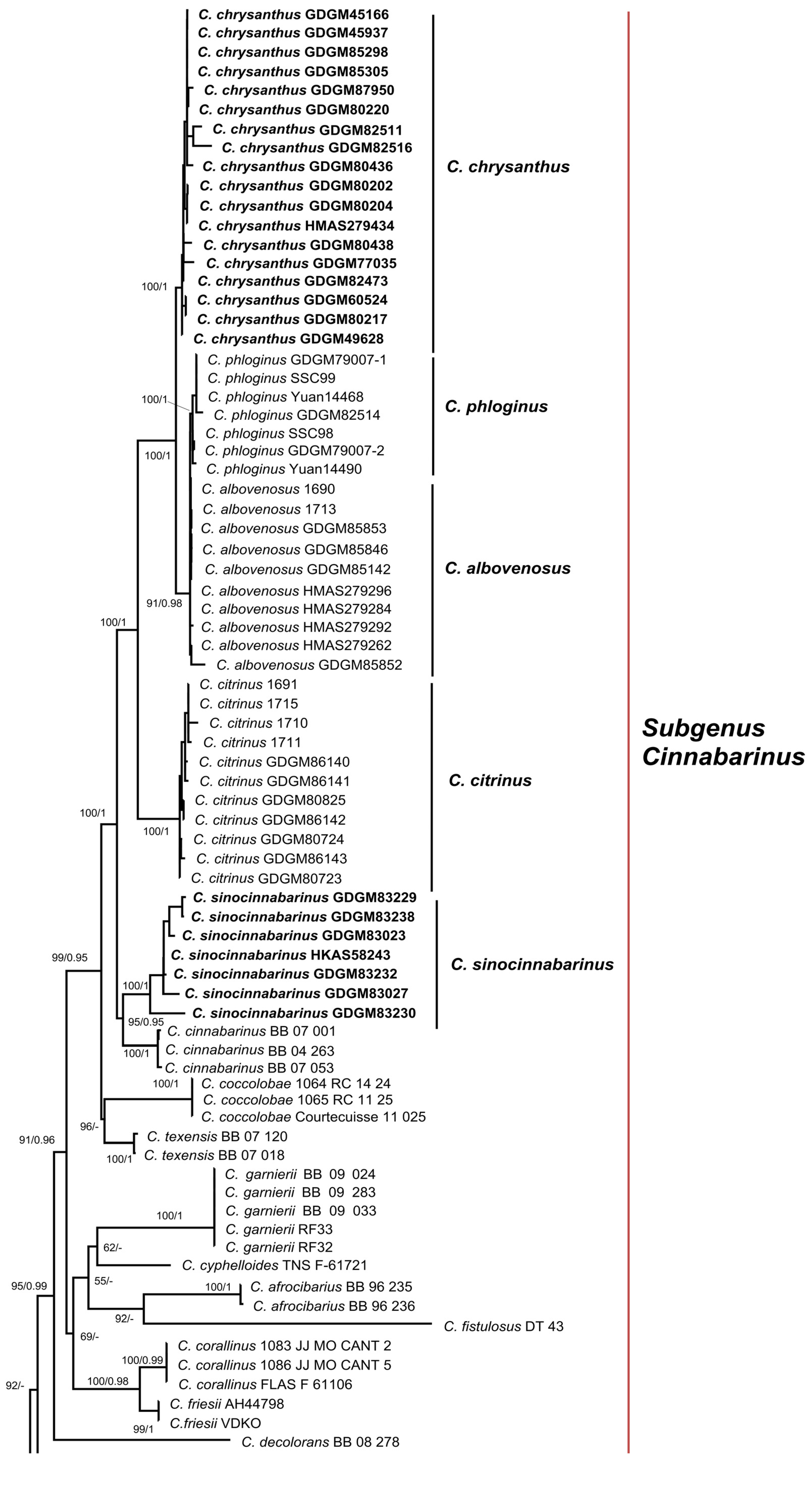
3.1. Molecular Phylogeny
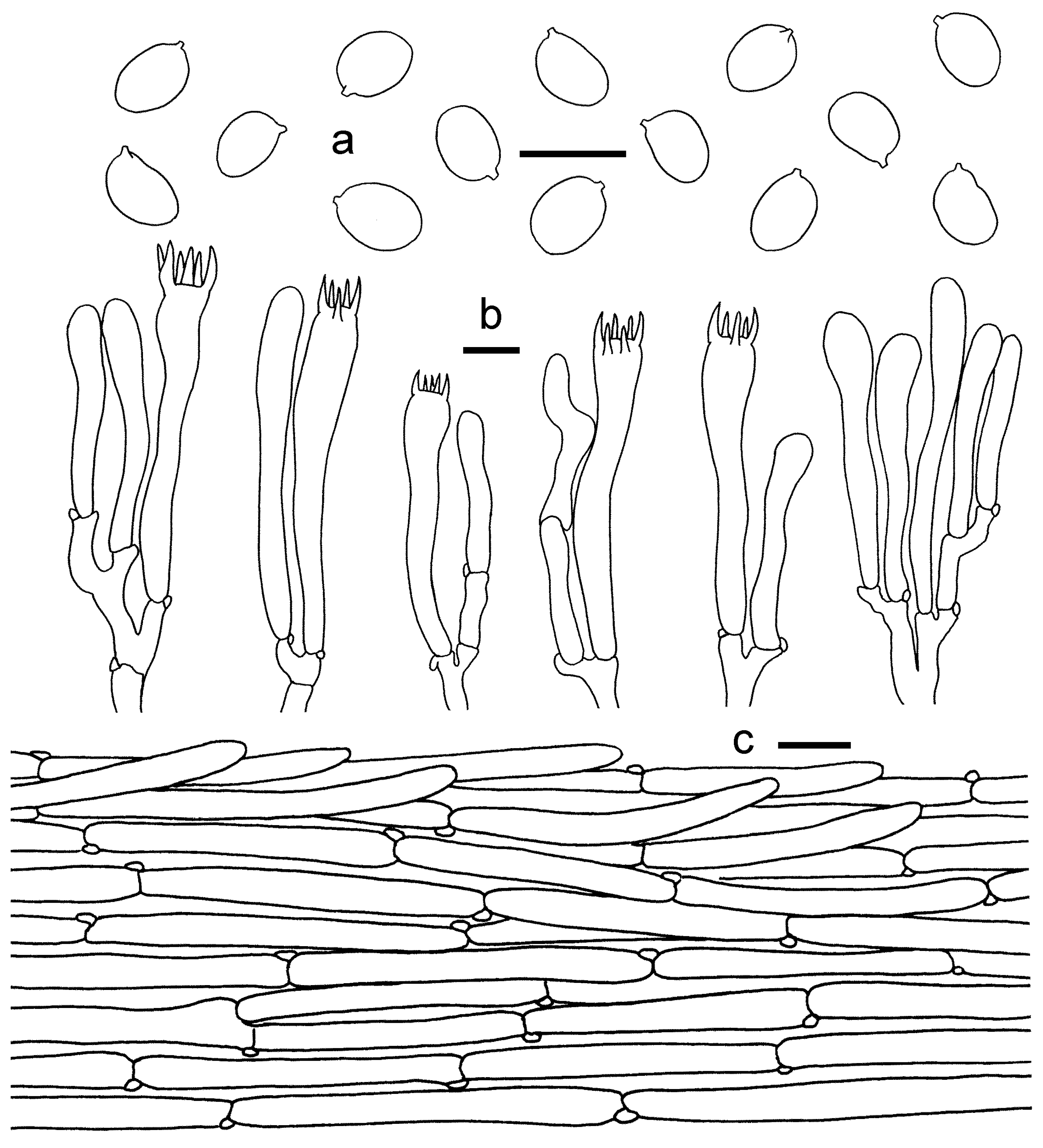
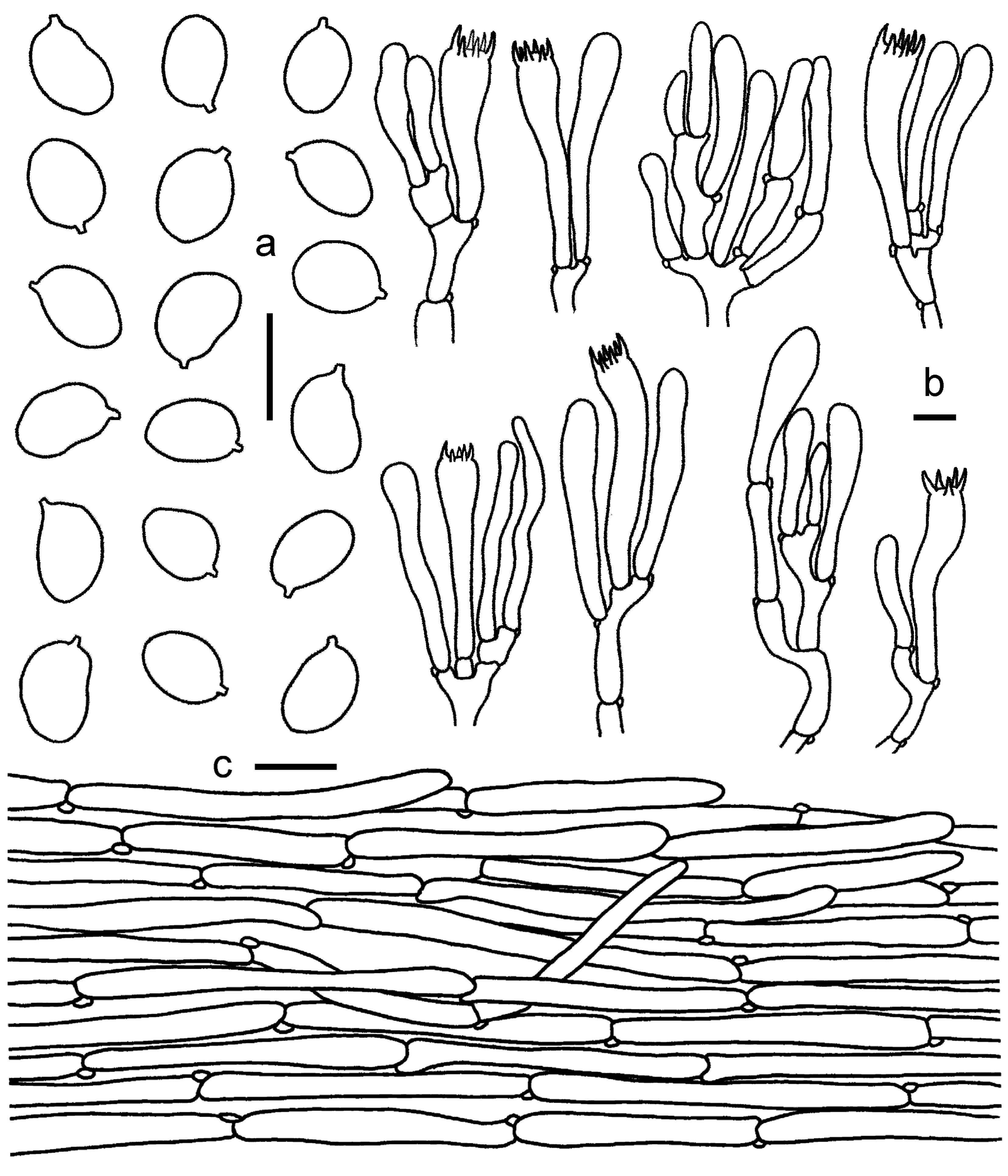
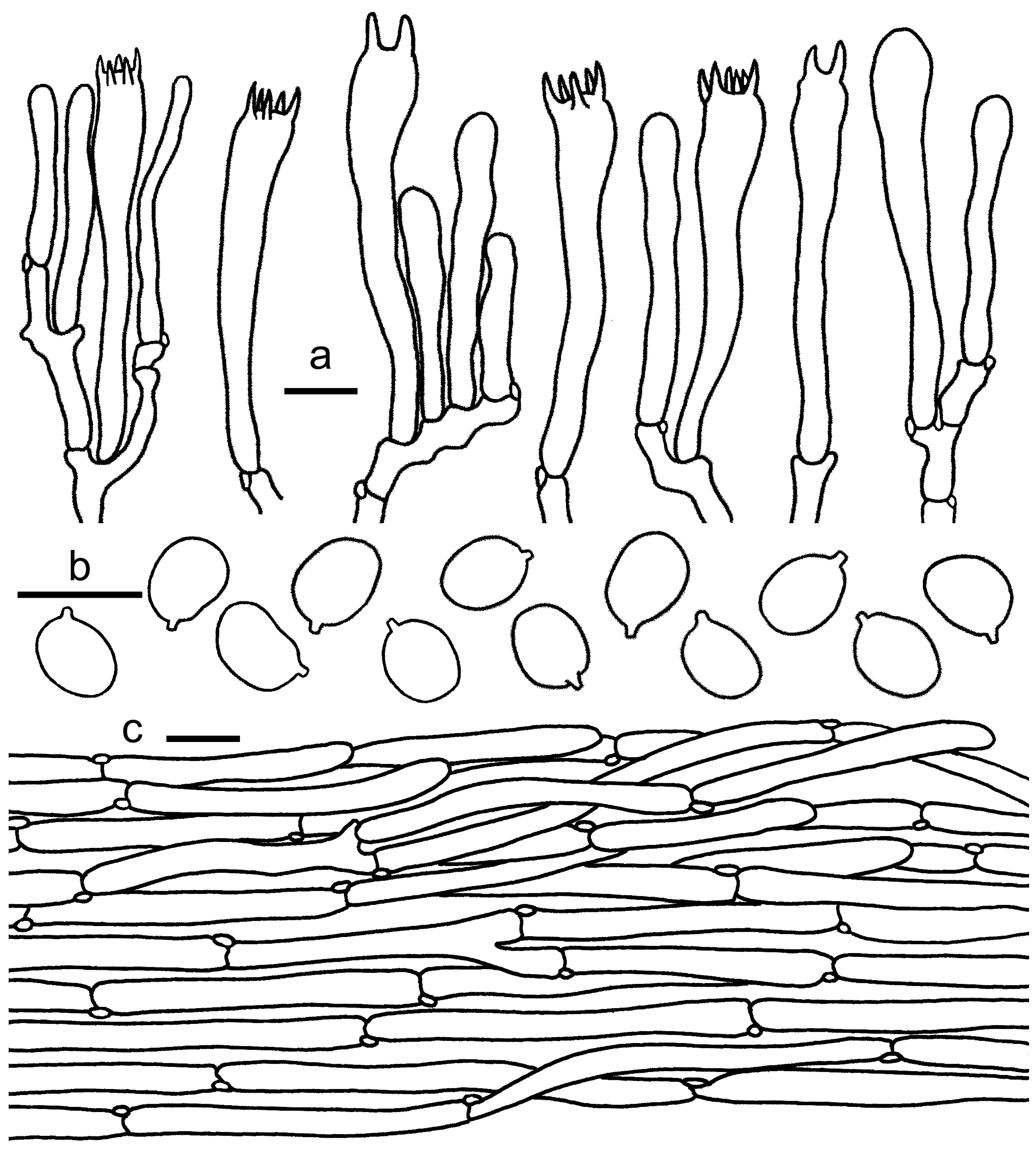
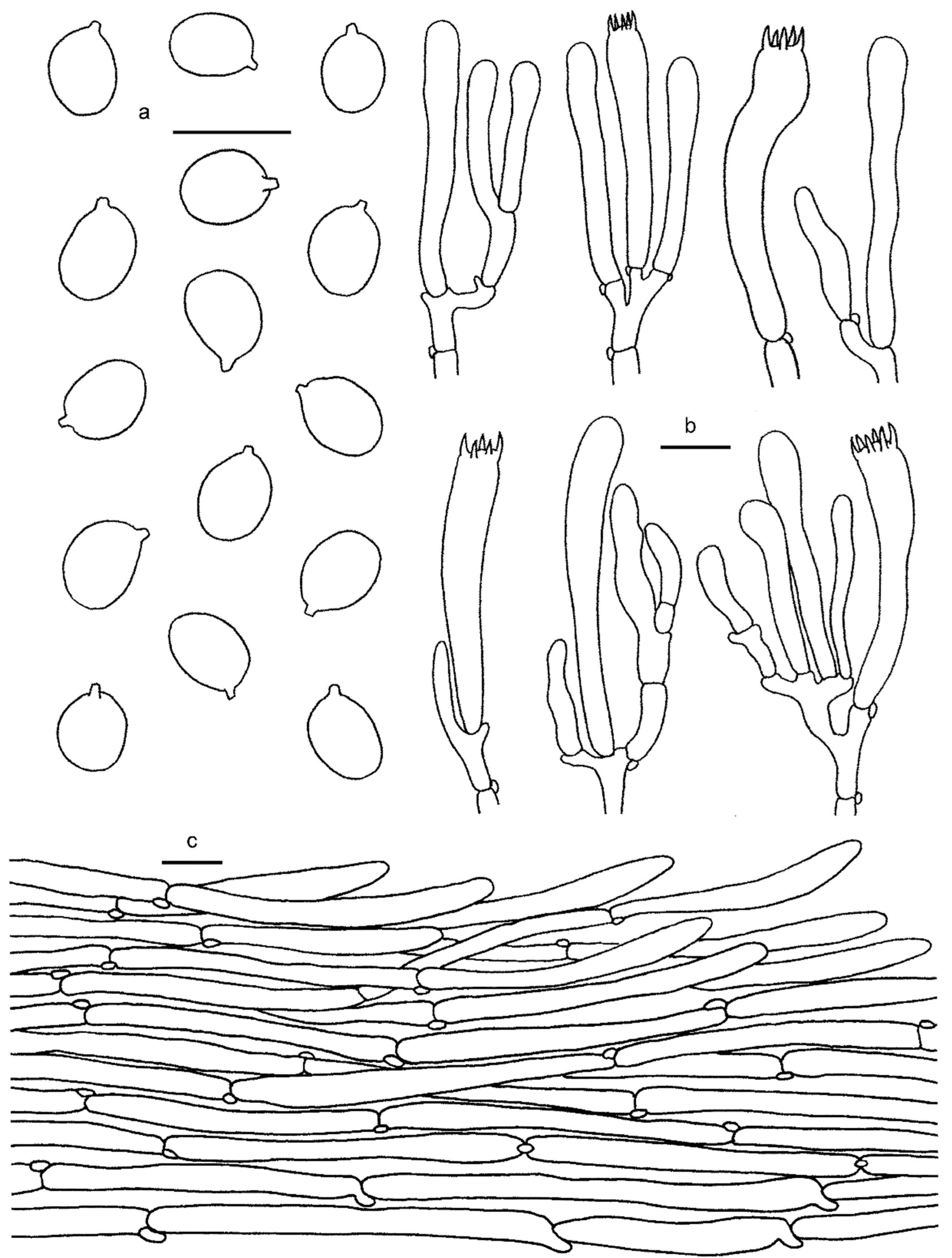
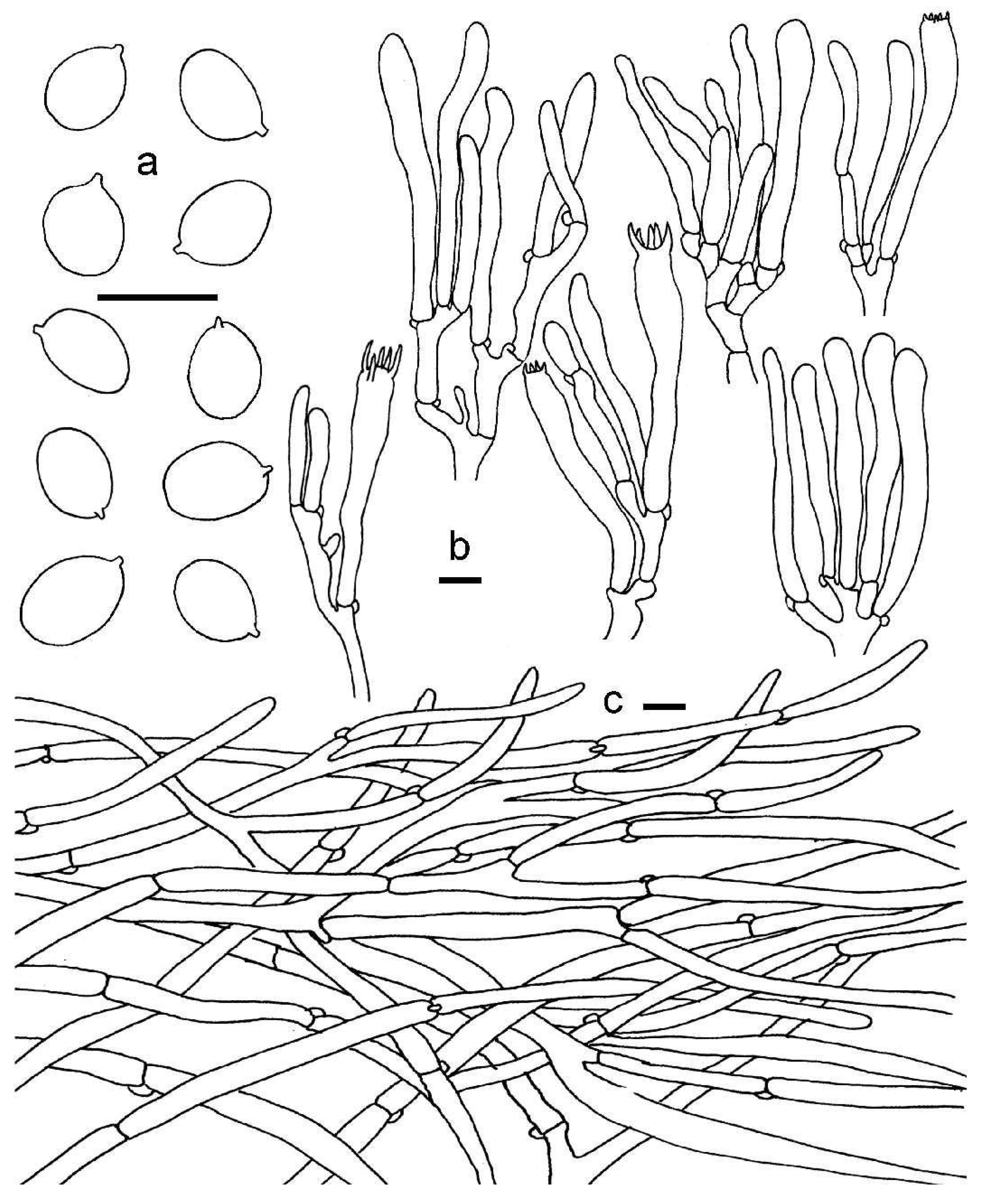
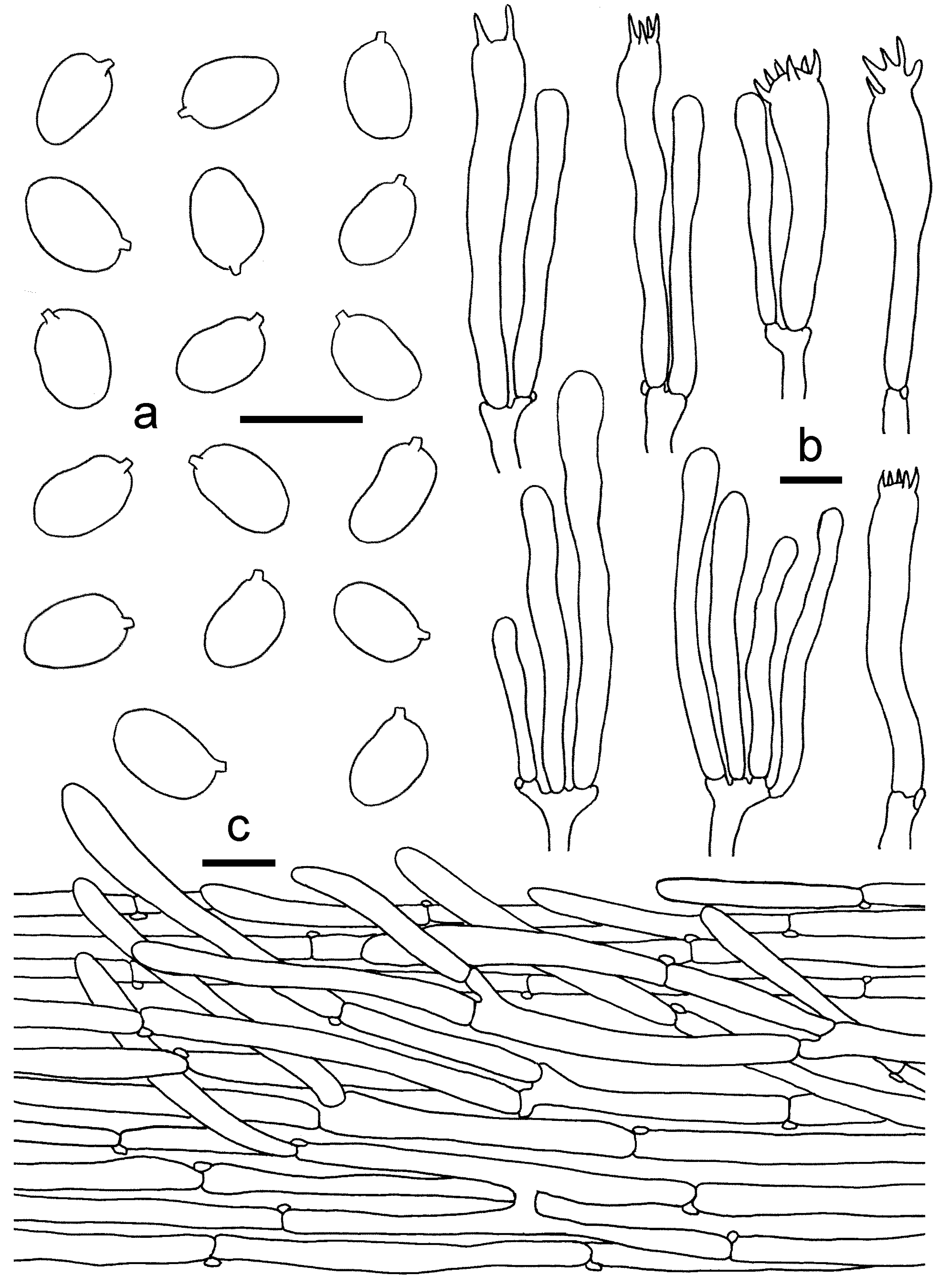
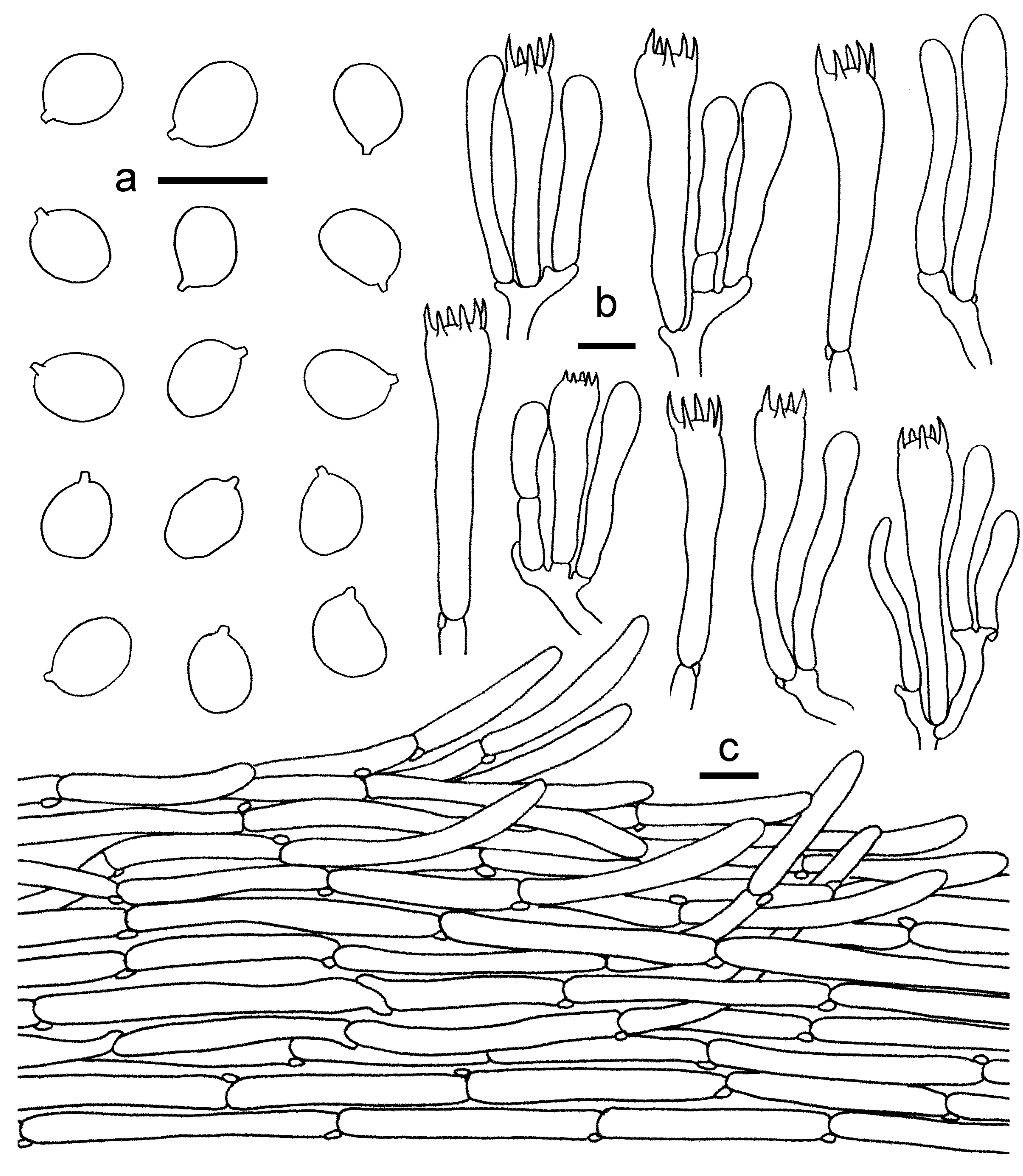
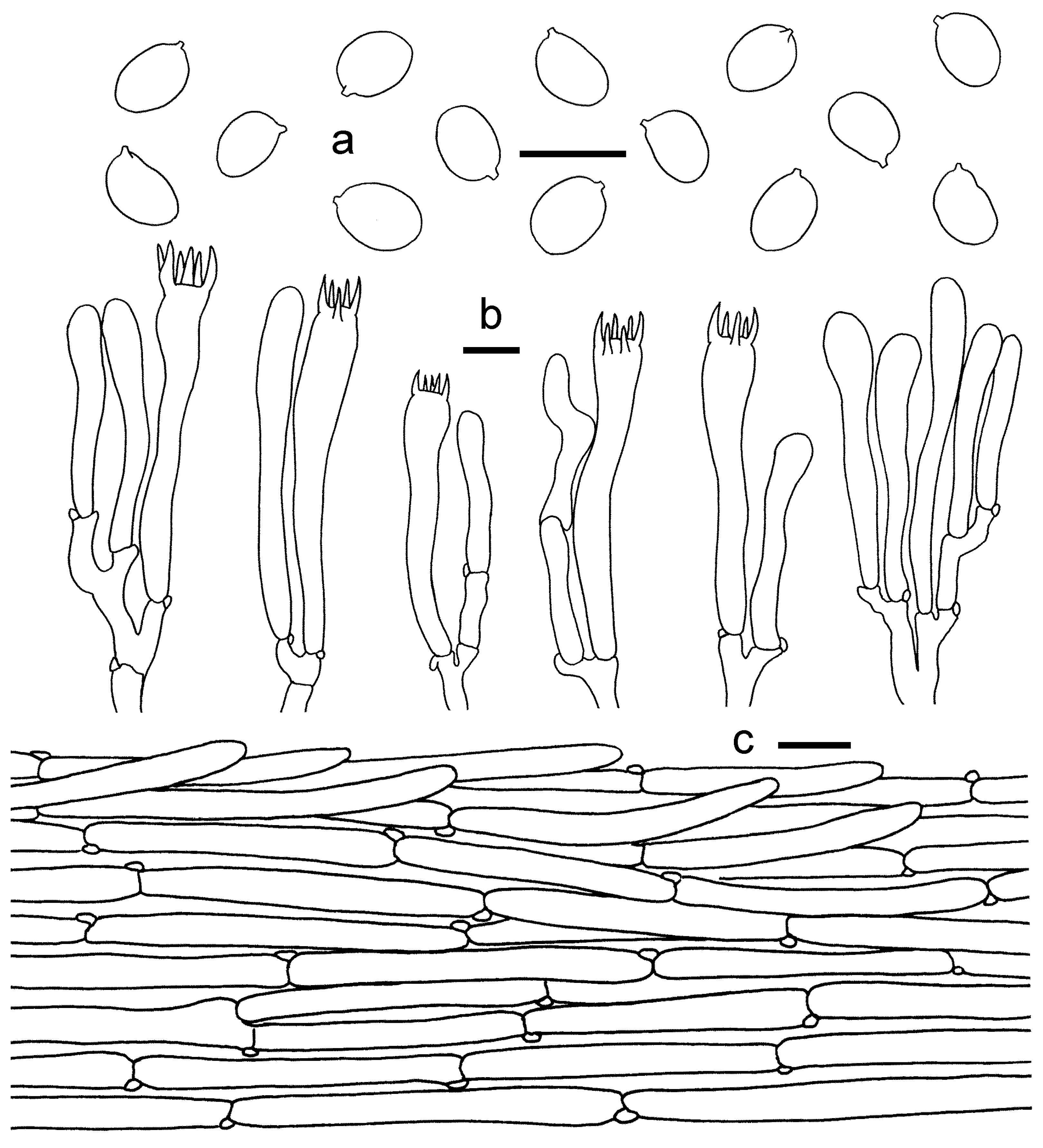
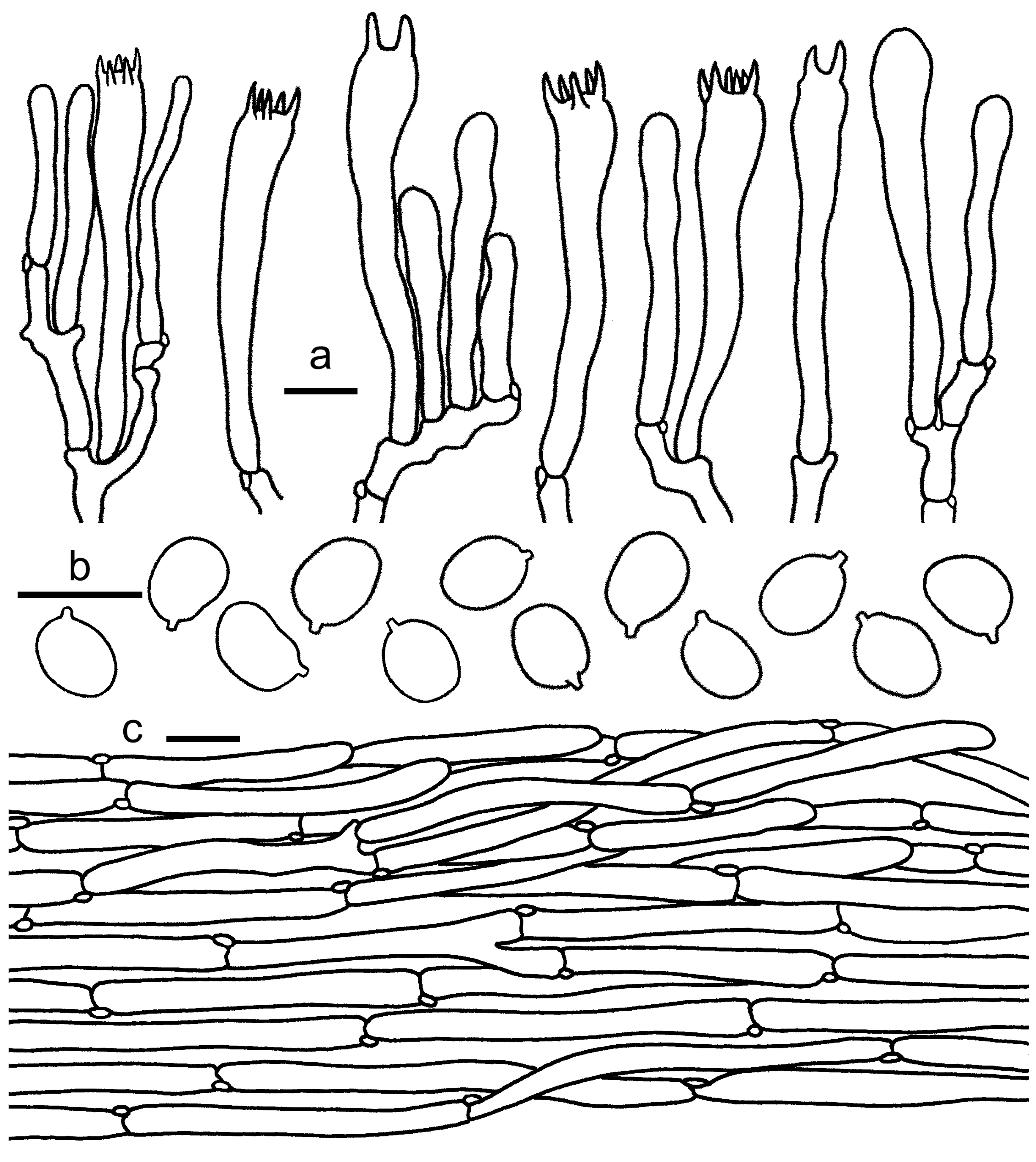
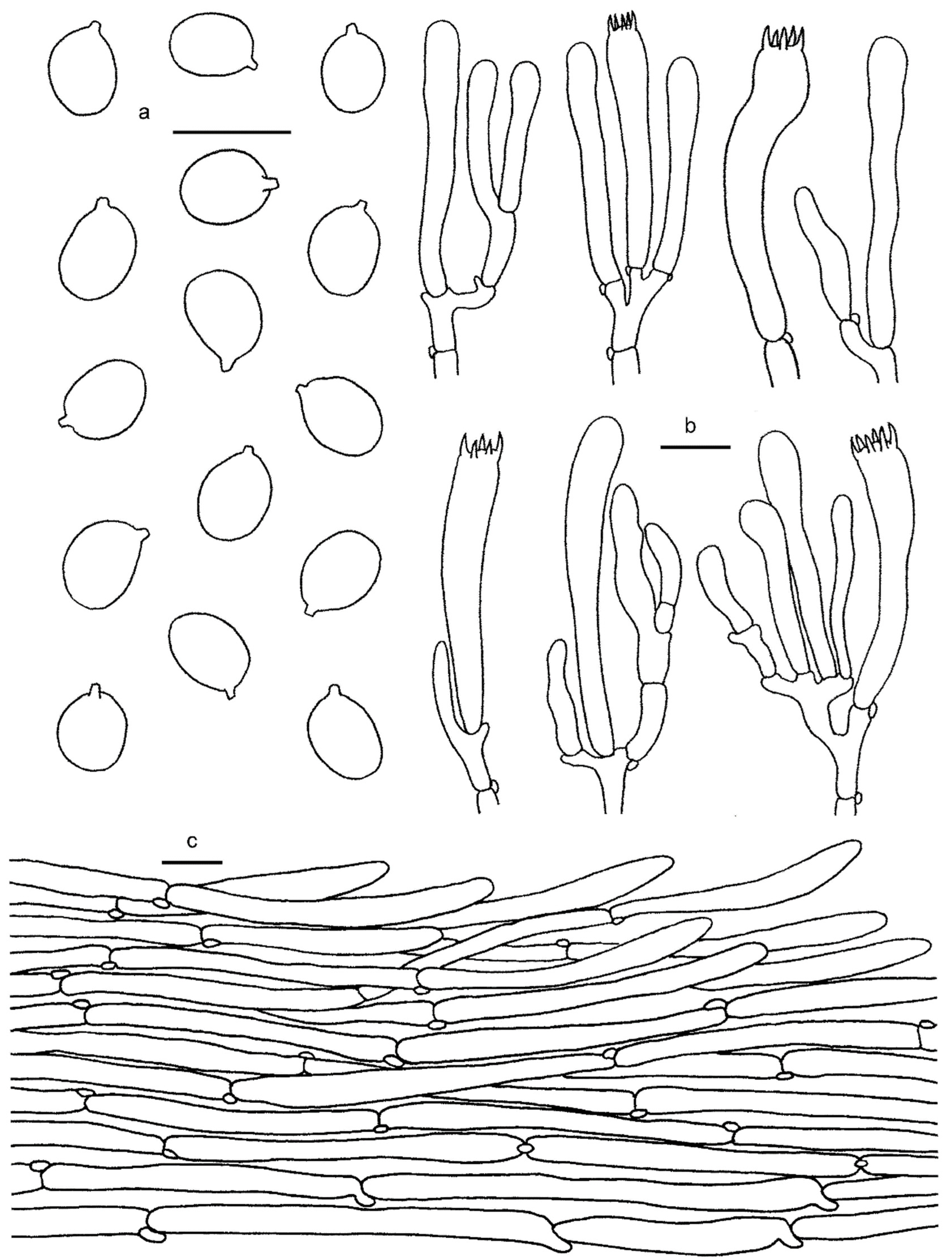
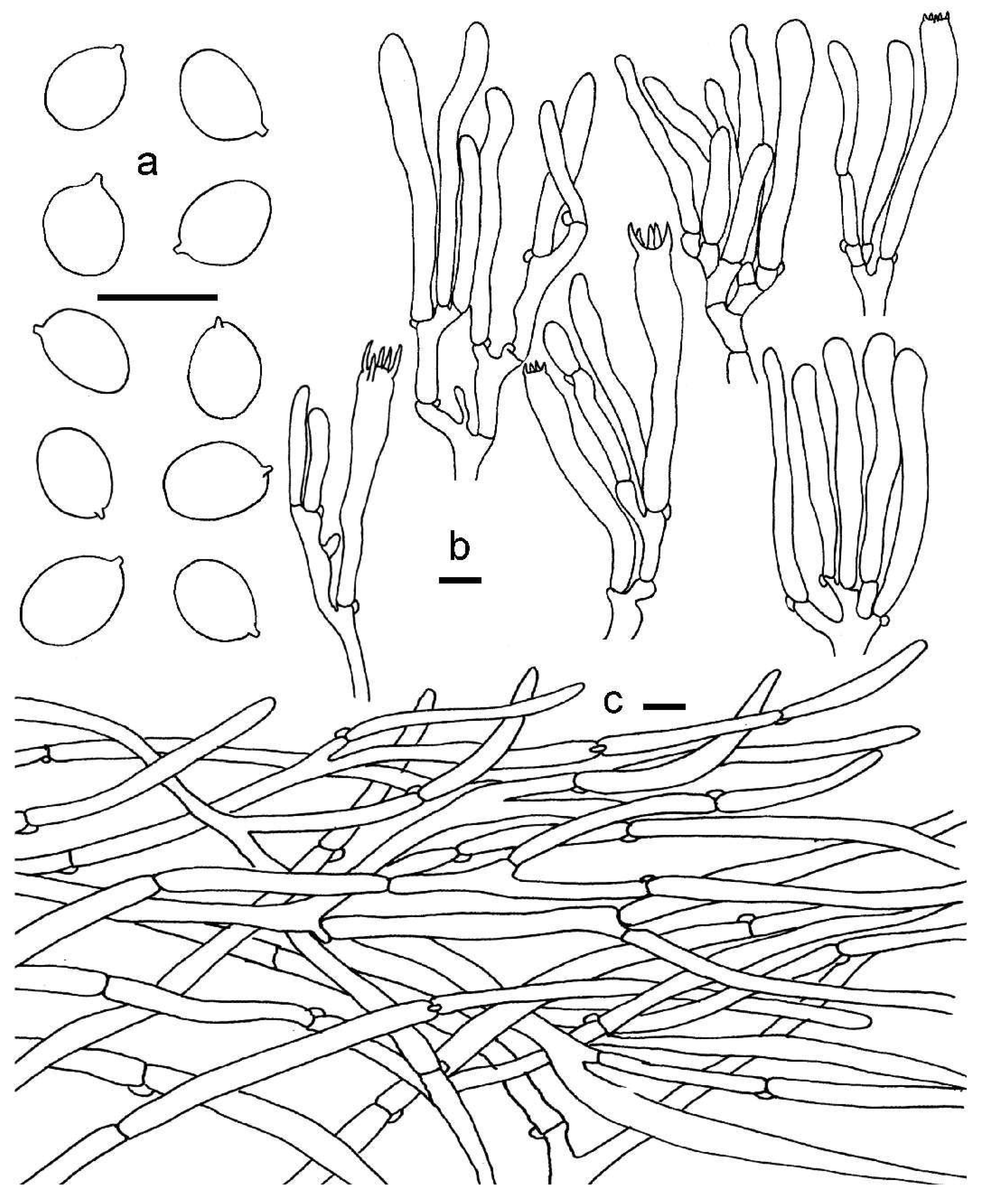
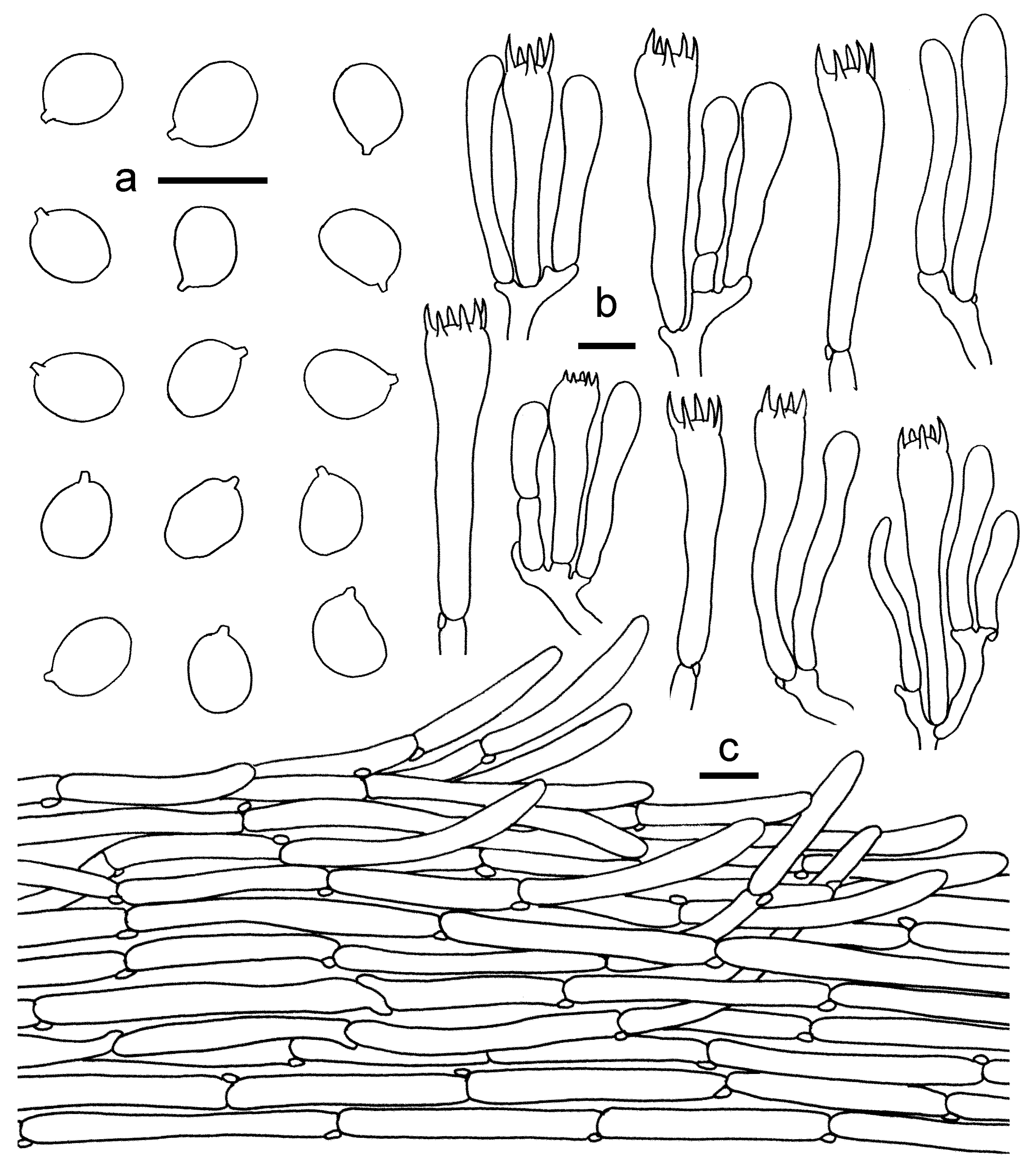
3.2. Taxonomy
3.2.1. Cantharellus subgen. Cinnabarinus Buyck & V. Hofst.
3.2.2. Cantharellus subgen. Parvocantharellus Eyssart. & Buyck
3.3. Key to Species of Subgenus Cinnabarinus in China
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fries, E.M. Systema Mycologicum, Sistens Fungorum Ordines, Genera et Species; Ex Offificina Berlingiana: Lundae, Sweden, 1821; Volume 1. [Google Scholar]
- Kumari, D.; Upadhyay, R.C.; Reddy, M.S. Cantharellus pseudoformosus, a new species associated with Cedrus deodara from India. Mycoscience 2011, 52, 147–151. [Google Scholar] [CrossRef]
- Buyck, B.; Kauff, F.; Eyssartier, G.; Couloux, A.; Hofstetter, V. A multilocus phylogeny for worldwide Cantharellus (Cantharellales, Agaricomycetidae). Fungal Divers. 2014, 64, 101–121. [Google Scholar] [CrossRef]
