
Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens and Strains
2.2. Microscopic Observations
2.3. DNA Extraction, Amplification, and Phylogeny
2.4. Pathogenicity Test
2.5. Temperature, pH, and Carbon Source Tests
2.6. Geographic Distribution
3. Results
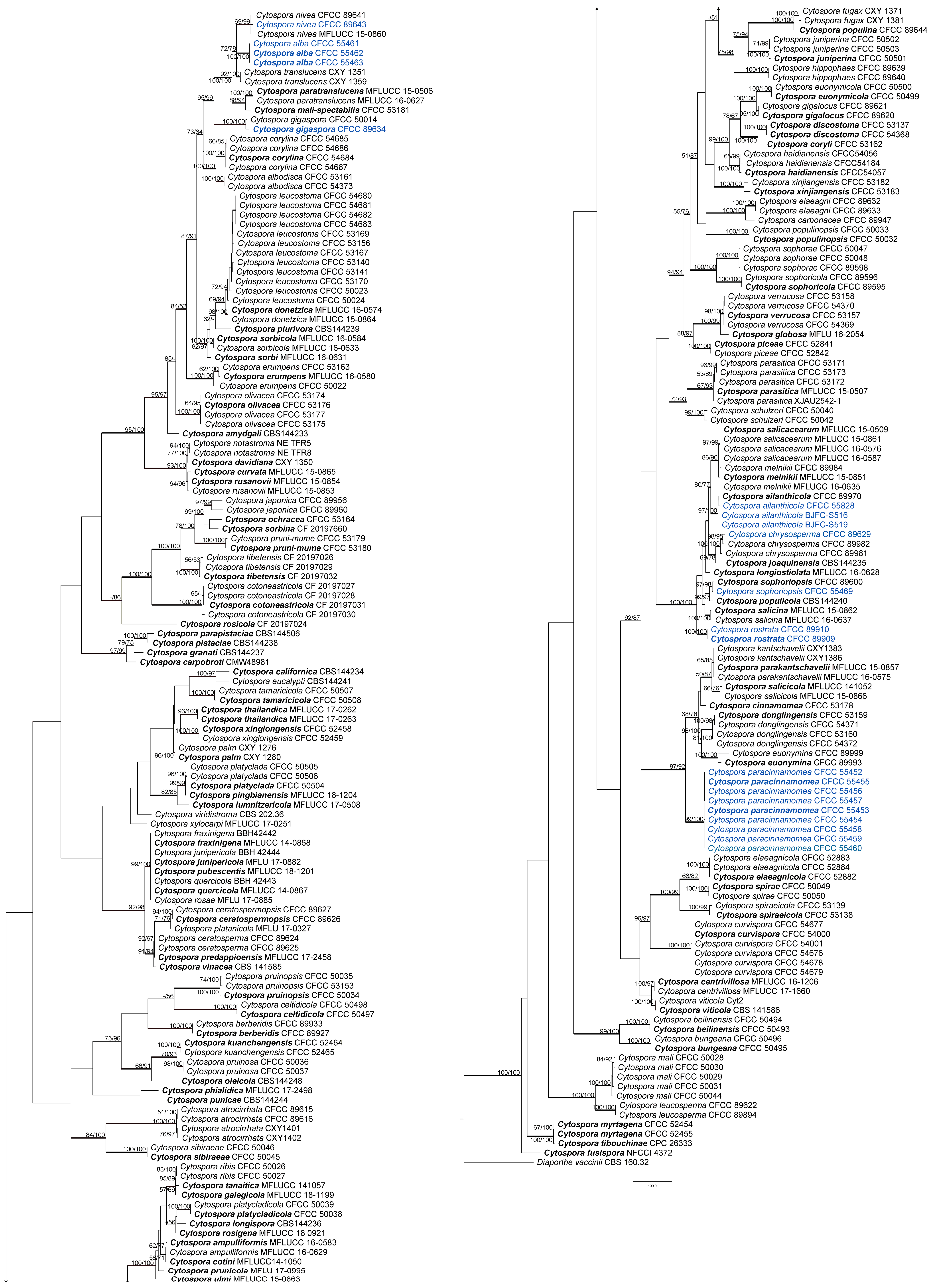
3.1. Phylogenetic Analyses
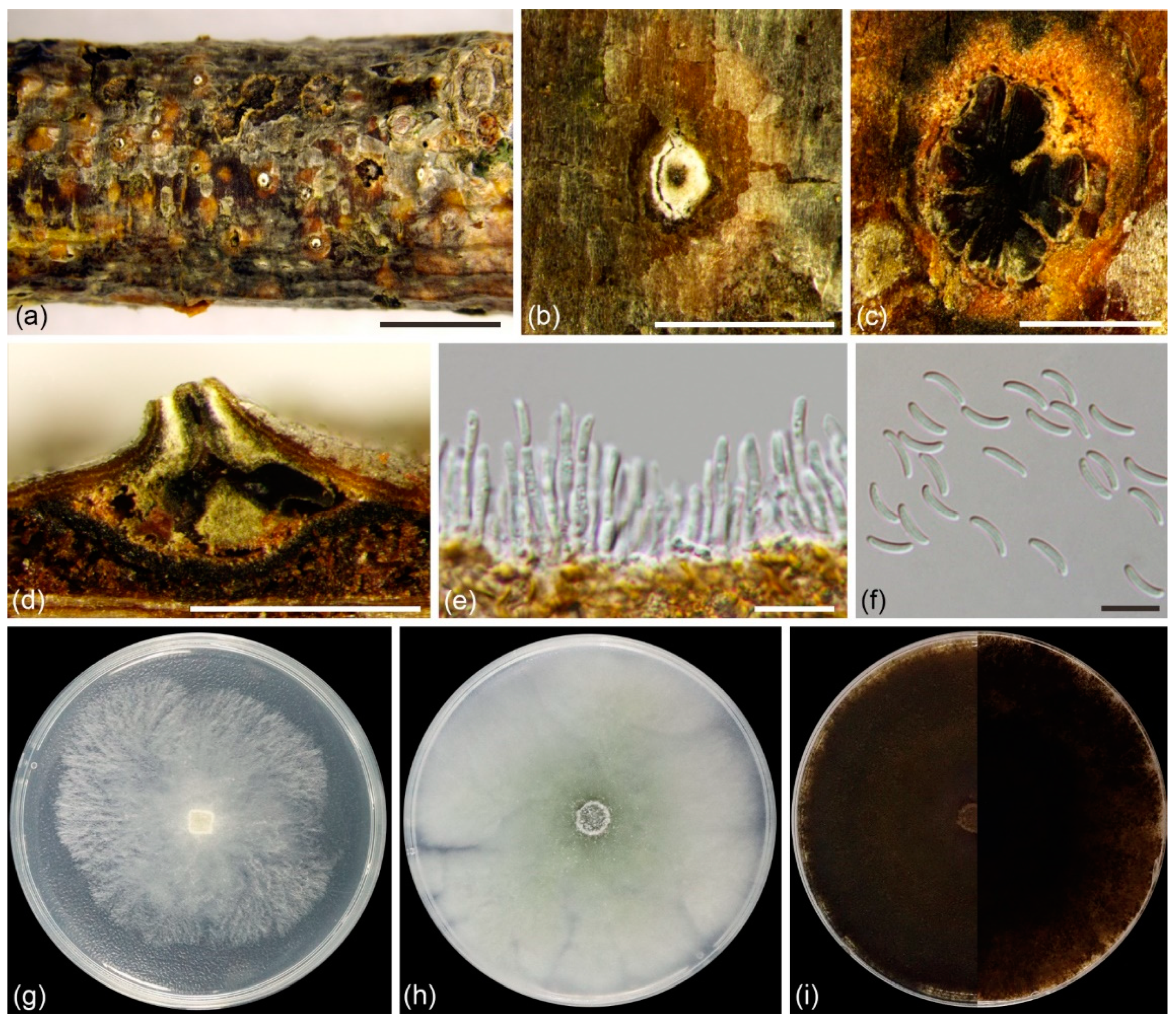
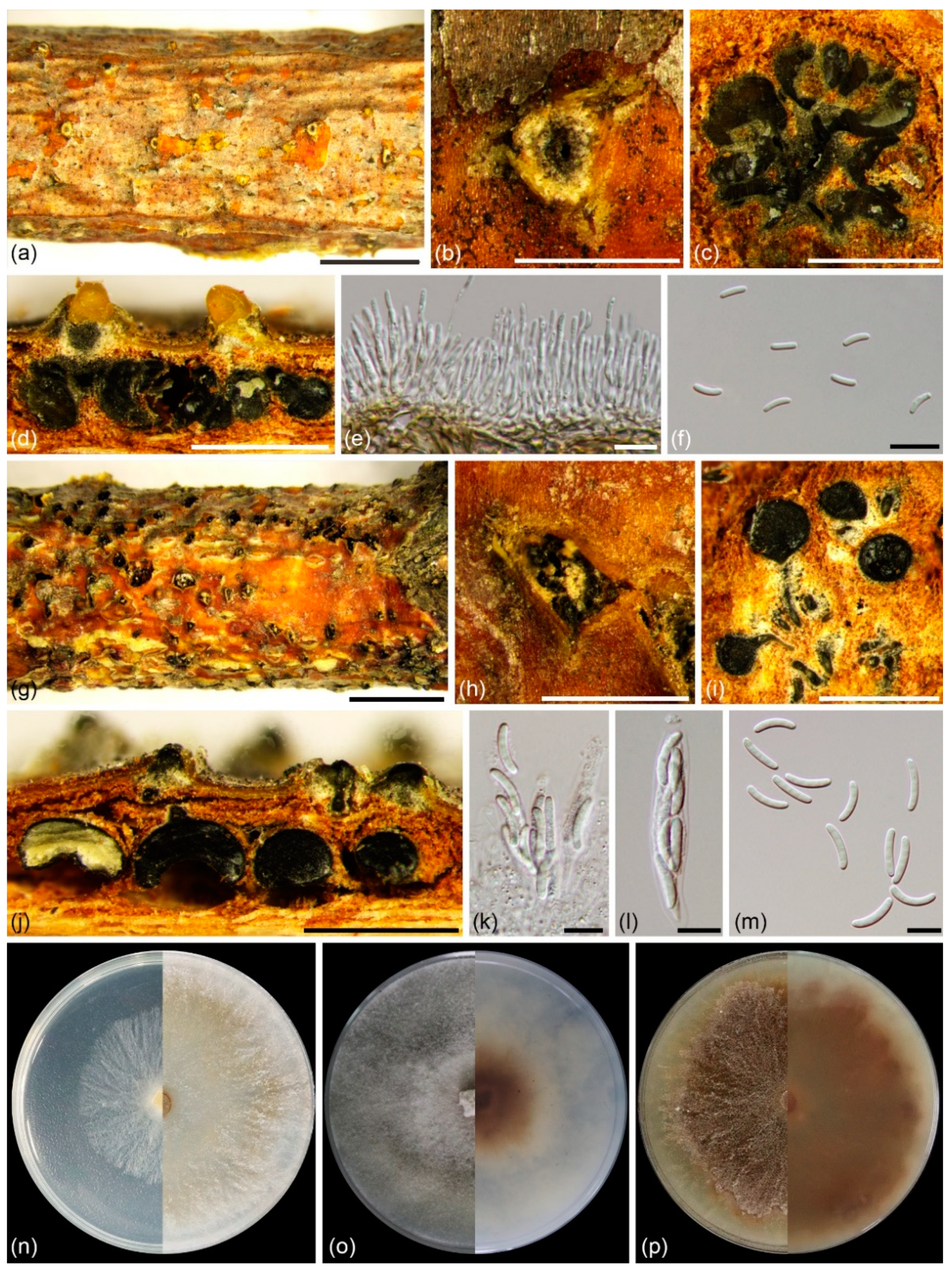
3.2. Taxonomy
3.3. Pathogenicity Test
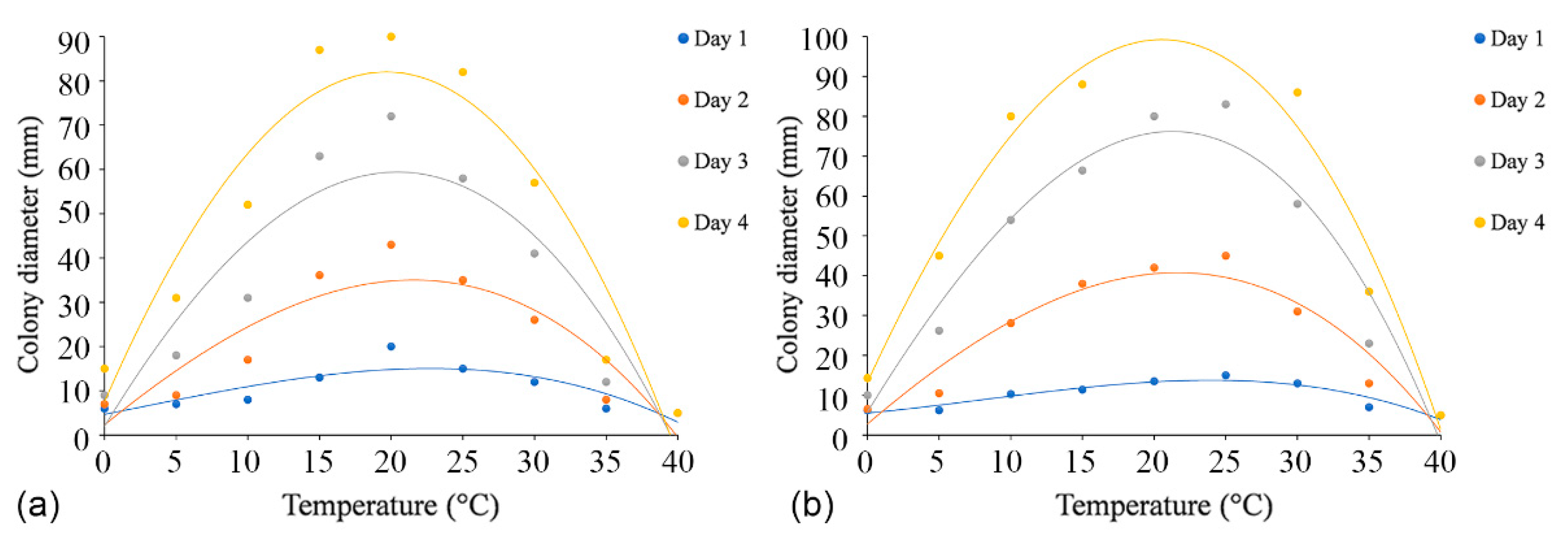
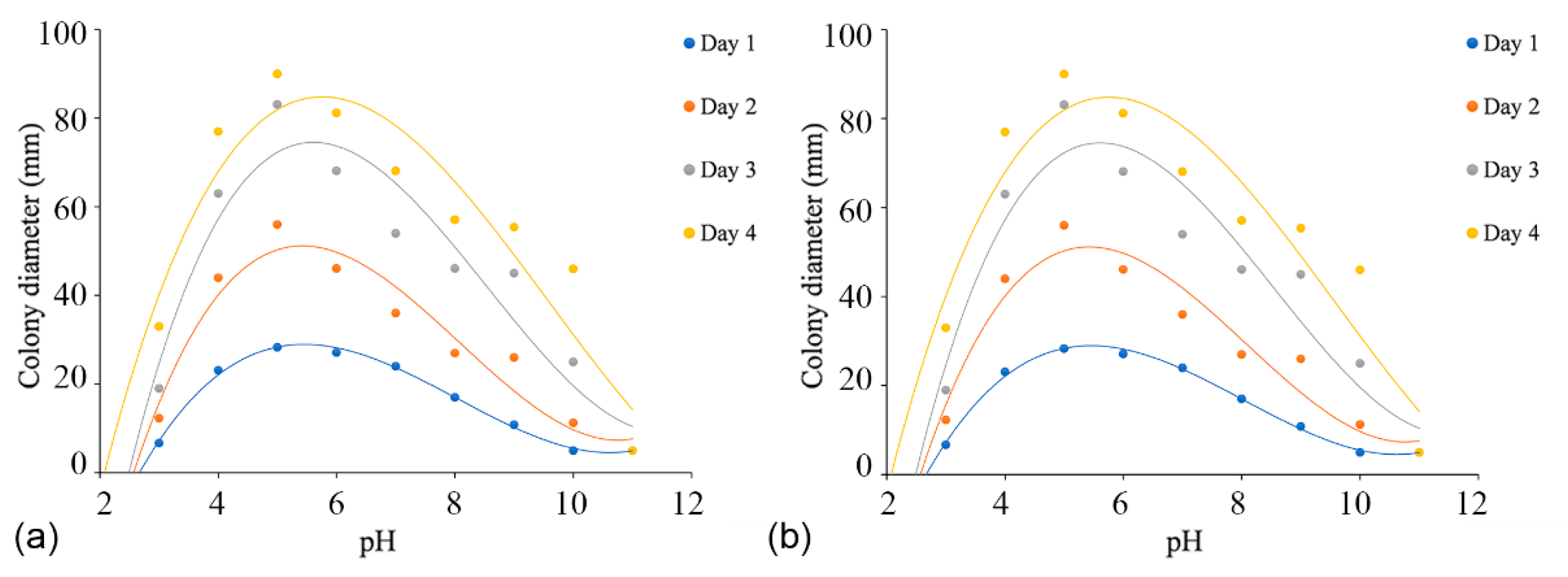
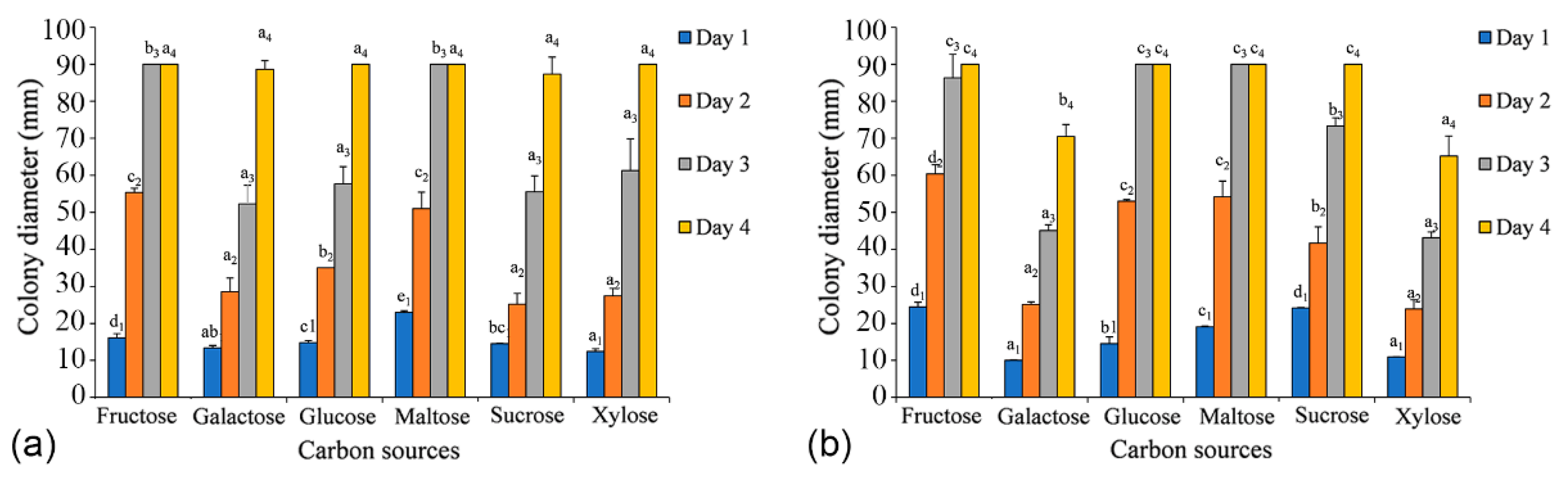
3.4. Temperature, pH, and Carbon Source Tests
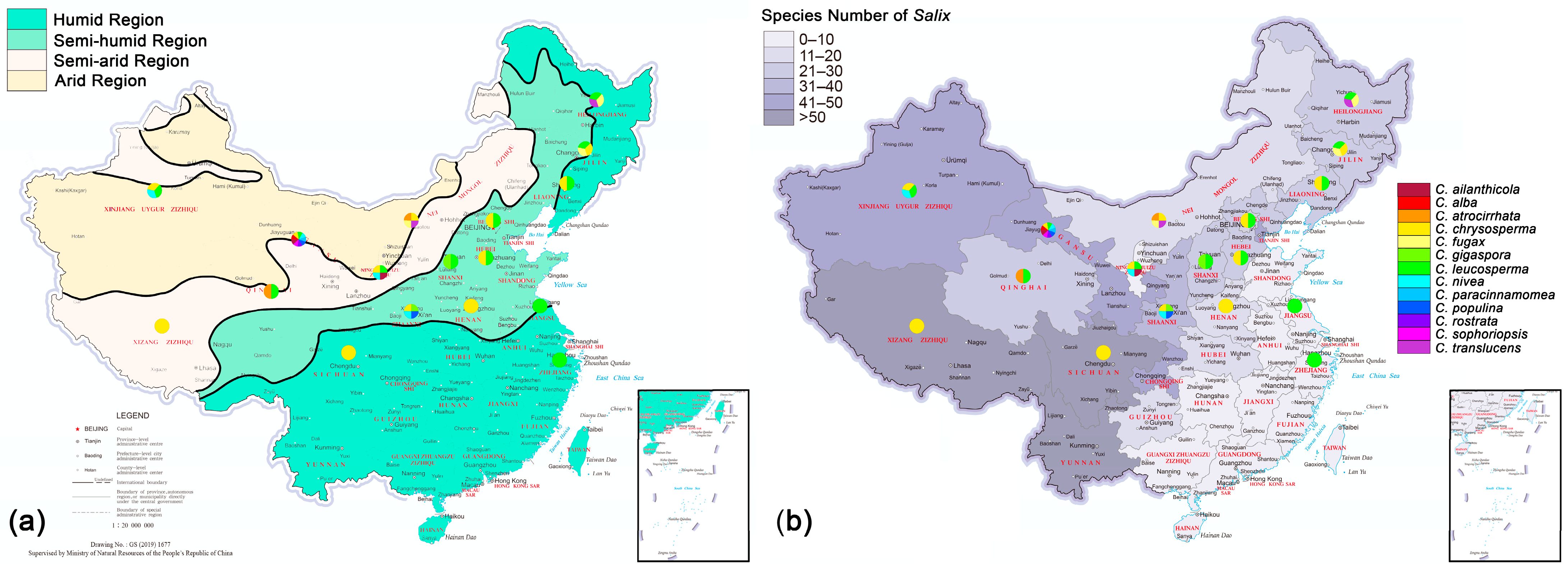
3.5. Geographic Distribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuzovkina, Y.A.; Volk, T.A. The characterization of willow (Salix L.) varieties for use in ecological engineering applications: Co–ordination of structure, function and autecology. Ecol. Eng. 2009, 358, 1178–1189. [Google Scholar] [CrossRef]
- Yu, Z.X.; Qin, G.H.; Jiang, Y.Z.; Song, Y.M.; Qiao, Y.L. Analysis of the reasons for the difficulties in willow classification based on the research status of willow in China. J. Agric. Sci. 2018, 8, 22–25. (In Chinese) [Google Scholar]
- Wang, Y.L.; Lu, Q.; Decock, C.; Li, Y.X.; Zhang, X.Y. Cytospora species from Populus and Salix in China with C. davidiana sp. nov. Fungal Biol. 2015, 119, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Persoonia 2020, 451, 1–45. [Google Scholar] [CrossRef]
