
Volatiles Produced by Yeasts Related to Prunus avium and P. cerasus Fruits and Their Potentials to Modulate the Behaviour of the Pest Rhagoletis cerasi Fruit Flies

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Sampling, Culturing, and Identification
2.2. Insects
2.3. Sampling and Analysis of Volatiles Produced by Yeasts
2.4. Gas Chromatography-Electroantennogram Detection
2.5. Electroantennogram Dose-Response
2.6. Behavioural Assay
2.7. Statistical Analysis
3. Results
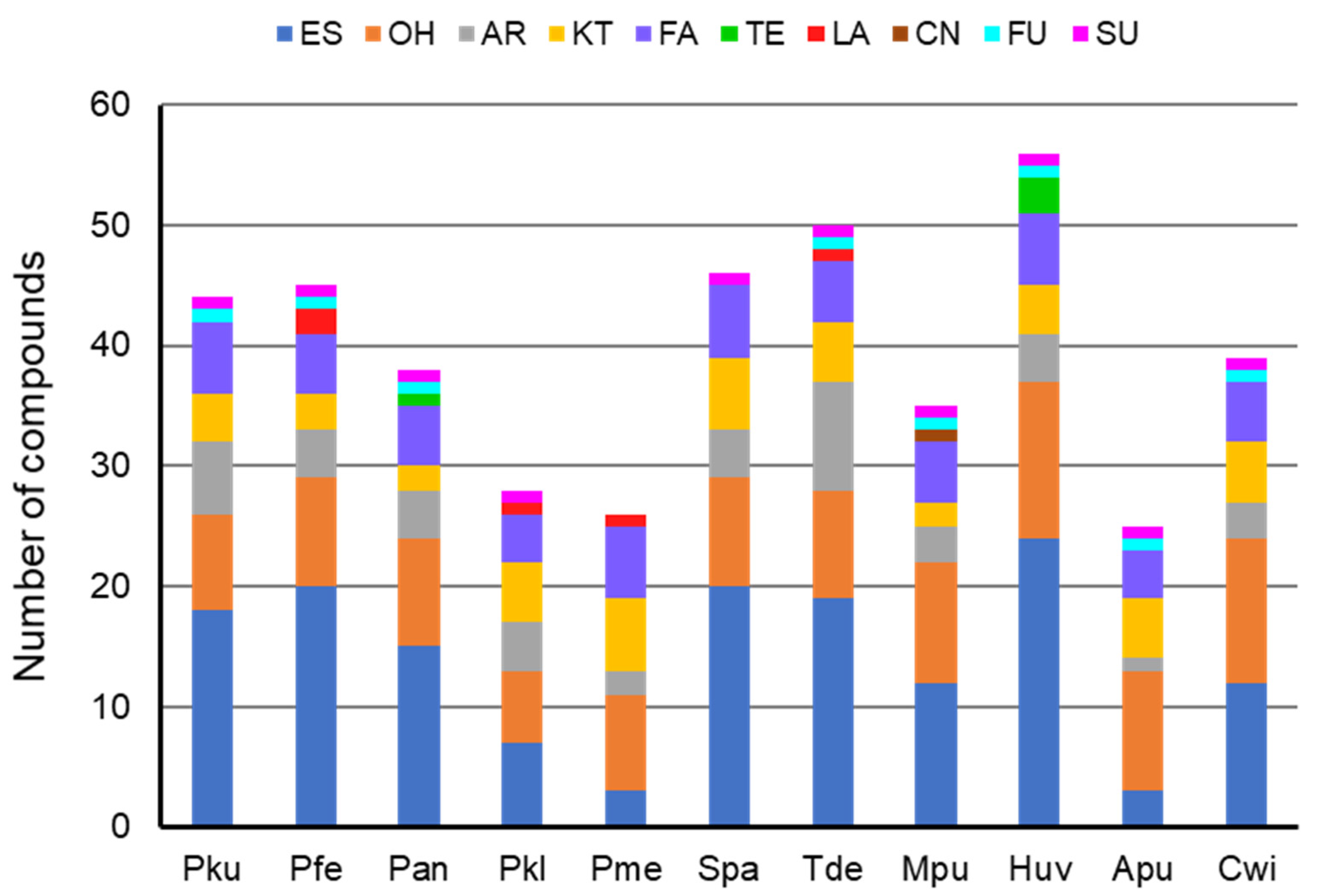
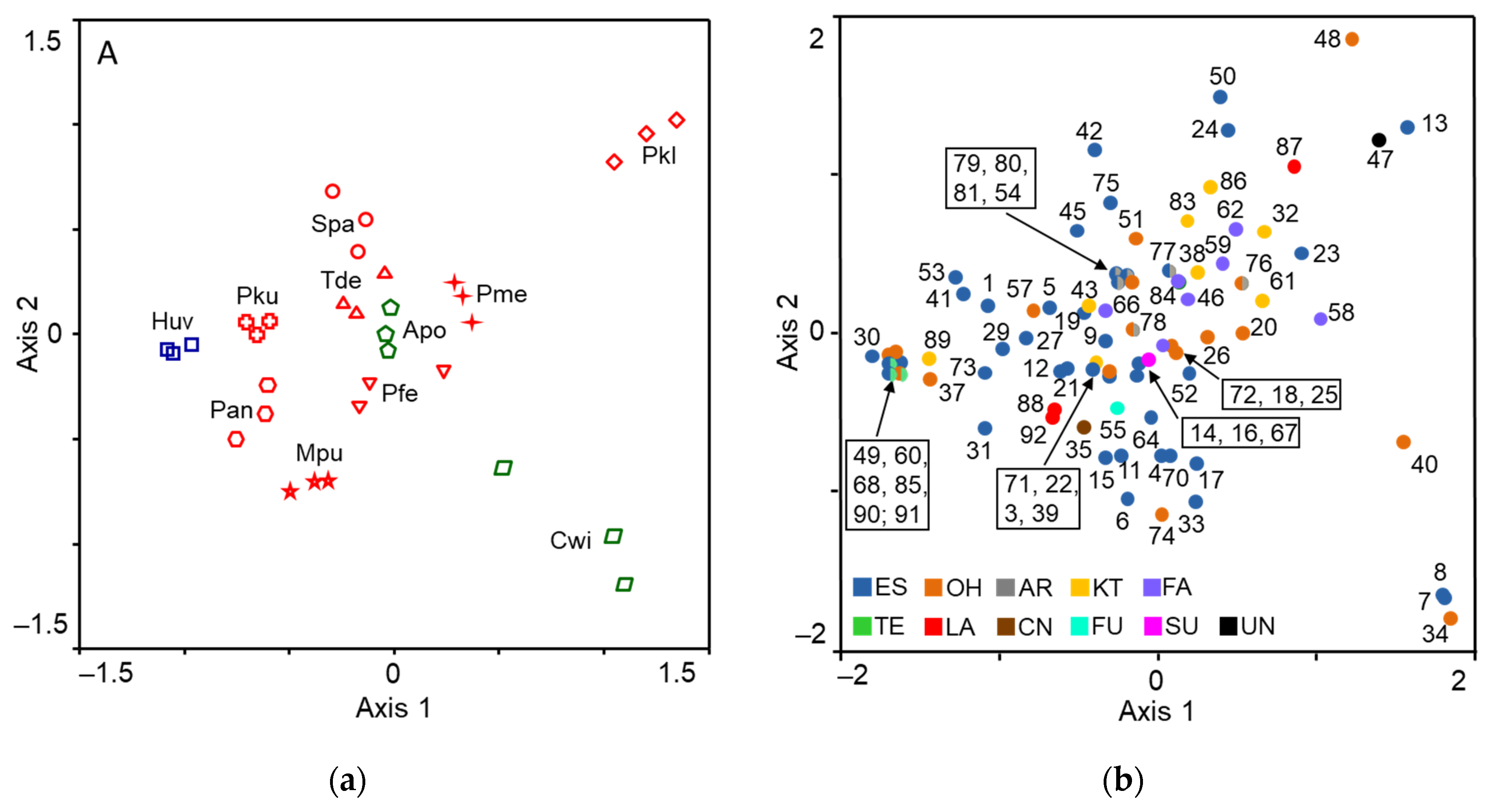
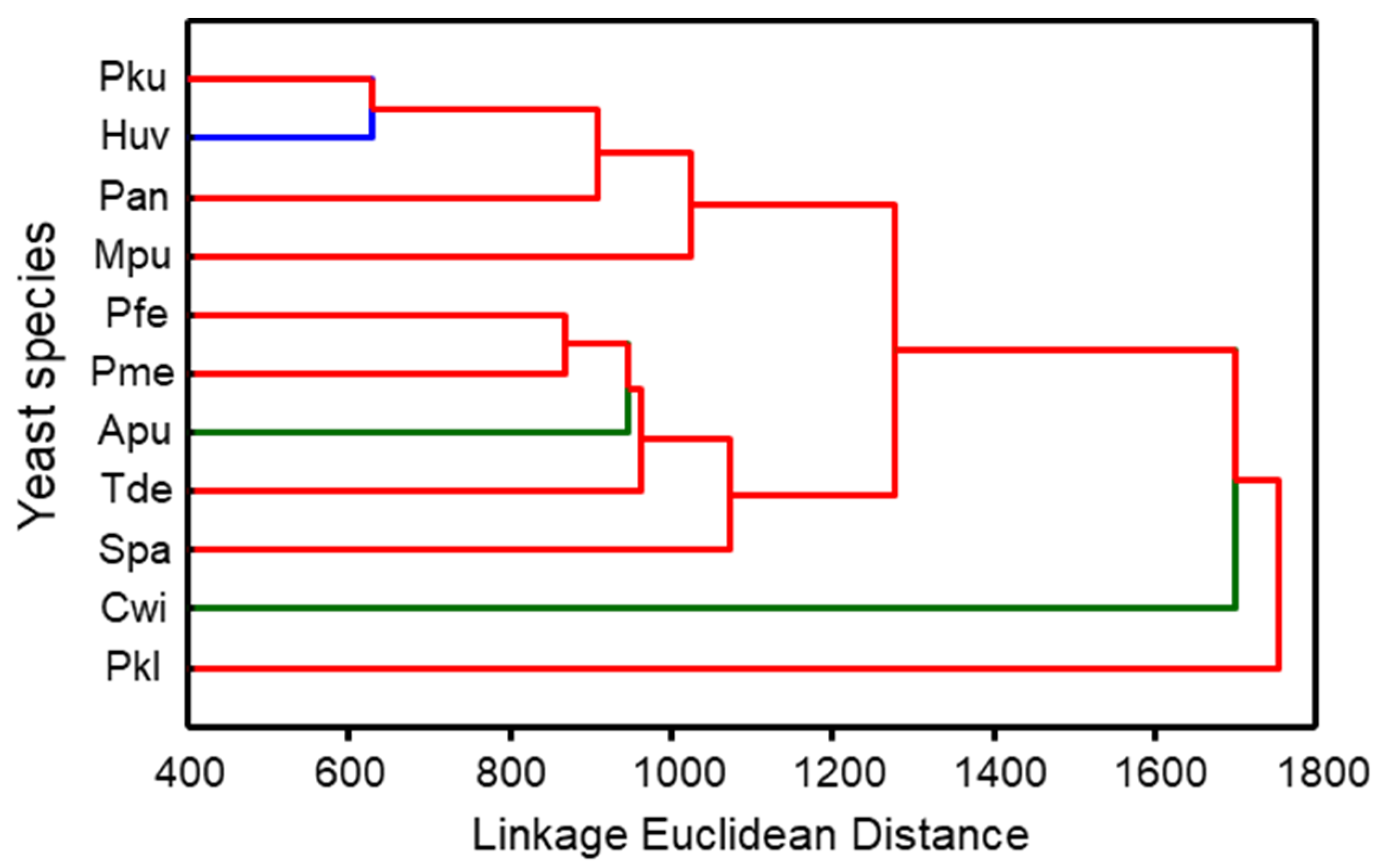
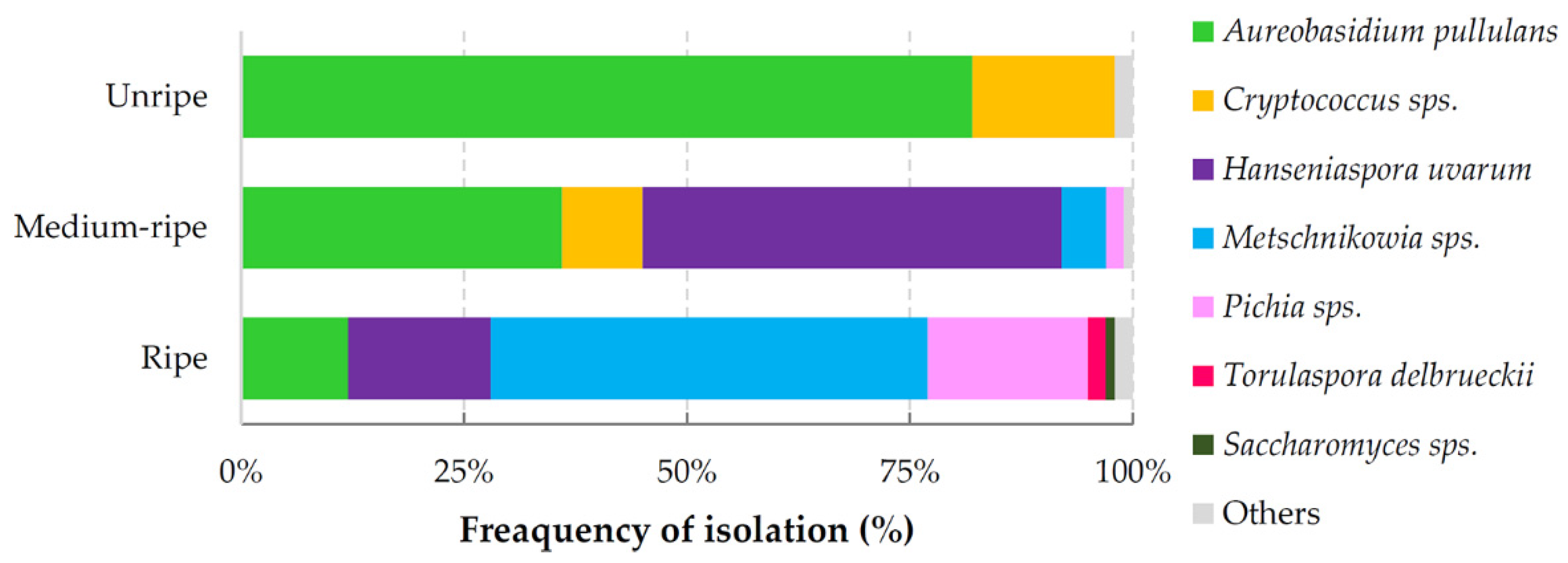
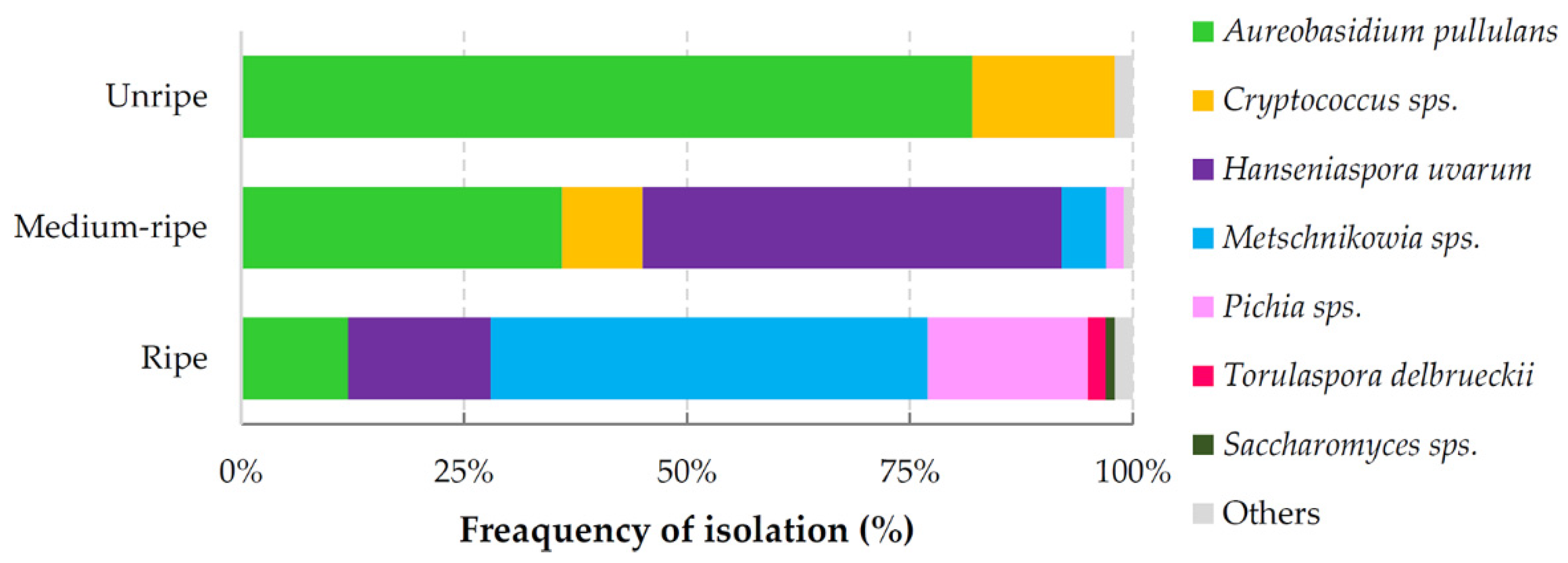
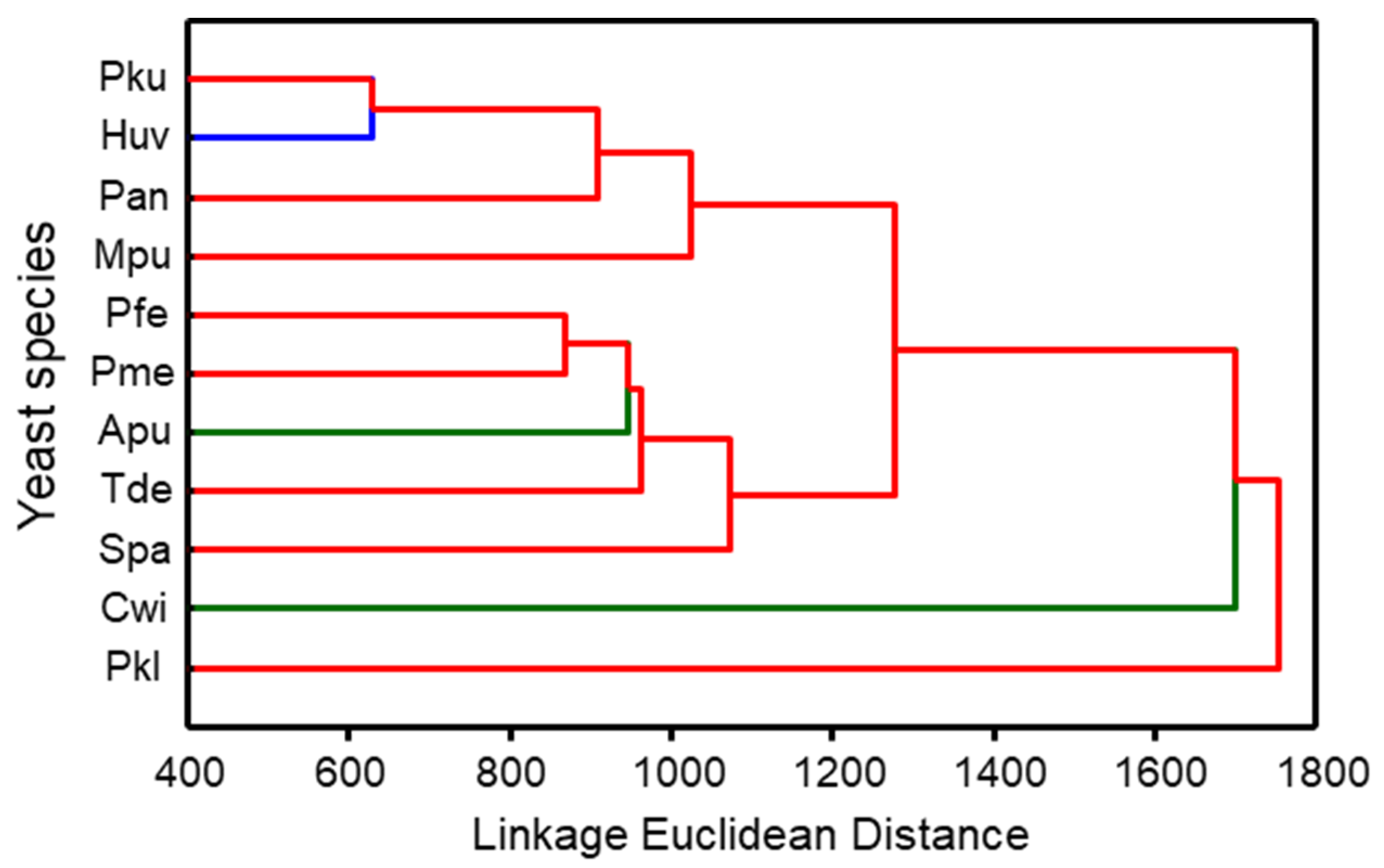
3.1. Composition of Yeast Produced Volatile Blends
3.2. EAD Active Compounds
3.3. EAG Dose–Response
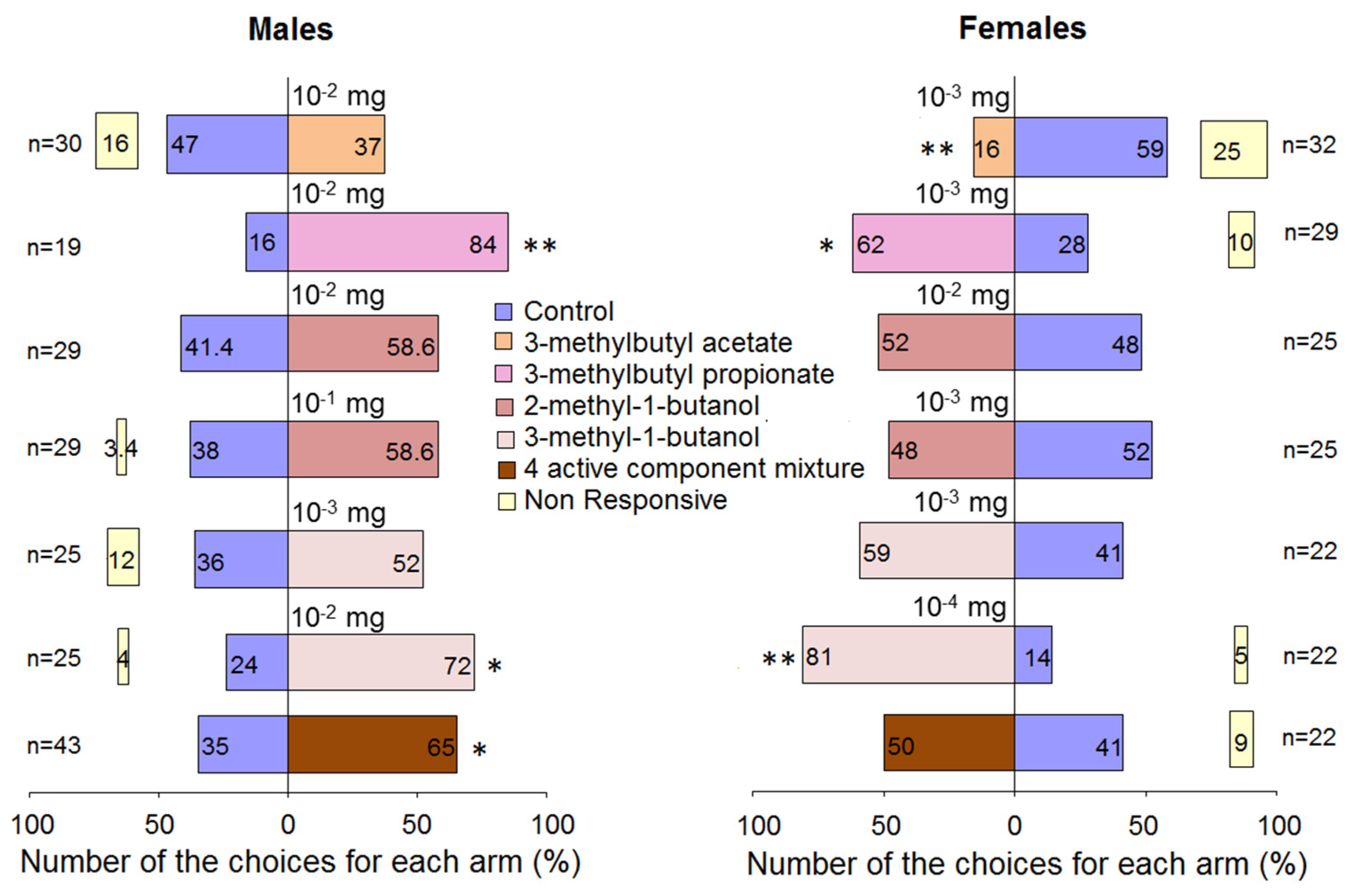
3.4. Behavioural Tests in Olfactometer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Species | Strain | GenBank Reference | Identity (%) |
|---|---|---|---|
| Aureobasidium pullulans | PA-4-13 | MN400109 | 100 |
| PC-5-28 | HQ909088 | 99.6 | |
| PC-4-8 | MW361317 | 99.8 | |
| Cryptococcus wieringae | PC-4-6 | KF981864 | 100 |
| PC-4-9 | KF981864 | 100 | |
| PA-5-41 | KF981864 | 100 | |
| Metschnikowia pulcherrima | PC-4-5 | MF574308 | 100 |
| PA-5-13 | KT029787 | 99.7 | |
| PA-5-47 | MK352050 | 100 | |
| Hanseniaspora uvarum | PA-5-27 | MF062209 | 100 |
| PC-3-8 | MK352020 | 99.9 | |
| PC-5-12 | KY103573 | 100 | |
| Pichia kudriavzevii | PA-1-15 | MF685423 | 100 |
| PC-4-25 | MF685411 | 100 | |
| PC-4-19.3 | MG015972 | 99.6 | |
| Pichia fermentans | PC-5-47 | FJ713081 | 100 |
| PC-4-19.1 | MF462777 | 100 | |
| PA-4-39 | KY104537 | 100 | |
| Pichia kluyveri | PA-4-30 | JX103190 | 100 |
| PA-4-34 | KY108823 | 99.8 | |
| PC-4-35 | KC510043 | 100 | |
| Pichia membranifaciens | PC-3-36 | JX188207 | 100 |
| PC-2-71 | FJ231461 | 100 | |
| PA-2-55 | JX188207 | 99.6 | |
| Saccharomyces paradoxus | PA-5-14 | FJ713072 | 100 |
| PC-3-33 | FJ713072 | 100 | |
| PC-3-59 | KY105204 | 99.9 | |
| Torulaspora delbrueckii | PA-4-16 | KY105641 | 100 |