- Henkel, T.W.; Wilson, A.W.; Amie, M.C.; Dierks, J.; Uehling, J.K.; Roy, M.; Schimann, H.; Wartchow, F.; Mueller, G.M. Cantharellaceae of Guyana II: New species of Craterellus, new South American distribution records for Cantharellus guyanensis and Craterellus excelsus, and a key to the neotropical taxa. Mycologia 2014, 106, 307–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kesel, A.D.; Amalfi, M.; Kasongo Wa Ngoy, B.; Yorou, N.S.; Raspé, O.; Degreef, J.; Buyck, B. New and interesting Cantharellus from tropical Africa. Cryptog. Mycol. 2016, 37, 283–327. [Google Scholar] [CrossRef]
- Ogawa, W.; Endo, N.; Fukuda, M.; Yamada, A. Phylogenetic analyses of Japanese golden chanterelles and a new species description, Cantharellus anzutake sp. nov. Mycoscience 2018, 59, 153–165. [Google Scholar] [CrossRef]
- Cao, T.; Hu, Y.P.; Yu, J.R.; Wei, T.Z.; Yuan, H.S. A phylogenetic overview of the Hydnaceae (Cantharellales, Basidiomycota) with new taxa from China. Stud. Mycol. 2021, 99, 100121. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.C.; Liu, P.G.; Wei, T.Z.; Herrera, M. New insights into the taxonomy of the genus Cantharellus in China: Epityfication of C. yunnanensis W.F. Chiu and the first record of C. cibarius Fr. Cryptog. Mycol. 2021, 42, 25–37. [Google Scholar] [CrossRef]