- Bubner, B.; Wunder, S.; Zaspel, I.; Zander, M.; Gloger, J.; Fehrenz, S.; Ulrichs, C. Melampsora rust species on biomass willows in central and north-eastern Germany. Fungal Biol. 2014, 118, 910–923. [Google Scholar] [CrossRef]
- Zhao, P.; Tian, C.M.; Yao, Y.J.; Wang, Q.; Yamaoka, Y.; Kakishima, M. Two new species and one new record of Melampsora on willows from China. Mycol. Prog. 2015, 14, 66. [Google Scholar] [CrossRef]
- Yun, Y.H.; Ahn, G.R.; Yoon, S.K.; Kim, H.H.; Son, S.Y.; Kim, S.H. New rust disease of Korean willow (Salix koreensis) caused by Melampsora yezoensis, unrecorded pathogen in Korea. Mycobiology 2016, 444, 335–337. [Google Scholar] [CrossRef]
- Ishaq, A.; Afshan, N.U.S.; Khalid, A.N. First report of Melampsora dimorphospora on white willow in Pakistan. J. Plant Pathol. 2019, 101, 197. [Google Scholar] [CrossRef]
- Stupar, M.; Ljaljevic Grbic, M.; Vukojevic, J.; Lakusic, D. New reports of Melampsora rust (Pucciniomycetes) on the Salix retusa complex in Balkan countries. Bot. Serbica 2020, 44, 89–93. [Google Scholar] [CrossRef]
- Okorski, A.; Pszczolkowska, A.; Sulima, P.; Paukszto, L.; Jastrzebski, J.P.; Przyborowski, J.; Makowczenko, K.G. First report of willow anthracnose caused by Colletotrichum salicis in Poland. Plant Dis. 2018, 102, 2036–2037. [Google Scholar] [CrossRef]