| PA-5-17 | MN371902 | 99.8 | |
| PC-2-42 | MK352012 | 100 | |
| Pichia anomala | PC-5-5 | KJ527050 | 100 |
| PA-5-18 | MH248067 | 100 | |
| PC-5-24 | MK343437 | 100 |
References
- Starmer, W.T.; Lachance, M.-A. Chapter 6-Yeast Ecology. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 65–83. [Google Scholar] [CrossRef]
- Allard, S.M.; Ottesen, A.R.; Brown, E.W.; Micallef, S.A. Insect exclusion limits variation in bacterial microbiomes of tomato flowers and fruit. J. Appl. Microbiol. 2018, 125, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Gobbi, A.; Santoni, S.; Hansen, L.H.; This, P.; Peros, J.P. Assessing the impact of plant genetic diversity in shaping the microbial community structure of Vitis vinifera phyllosphere in the Mediterranean. Front. Life Sci. 2018, 11, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Vepstaite-Monstavic, I.; Luksa, J.; Staneviclene, R.; Strazdaite-Zieliene, Z.; Yurchenko, V.; Serva, S.; Serviene, E. Distribution of apple and blackcurrant microbiota in Lithuania and the Czech Republic. Microbiol. Res. 2018, 206, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kioroglou, D.; Kraeva-Deloire, E.; Schmidtke, L.M.; Mas, A.; Portillo, M.C. Geographical origin has a greater impact on grape berry fungal community than grape variety and maturation state. Microorganisms 2019, 7, 669. [Google Scholar] [CrossRef] [Green Version]
- Luksa, J.; Vepstaite-Monstavice, I.; Apsegaite, V.; Blazyte-Cereskiene, L.; Staneviciene, R.; Strazdaite-Zieliene, Z.; Ravoityte, B.; Aleknavicius, D.; Buda, V.; Mozuraitis, R.; et al. Fungal microbiota of sea buckthorn berries at two ripening stages and volatile profiling of potential biocontrol yeasts. Microorganisms 2020, 8, 456. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, M. Made for each other: Ascomycete yeasts and insects. Microbiol. Spectr. 2017, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Janson, E.M.; Stireman, J.O.; Singer, M.S.; Abbot, P. Phytophagous insect-microbe mutualisms and adaptive evolutionary diversification. Evolution 2008, 62, 997–1012. [Google Scholar] [CrossRef] [Green Version]