- Corner, E.J.H. A Monograph of Cantharelloid Fungi; Oxford University Press: Oxford, UK, 1966. [Google Scholar]
- Buyck, B.; Kauff, F.; Cruaud, C.; Hofstetter, V. Molecular evidence for novel Cantharellus (Cantharellales, Basidiomycota) from tropical African miombo woodland and a key to all tropical African chanterelles. Fungal Divers. 2013, 58, 281–298. [Google Scholar] [CrossRef]
- Antonín, V.; Hofstetter, V.; Ryoo, R.; Ka, K.H.; Buyck, B. New Cantharellus species from the Republic of Korea. Mycol. Prog. 2017, 16, 75–759. [Google Scholar] [CrossRef]
- Jian, S.P.; Dai, R.; Gao, J.U.N.; Feng, B. Cantharellus albus, a striking new species from Southwest China. Phytotaxa 2020, 470, 133–144. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.Q.; Buyck, B.; Deng, W.Q.; Li, T.H. Multigene Phylogeny and Morphology Reveal Unexpectedly High Number of New Species of Cantharellus Subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China. J. Fungi 2021, 7, 919. [Google Scholar] [CrossRef] [PubMed]
- An, D.Y.; Liang, Z.Q.; Jiang, S.; Su, M.S.; Zeng, N.K. Cantharellus hainanensis, a new species with a smooth hymenophore from tropical China. Mycoscience 2017, 58, 438–444. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Liang, Z.Q.; Xie, H.J.; Wu, L.L.; Xue, R.; Zeng, N.K. Cantharellus macrocarpus (Cantharellaceae, Cantharellales), a new species from tropical China. Phytotaxa 2021, 484, 170–180. [Google Scholar] [CrossRef]
- Buyck, B.; Hofstetter, V.; Ryoo, R.; Ka, K.-H.; Antonín, V. New Cantharellus species from South Korea. MycoKeys 2020, 76, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Eyssartier, G.; Buyck, B. Note nomenclaturale et systématique sur le genre Cantharellus. Doc. Mycol. 2001, 31, 55–56. [Google Scholar]
- Buyck, B.; Henkel, T.W.; Dentinger, B.T.M.; Séné, O.; Hofstetter, V. Multigene sequencing provides a suitable epitype, barcode sequences and a precise systematic position for the enigmatic, African Cantharellus miniatescens. Cryptog. Mycol. 2016, 37, 269–282. [Google Scholar] [CrossRef]
- Wu, X.L.; Dai, Y.C.; Li, T.H.; Yang, Z.L.; Song, B. Fungi of Tropical China; Science Press: Beijing, China, 2011. [Google Scholar]
- Shao, S.C.; Tian, X.F.; Liu, P.G. Two species with intercontinental disjunct distribution of the genus Cantharellus. J. Yunnan Agric. Univ. 2012, 27, 150–155. [Google Scholar]