- Masumoto, S.; Tojo, M.; Uchida, M.; Imura, S. Morphological and molecular characterization of Rhytisma filamentosum sp. nov. from Nagano Prefecture, Japan. Mycol. Prog. 2015, 14, 44. [Google Scholar] [CrossRef]
- Jung, B.N.; Park, J.H.; Shin, H.D. First report of Rhytisma filamentosum causing tar-spot disease on Salix koriyanagi. For. Pathol. 2020, 50, e12577. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. (In Chinese) [Google Scholar]
- Wu, C.L. Cytospora canker outbreak in the main city of northern China. China J. For. Pest Dis. 1999, 2, 36. (In Chinese) [Google Scholar]
- Zhang, X.Y.; Luo, Y.Q. Major Forest Diseases and Insect Pests in China; China Forest Press: Beijing, China, 2003. (In Chinese) [Google Scholar]
- Wang, Y.L.; Lu, Q.; Jia, X.Z.; Liang, J.; Zhang, X.Y. First report of branch canker caused by Cytospora atrocirrhata on Populus sp. and Salix sp. in China. Plant Dis. 2013, 97, 426. [Google Scholar] [CrossRef]
- Xu, Y.L. Synthetic Cure and Management of Valsa Sordida Nits of Main Virescence Wood Species in Harbin; Master-Northeast Forestry University: Harbin, China, 2003. (In Chinese) [Google Scholar]
- Adams, G.C.; Wingfield, M.J.; Common, R.; Roux, J. Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Stud. Mycol. 2005, 52, 1–144. [Google Scholar]
- Christensen, C.M. Studies on the biology of Valsa sordida and Cytospora chrysosperma. Phytopathology 1940, 30, 459–475. [Google Scholar]