- Dobzhansky, T.; Cooper, D.M.; Phaff, H.J.; Knapp, E.P.; Carson, H.L. Studies on the ecology of Drosophila in the Yosemite region of California Differential attraction of species of Drosophila to different species of yeasts. Ecology 1956, 37, 544–550. [Google Scholar] [CrossRef]
- Date, P.; Dweck, H.K.M.; Stensmyr, M.C.; Shann, J.; Hansson, B.S.; Rollmann, S.M. Divergence in olfactory host plant preference in D-mojavensis in response to cactus host use. PLoS ONE 2013, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Scheidler, N.H.; Liu, C.; Hamby, K.A.; Zalom, F.G.; Syed, Z. Volatile codes: Correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [Green Version]
- Holighaus, G.; Rohlfs, M. Fungal allelochemicals in insect pest management. Appl. Microbiol. Biotechnol. 2016, 100, 5681–5689. [Google Scholar] [CrossRef]
- Staneviciene, R.; Luksa, J.; Strazdaite-Zieliene, Z.; Ravoityte, B.; Losinska-Siciuniene, R.; Mozuraitis, R.; Serviene, E. Mycobiota in the carposphere of sour and sweet cherries and antagonistic features of potential biocontrol yeasts. Microorganisms 2021, 9, 1423. [Google Scholar] [CrossRef] [PubMed]
- Buda, V.; Blazyte-Cereskiene, L.; Radziute, S.; Apsegaite, V.; Stamm, P.; Schulz, S.; Aleknavicius, D.; Mozuraitis, R. Male-produced (-)-delta-heptalactone, pheromone of fruit fly Rhagoletis batava (Diptera: Tephritidae), a sea buckthorn berries pest. Insects 2020, 11, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, D.J. Bioassay methods with terrestrial invertebrates. In Methods in Chemical Ecology: Bioassay Methods; Millar, J.G., Haynes, K.F., Eds.; Kluwer Academic Publishers: Norwell, MA, USA, 2000; pp. 212–270. [Google Scholar]
- Lemfack, M.C.; Nickel, J.; Dunkel, M.; Preissner, R.; Piechulla, B. mVOC: A database of microbial volatiles. Nucleic Acids Res. 2014, 42, D744–D748. [Google Scholar] [CrossRef] [PubMed]