- Yang, Z.L.; Wu, G.; Li, Y.C.; Wang, X.H.; Cai, Q. Common Edible and Poisonous Mushrooms of Southwestern China; Science Press: Beijing, China, 2021. [Google Scholar]
- Shao, S.C.; Buyck, B.; Tian, X.F.; Liu, P.G.; Geng, Y.H. Cantharellus phloginus, a new pink-colored species from southwestern China. Mycoscience 2016, 57, 144–149. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben, 3rd ed.; Muster-Schmidt Verlag: Göttingen, Germany, 1981. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Morehouse, E.A.; James, T.Y.; Ganley, A.R.D.; Vilgalys, R.; Berger, L.; Murphy, P.J.; Longcore, J.E. Multilocus sequence typing suggests the chytrid pathogen of amphibians is a recently emerged clone. Mol. Ecol. 2003, 12, 395–403. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Effificient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Moncalvo, J.M.; Nilsson, R.H.; Koster, B.; Dunham, S.M.; Bernauer, T.; Matheny, P.B.; McLenon, T.; Margaritescu, S.; Weiß, M.; Garnica, S.; et al. The cantharelloid clade: Dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 2006, 98, 937–948. [Google Scholar] [CrossRef]
- Buyck, B.; Cruaud, C.; Couloux, A.; Hofstetter, V. Cantharellus texensis sp. nov. from Texas, a Southern lookalike of C. cinnabarinus revealed by tef-1 sequence data. Mycologia 2011, 103, 1037–1046. [Google Scholar] [CrossRef]