- Bills, G.F. Isolation and analysis of endophytic fungal communities from woody plants. In Endophytic Fungi in Grasses and Woody Plants; Redlin, S.C., Carris, L.M., Eds.; APS Press: St. Paul, MN, USA, 1996; pp. 31–65. [Google Scholar]
- Ehrenberg, C.G. Sylvae Mycologicae Berolinenses; Formis Theophili Bruschcke: Berlin, Germany, 1818. [Google Scholar]
- Fan, X.L.; Tian, C.M.; Yang, Q.; Liang, Y.M.; You, C.J.; Zhang, Y.B. Cytospora from Salix in northern China. Mycotaxon 2014, 129, 303–315. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Liu, M.; Liang, Y.M.; Tian, C.M. Cytospora species associated with walnut canker disease in China, with description of a new species C. gigalocus. Fungal Biol. 2015, 1195, 310–319. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Ma, R.; Tian, C.M. Cytospora species associated with canker disease of three anti-desertification plants in northwestern China. Phytotaxa 2015, 1974, 227–244. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Crous, P.W.; Liang, Y.M.; Tian, C.M. Cytospora from Ulmus pumila in Northern China. Mycol. Prog. 2015, 14, 74. [Google Scholar] [CrossRef]
- Pan, M.; Zhu, H.Y.; Bonthond, G.; Tian, C.M.; Fan, X.L. High diversity of Cytospora associated with canker and dieback of Rosaceae in China, with 10 new species described. Front. Plant Sci. 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Norphanphoun, C.; Doilom, M.; Daranagama, D.A.; Phookamsak, R.; Wen, T.C.; Bulgakov, T.S.; Hyde, K.D. Revisiting the genus Cytospora and allied species. Mycosphere 2017, 8, 51–97. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Raspé, O.; Jeewon, R.; Wen, T.C.; Hyde, K.D. Morphological and phylogenetic characterisation of novel Cytospora species associated with mangroves. MycoKeys 2018, 38, 93–120. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Holland, L.A.; Nouri, M.T.; Travadon, R.; Abramians, A.; Michailides, T.J.; Trouillas, F.P. Molecular phylogeny of Cytospora species associated with canker diseases of fruit and nut crops in California, with the descriptions of ten new species and one new combination. IMA Fungus 2018, 92, 333–369. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Pan, M.; Bonthond, G.; Tian, C.M.; Fan, X.L. Diaporthalean fungi associated with canker and dieback of trees from Mount Dongling in Beijing, China. MycoKeys 2019, 59, 67–94. [Google Scholar] [CrossRef]
- Jiang, N.; Yang, Q.; Fan, X.L.; Tian, C.M. Identification of six Cytospora species on Chinese chestnut in China. MycoKeys 2020, 62, 1–25. [Google Scholar] [CrossRef]
- Pan, M.; Zhu, H.Y.; Tian, C.M.; Huang, M.R.; Fan, X.L. Assessment of Cytospora strains from conifer cankers in China, with the descriptions of four new Cytospora species. Front. Plant Sci. 2021, 12, 636460. [Google Scholar] [CrossRef]