- Ebert, B.E.; Halbfeld, C.; Blank, L.M. Exploration and Exploitation of the Yeast Volatilome. Curr. Metab. 2017, 5, 102–118. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [Green Version]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpedowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piskur, J. Chemical signaling and insect attraction is a conserved trait in yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Haustein, M.; Molina, F.; Petri, A.; Scholz, B.; Piechulla, B. Bacterial volatiles and their action potential. Appl. Microbiol. Biotechnol. 2009, 81, 1001–1012. [Google Scholar] [CrossRef]
- Mozuraitis, R.; Aleknavicius, D.; Vepstaite-Monstavice, I.; Staneviciene, R.; Emami, S.N.; Apsegaite, V.; Radziute, S.; Blazyte-Cereskiene, L.; Serviene, E.; Buda, V. Hippophae rhamnoides berry related Pichia kudriavzevii yeast volatiles modify behaviour of Rhagoletis batava flies. J. Adv. Res. 2020, 21, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Vitanovic, E.; Aldrich, J.R.; Boundy-Mills, K.; Cagalj, M.; Ebeler, S.E.; Burrack, H.; Zalom, F.G. Olive Fruit Fly, Bactrocera oleae (Diptera: Tephritidae), attraction to volatile compounds produced by host and insect-associated yeast strains. J. Econ. Entomol. 2020, 113, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.S.; Landolt, P.J. A survey of insect assemblages responding to volatiles from a ubiquitous fungus in an agricultural landscape. J. Chem. Ecol. 2013, 39, 860–868. [Google Scholar] [CrossRef]
- Daniel, C.; Grunder, J. Integrated management of European cherry fruit fly Rhagoletis cerasi (L.): Situation in Switzerland and Europe. Insects 2012, 3, 956–988. [Google Scholar] [CrossRef] [Green Version]
- Ghanbarzadeh, B.; Sampiao, J.P.; Arzanlou, M. Grape maturity significantly influences yeast community on grape berries: Basidiomycetous yeasts are dominant colonizers of immature grape berries in northwestern Iran. Nova Hedwig. 2021, 113, 191–206. [Google Scholar] [CrossRef]
- Revadi, S.; Vitagliano, S.; Stacconi, M.V.R.; Ramasamy, S.; Mansourian, S.; Carlin, S.; Vrhovsek, U.; Becher, P.G.; Mazzoni, V.; Rota-Stabelli, O.; et al. Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Cha, D.H.; Gill, M.A.; Epsky, N.D.; Werle, C.T.; Adamczyk, J.J.; Landolt, P.J. From a non-target to a target: Identification of a fermentation volatile blend attractive to Zaprionus indianus. J. Appl. Entomol. 2015, 139, 114–122. [Google Scholar] [CrossRef]