- Buyck, B.; Moreau, P.-A.; Courtecuisse, R.; Kong, A.; Roy, M.; Hofstetter, V. Cantharellus coccolobae sp. nov. and Cantharellus garnieri two tropical members of Cantharellus subg. Cinnabarinus. Cryptog. Mycol. 2016, 37, 391–403. [Google Scholar] [CrossRef]
- Buyck, B. Special issue: Cantharellus. Cryptog. Mycol. 2016, 37, 255–258. [Google Scholar] [CrossRef]
- Suhara, H.; Kurogi, S. Cantharellus cyphelloides (Cantharellales), a new and unusual species from a Japanese evergreen broad-leaved forest. Mycol. Prog. 2015, 14, 55. [Google Scholar] [CrossRef]
- Olariaga, I.; Moreno, G.; Manjón, J.L.; Salcedo, I.; Hofstetter, V.; Rodríguez, D.; Buyck, B. Cantharellus (Cantharellales, Basidiomycota) revisited in Europe through a multigene phylogeny. Fungal Divers. 2017, 83, 263–292. [Google Scholar] [CrossRef]
- Ducousso, M.; Contesto, C.; Cossegal, M.; Prin, Y.; Rigault, F.; Eyssartier, G. Cantharellus garnierii sp. nov. from nickel mine maquis in New Caledonia. Cryptog. Mycol. 2004, 25, 115–125. [Google Scholar]
- Buyck, B.; Hofstetter, V. The contribution of tef-1 sequences to species delimitation in the Cantharellus cibarius complex in the southeastern USA. Fungal Divers. 2011, 49, 35–46. [Google Scholar] [CrossRef]