- Shang, Q.J.; Hyde, K.D.; Camporesi, E.; Maharachchikumbura, S.S.N.; Norphanphoun, C.; Brooks, S.; Liu, J.K. Additions to the genus Cytospora with sexual morph in Cytosporaceae. Mycosphere 2020, 11, 189–224. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: London, UK, 1970. [Google Scholar]
- Doyle, J.J. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 304, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony, * and Other Methods, version 4.0b10; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, H.O. New algorithms and methods to estimate maximum–likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 593, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. Mr. Bayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A. FigTree, version 1.3.1; Institute of Evolutionary Biology: Edinburgh, UK, 2010. [Google Scholar]
- Koch, R. Uber bakteriologische forschung. In Proceedings of the Xth International Congress of Medicine, Berlin, Germany, 4 August 1890; Volume 1, p. 35. [Google Scholar]
- Zhou, X.; Pan, M.; Li, H.Y.; Tian, C.M.; Fan, X.L. Dieback of Euonymus alatus (Celastraceae) caused by Cytospora haidianensis sp. nov. in China. Forests 2020, 115, 524. [Google Scholar] [CrossRef]
- Gao, H.; Pan, M.; Tian, C.M.; Fan, X.L. Cytospora and Diaporthe species associated with hazelnut canker and dieback in Beijing, China. Front. Cell. Infect. Microbiol. 2021, 11, 664366. [Google Scholar] [CrossRef]
- Wu, S.H.; Yin, Y.H.; Zheng, D.; Yang, Q.Y. Study on dry and wet conditions of land surface in China in recent 30 years. Sci. China 2005, 3, 276–283. (In Chinese) [Google Scholar]