- Katsoyannos, B.I.; Papadopoulos, N.T.; Stavridis, D. Evaluation of trap types and food attractants for Rhagoletis cerasi (Diptera: Tephritidae). J. Econ. Entomol. 2000, 93, 1005–1010. [Google Scholar] [CrossRef]
- Biasazin, T.D.; Chernet, H.T.; Herrera, S.L.; Bengtsson, M.; Karlsson, M.F.; Lemmen-Lechelt, J.K.; Dekker, T. Detection of volatile constituents from food lures by Tephritid fruit flies. Insects 2018, 9, 119. [Google Scholar] [CrossRef] [Green Version]
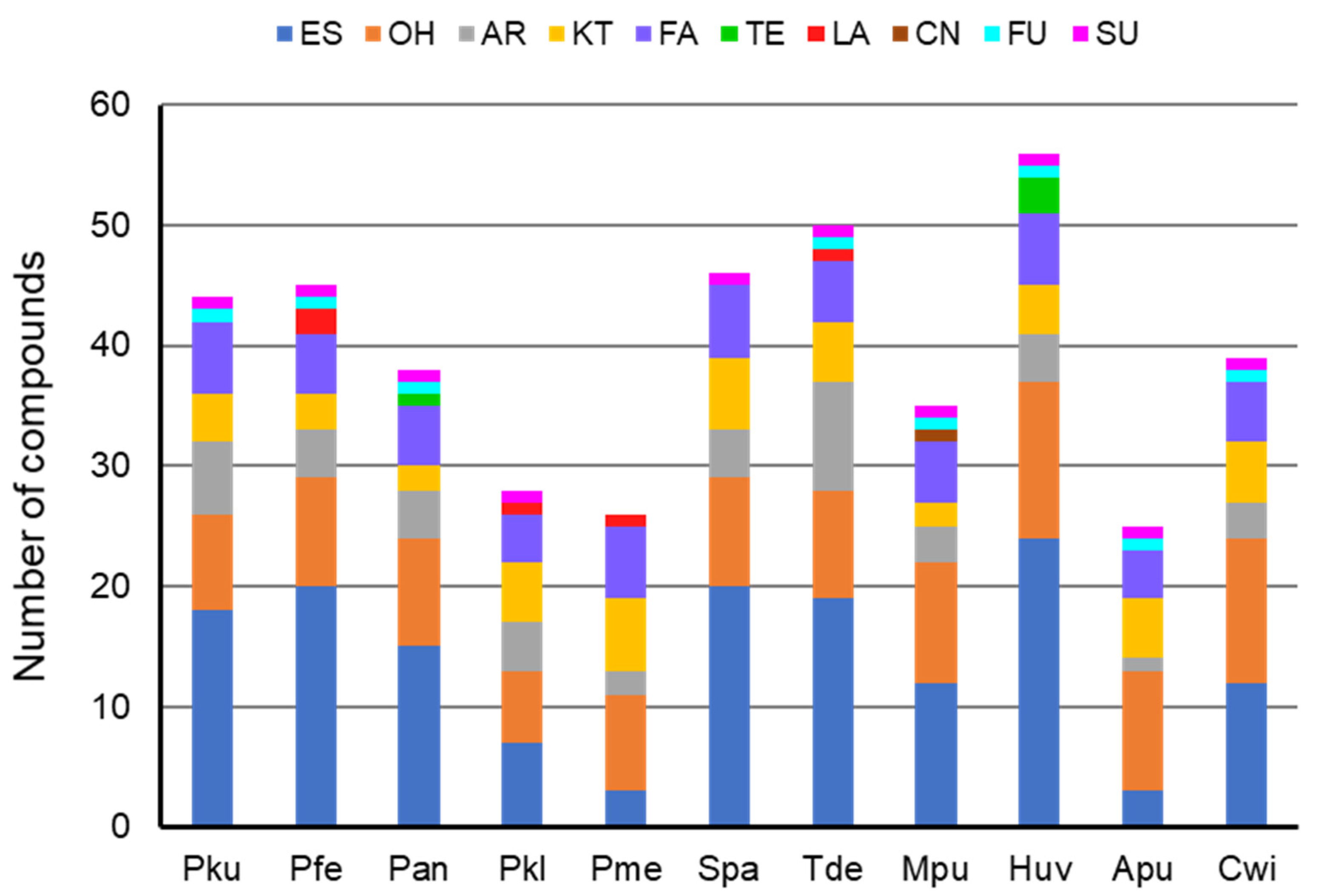




| No | Compound | CAS No 3 | RI 4 | GR 5 | Control | A. pullulans | C. wieringae | H. uvarum | P. kudriavzevii | P. fermentans | P. anomala | P. kluyveri | P. membranifac | S. paradoxus | T. delbrueckii | M. pulcherrima |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Ethyl acetate | 141-78-6 | >900 | ES 6 | 11 19 ± 6 | 0 | 0 | 2256 ± 657 | 2092 ± 150 | 1.6 ± 1.2 | 2816 ± 116 | 0 | 0 | 118 ± 76 | 202 ± 55 | 1586 ± 765 |
| 2 | 3-Methylbutanal * 1 | 590-86-3 | 927 | AL 7 | 38 ± 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 15 ± 8 | 0 | 0 |
| 3 | Ethanol | 64-17-5 | 944 | OH 8 | 0 | 707 ± 106 | 48 ± 11 | 386 ± 34 | 425 ± 5 | 152 ± 23 | 39 ± 23 | 0 | 0 | 2594 ± 1683 | 468 ± 19 | 677 ± 44 |
| 4 | Ethyl propionate | 1105-37-3 | 953 | ES | 0 | 0 | 0.9 ± 0.3 | 139 ± 38 | 149 ± 9 | 0 | 147 ± 14 | 0 | 0 | 1.00 ± 0.05 | 63 ± 8 | 65 ± 25 |
| 5 | Ethyl 2-methylpropionate | 97-62-1 | 960 | ES | 0 | 0 | 0 | 0 | 0 | 0.9 ± 0.7 | 0 | 0 | 0 | 0 | 0 | 0 |
| 6 | Propyl acetate | 109-60-4 | 969 | ES | 0 | 0 | 0 | 8.0 ± 0.7 | 0 | 0 | 99 ± 11 | 0 | 0 | 0 | 0 | 6.3 ± 3.6 |
| 7 | Methyl 2-methylbutanoate | 868-57-5 | 989 | ES | 0 | 0 | 0.8 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 8 | Methyl 3-methylbutanoate | 556-24-1 | 996 | ES | 0 | 0 | 0.7 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 9 | 2-Methylprop-1-yl acetate | 110-19-0 | 994 | ES | 0 | 0 | 0 | 15 ± 5 | 91 ± 6 | 0 | 59 ± 19 | 0.4 ± 0.4 | 0 | 5.4± 2.0 | 12 ± 3 | 8.9 ± 2.1 |
| 10 | Toluene * | 108-88-3 | 1013 | AR 9 | 6.3 ± 1.0 | 1.9 ± 0.1 | 1.7 ± 0.2 | 1.3 ± 0.6 | 0.8 ± 0.3 | 2.2 ± 0.3 | 0.5 ± 0.1 | 1.8 ± 0.8 | 3.8 ± 0.2 | 3.8 ± 1.7 ns | 2.2 ± 0.7 | 1.6 ± 0.1 |