- Buyck, B.; Ndolo Ebika, S.T.; De Kesel, A.; Hofstetter, V. Tropical African Cantharellus Adans.: Fr. (Hydnaceae, Cantharellales) with lilac-purplish tinges revisited. Cryptog. Mycol. 2020, 41, 161–177. [Google Scholar] [CrossRef]
- Feibelman, T.P.; Bennett, J.W.; Cibula, W.G. Cantharellus tabernensis: A new species from the Southeastern United States. Mycologia 1996, 88, 295–301. [Google Scholar] [CrossRef]
- Ryvarden, L.; Petersen, R. Notes on cantharelloid fungi IV. Two new species of Cantharellus. Sven. Bot. Tidskr. 1971, 65, 399–405. [Google Scholar]
- Buyck, B.; Lewis, D.P.; Eyssartier, G.; Hofstetter, V. Cantharellus quercophilus sp. nov. and its comparison to other small, yellow or brown American chanterelles. Cryptog. Mycol. 2010, 31, 17–33. [Google Scholar]
- Petersen, R.H. Notes on Clavarioid Fungi. XIX. Colored illustrations of selected taxa, with comments on Cantharellus. Nova Hedwig. 1985, 42, 151–160. [Google Scholar]
- Kuo, M. Cantharellus persicinus. Retrieved from the MushroomExpert.Com. 2015. Available online: http://www.mushroomexpert.com/cantharellus_persicinus.html (accessed on 20 March 2022).
- Bigelow, H.E. The cantharelloid fungi of New England and adjacent areas. Mycologia 1978, 70, 707–756. [Google Scholar] [CrossRef]