- Fang, Z.F.; Zhao, S.D.; Alexei, K.S. Salicaceae. In Cochairs of the Editorial Committee, Flora of China; Wu, Z.Y., Peter, H.R., Eds.; Science Press: Beijing, China, 2004; Volume 4. [Google Scholar]
- Zheng, W.J. Sylva Sinica; China Forestry Publishing House: Beijing, China, 1985. (In Chinese) [Google Scholar]
- Fang, J.Y.; Wang, Z.H.; Tang, Z.Y. Atlas of Woody Plant in China—Distribution and Climate; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963. (In Chinese) [Google Scholar]
- Wei, J.C. Identification of Fungus Handbook; Science Press: Shanghai, China, 1979. (In Chinese) [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Spielman, L.J. A monograph of Valsa on hardwoods in North America. Can. J. Bot. 1985, 63, 1355–1378. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from fagaceae leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Piao, C.G.; Li, Y. Two new species of Diaporthe (Diaporthaceae, Diaporthales) associated with tree cankers in the Netherlands. MycoKeys 2021, 85, 31. [Google Scholar] [CrossRef]
- Chen, M.M. Forest fungi Phytogeography: Forest fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; Pacific Mushroom Research & Education Center: Sacramento, CA, USA, 2003. [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China; Ithaca: New York, NY, USA, 2005. (In Chinese) [Google Scholar]