| 11 | Ethyl butanoate | 105-54-4 | 1015 | ES | 0 | 0 | 0 | 0.8 ± 0.5 | 1.1 ± 0.1 | 0.5 ± 0.4 | 0.6 ± 0.1 | 0 | 0 | 8.8 ± 0.8 | 1.2 ± 0.3 | 0.5 ± 0.4 |
| 12 | Ethyl 2-methylbutanoate | 7452-79-1 | 1035 | ES | 0 | 3.6 ± 0.5 | 1.6 ± 0.3 | 1.3 ± 0.5 | 1.2 ± 0.2 | 4.3 ± 3.7 | 3.8 ± 0.9 | 0 | 0 | 6.6 ± 0.2 | 2.6 ± 0.2 | 3.5 ± 0.5 |
| 13 | 3-Methylbutyl formate | 110-45-2 | 1053 | ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.0 ± 0.4 | 0 | 0 | 0 | 0 |
| 14 | Ethyl 3-methylbutanoate | 108-64-5 | 1053 | ES | 0 | 0.4 ± 0.1 | 0.10 ± 0.02 | 0 | 0 | 3.9 ± 2.8 | 0 | 0 | 0 | 1.7 ± 0.4 | 0.7 ± 0.1 | 0.6 ± 0.2 |
| 15 | Butyl acetate | 123-86-4 | 1055 | ES | 0 | 0 | 0 | 12 ± 2 | 1.8 ± 0.2 | 0 | 46 ± 4 | 0 | 0 | 0 | 0.7 ± 0.4 | 0.15 ± 0.03 |
| 16 | 2-Methylpropyl propionate | 540-42-1 | 1064 | ES | 0 | 0 | 0.4 ± 0.1 | 1.1 ± 0.2 | 3.7 ± 0.5 | 1.7 ± 0.9 | 3.6 ± 0.8 | 496 ± 496 | 0 | 0.8 ± 0.1 | 3.5 ± 0.4 | 0.4 ± 0.2 |
| 17 | 2-Methylpropyl 2-methylpropionate | 97-85-8 | 1074 | ES | 0 | 0 | 0.2 ± 0.1 | 0 | 0 | 0.3 ± 0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 18 | 2-Methylpropanol | 78-83-1 | 1093 | OH | 0 | 114 ± 6 | 143 ± 12 | 53 ± 9 | 61 ± 7 | 106 ± 17 | 25 ± 1 | 56 ± 14 | 26 ± 03 | 97 ± 6 | 56 ± 06 | 102 ± 20 |
| 19 | 3-Methylbutyl acetate | 123-92-2 | 1109 | ES | 5.4 ± 0.2 | 0 | 4.3 ± 1.1 ns | 658 ± 88 | 1202 ± 167 | 117 ± 84 | 1934 ± 948 | 70 ± 50 | 0 | 188 ± 55 | 623 ± 252 | 49 ± 19 |
| 20 | 1-Butanol * | 71-36-3 | 1144 | OH | 0.8 ± 0.03 | 2.9 ± 0.6 | 18 ± 5 | 4.4 ± 0.6 | 0.3 ± 0.2 | 0.4 ± 0.4 ns | 1.6 ± 0.6 | 0 | 0 | 2.2 ± 0.3 | 0.4 ± 0.2 ns | 4.5 ± 1.0 |
| 21 | 2-Methylpropyl 2-methylbutanoate | 2445-67-2 | 1166 | ES | 0 | 0 | 0 | 0 | 0 | 0.7 ± 0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22 | 2-Heptanone | 110-43-0 | 1171 | KT 10 | 0 | 10 ± 8 | 8.0 ± 1.7 | 0.5 ± 0.1 | 0 | 2.6 ± 1.1 | 0.2 ± 0.04 | 1.9 ± 0.9 | 3.3 ± 0.6 | 2.6 ± 0.6 | 4.8 ± 0.6 | 0.4 ± 0.4 |
| 23 | 3-Methylbutyl propionate | 105-68-0 | 1178 | ES | 0.4 ± 0.02 | 0 | 0.3 ± 0.2 ns | 84 ± 24 | 23 ± 4 | 20 ± 18 | 71 ± 8 | 2937 ± 1962 | 1.6 ± 0.7 | 11 ± 1 | 98 ± 11 | 1.4 ± 0.5 |