- Montoya, L.; Herrera, M.; Bandala, V.M.; Ramos, A. Two new species and a new record of yellow Cantharellus from tropical Quercus forests in eastern Mexico with the proposal of a new name for the replacement of Craterellus confluens. MycoKeys 2021, 80, 91–114. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wang, C.-Q.; Gan, M.-S.; Li, Y.; Shao, S.-C.; Qin, W.-Q.; Deng, W.-Q.; Li, T.-H. Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records. J. Fungi 2022, 8, 483. https://doi.org/10.3390/jof8050483
Zhang M, Wang C-Q, Gan M-S, Li Y, Shao S-C, Qin W-Q, Deng W-Q, Li T-H. Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records. Journal of Fungi. 2022; 8(5):483. https://doi.org/10.3390/jof8050483
Chicago/Turabian StyleZhang, Ming, Chao-Qun Wang, Man-Shui Gan, Yi Li, Shi-Cheng Shao, Wei-Qiang Qin, Wang-Qiu Deng, and Tai-Hui Li. 2022. "Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records" Journal of Fungi 8, no. 5: 483. https://doi.org/10.3390/jof8050483
APA StyleZhang, M., Wang, C.-Q., Gan, M.-S., Li, Y., Shao, S.-C., Qin, W.-Q., Deng, W.-Q., & Li, T.-H. (2022). Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records. Journal of Fungi, 8(5), 483. https://doi.org/10.3390/jof8050483