- Gvritishvili, M.N. New species of fungi belonging to genus Cytospora Fr. II. Mikol. Fitopatol. 1973, 7, 544–549. (In Russian) [Google Scholar]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 9, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Kepley, J.; Jacobi, W.R. Pathogenicity of Cytospora fungi on six hardwood species. J. Arboric. 2000, 26, 326–333. [Google Scholar] [CrossRef]
- McIntyre, G.A.; Jacobi, W.R.; Ramaley, A.W. Factors affecting Cytospora canker occurrence on aspen. J. Arboric. 1996, 22, 229–233. [Google Scholar]
- Kaczynski, K.M.; Cooper, D.J. Susceptibility of Salix monticola to Cytospora canker under increased temperatures and decreased water levels. For. Ecol. Manag. 2013, 305, 223–228. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.; Pan, M.; Tian, C.; Fan, X. Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China. J. Fungi 2022, 8, 377. https://doi.org/10.3390/jof8040377
Lin L, Pan M, Tian C, Fan X. Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China. Journal of Fungi. 2022; 8(4):377. https://doi.org/10.3390/jof8040377
Chicago/Turabian StyleLin, Lu, Meng Pan, Chengming Tian, and Xinlei Fan. 2022. "Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China" Journal of Fungi 8, no. 4: 377. https://doi.org/10.3390/jof8040377
APA StyleLin, L., Pan, M., Tian, C., & Fan, X. (2022). Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China. Journal of Fungi, 8(4), 377. https://doi.org/10.3390/jof8040377