| 24 | 3-Methylbutyl 2-methylpropionate | 2050-01-3 | 1184 | ES | 0 | 0 | 0 | 0 | 0 | 4.5 ± 1.8 | 0 | 0 | 0.5 ± 0.2 | 0.3 ± 0.1 | 12 ± 3 | 0 |
| 25 | 2-Methyl-1-butanol | 137-32-6 | 1205 | OH | 0 | 80 ± 11 | 262 ± 64 | 129 ± 25 | 201 ± 61 | 209 ± 19 | 96 ± 11 | 154 ± 15 | 29 ± 4 | 172 ± 53 | 199 ± 37 | 193 ± 4 |
| 26 | 3-Methyl-1-butanol | 123-51-3 | 1212 | OH | 0.8 ± 0.1 | 640 ± 86 | 889 ± 297 | 461 ± 84 | 874 ± 74 | 1876 ± 117 | 416 ± 21 | 568 ± 118 | 687 ± 169 | 1418 ± 93 | 1787 ± 61 | 644 ± 226 |
| 27 | Ethyl hexanoate | 123-66-0 | 1225 | ES | 0 | 0.8 ± 0.4 | 0 | 0.6 ± 0.1 | 0.7 ± 0.2 | 3.6 ± 3.6 | 0 | 0 | 0 | 16 ± 9 | 0.1 ± 0.03 | 0 |
| 28 | Styrene * | 100-42-5 | 1237 | AR | 11 ± 4 | 2.4 ± 2.0 | 19 ± 5 ns | 0.5 ± 0.2 | 3.5 ± 2.9 ns 20 | 5.2 ± 2.6 ns | 19 ± 24 ns | 14 ± 0.7 ns | 7.2 ± 2.1 ns | 9 ± 2 ns | 10 ± 3 ns | 11 ± 6 ns |
| 29 | 3-Methylbutyl butanoate | 106-27-4 | 1256 | ES | 0 | 0 | 0 | 0.4 ± 0.1 | 0 | 0.5 ± 0.02 | 0 | 0 | 0 | 0 | 0.4 ± 0.1 | 0 |
| 30 | Hexyl acetate | 142-92-7 | 1265 | ES | 0 | 0 | 0 | 0.45 ± 0.03 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 31 | 3-Methylbutyl 2-methylbutanoate | 27625-35-0 | 1272 | ES | 0 | 0 | 0 | 0 | 0 | 3.0 ± 0.8 | 1.5 ± 0.3 | 0 | 0 | 0 | 0 | 0 |
| 32 | 2-hydroxy-3-butanone | 513-86-0 | 1274 | KT | 0 | 2.2 ± 0.9 | 3.6 ± 0.4 | 10 ± 1 | 0 | 0 | 0 | 0.7 ± 0.1 | 0 | 6.7±0.7 | 2.2 ± 0.2 | 17 ± 2 |
| 33 | 3-Methylbutyl 3-methylbutanoate | 659-70-1 | 1288 | ES | 0 | 0 | 0.4 ± 0.2 | 0 | 0 | 8.9 ± 3.9 | 0.7 ± 0.1 | 0 | 0 | 0 | 0 | 0 |
| 34 | 2-Methylpentanol | 105-30-6 | 1299 | OH | 0 | 0 | 13 ± 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 35 | 2-Methylpropyl isothiocyanoate | 591-82-2 | 1304 | CN 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.4 ± 0.1 |
| 36 | 2,5- Dimethyl pyrazine * | 123-32-0 | 1314 | PY 12 | 9.1 ± 0.5 | 1.2 ± 0.2 | 22 ± 13 ns | 3.8 ± 0.6 | 3.4 ± 0.4 | 4.8 ± 0.2 | 2.0 ± 0.6 | 6.0 ± 3.1 ns | 7.6 ± 0.4 ns | 6.4±0.5 ns | 3.9 ± 0.5 | 4.6 ±0.6 |
| 37 | 2-Heptanol | 543-49-7 | 1321 | OH | 0 | 2.0 ± 1.3 | 0 | 0.50 ± 0.01 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.9 ± 0.7 |
| 38 | 6-Methyl 5-hepten-2-one | 110-93-0 | 1327 | KT | 0.4 ± 0.1 | 4.4 ± 3.8 | 103 ± 23 | 0.29 ± 0.03 ns | 0 | 0.7 ± 0.1 | 0 | 0.8 ± 0.5 ns | 2.8 ± 1.0 | 1.1 ± 0.2 | 0.5 ± 0.4 ns | 0 |
| 39 | Ethyl heptanoate | 106-30-9 | 1331 | ES | 0 | 0 | 0 | 0 | 0.4 ± 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 40 | 1-Hexanol | 111-27-3 | 1346 | OH | 0 | 0 | 0.5 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 4.3 ± 1.9 | 0 | 0 | 0 |
| 41 | Heptyl acetate | 112-06-1 | 1369 | ES | 0 | 0 | 0 | 0.7 ± 0.1 | 0 | 0 | 0 | 0 | 0 | 0.3 ± 0.2 | 0 | 0 |
| 42 | 2-Ethylhexyl acetate | 103-09-3 | 1382 | ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.0 ± 0.4 | 0 | 0 |
| 43 | Nonan-2-one | 821-55-6 | 1383 | KT | 0 | 49 ± 28 | 13 ± 4 | 0.5 ± 0.2 | 0.3 ± 0.1 | 2.3 ± 1.0 | 1.7 ± 0.4 | 1.3 ± 0.2 | 2.9 ± 1.0 | 5.2 ± 4.2 | 2.8 ± 0.6 | 0 |
| 44 | 2,3,5-Trimethyl pyrazine * | 14667-55-1 | 1397 | PY | 0.6 ± 0.1 | 0.6 ± 0.1 ns | 0.8 ± 0.1 ns | 0.58 ± 0.01 ns | 0.6 ± 0.04 ns | 0.6 ± 0.1 ns | 0.6 ± 0.1 ns | 0.4 ± 0.2 ns | 0.7 ± 0.2 ns | 0.8 ± 0.1 ns | 0.5 ± 0.4 ns | 0.60 ± 0.02 ns |
| 45 | Ethyl octanoate | 106-32-1 | 1427 | ES | 0 | 0 | 0 | 4.1 ± 0.1 | 3.7 ± 0.6 | 0.2 ± 0.2 | 0 | 0 | 0 | 5.6 ± 5.2 | 0 | 0 |
| 46 | Acetic acid | 64-19-7 | 1439 | FA 13 | 0.3 ± 0.01 | 0.7 ± 0.01 | 4.5 ± 3.5 ns | 69 ± 11 | 5.3 ± 3.9 ns | 0.6 ± 0.1 | 7.9 ± 3.0 | 0 | 0.8 ± 0.1 | 0.79 ± 0.04 | 0.77 ± 0.04 | 1.1 ±0.4 |
| 47 | Unknown | 1440 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 ± 4 | 0 | 0 | 0 | 0 | ||
| 48 | 1-Octen-3-ol | 3391-86-4 | 1450 | OH | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.12 ± 0.01 | 0 | 0 | 0 |
| 49 | 1-Heptanol | 111-70-6 | 1454 | OH | 0 | 0 | 0 | 0.8 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 50 | 3-Methylbutyl hexanoate | 2198-61-0 | 1457 | ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.25 ± 0.01 | 0.8 ± 0.2 | 0 | 0 |
| 51 | 6-Methyl-5-hepten-2-ol | 1569-60-4 | 1460 | OH | 0 | 1.1 ± 0.7 | 12 ± 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 52 | MHMP 2 | 40348-72-9 | 1462 | ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.2 ± 0.3 | 0 |
| 53 | Octyl acetate | 112-14-1 | 1472 | ES | 0 | 0 | 0 | 0.3 ± 0.1 | 0 | 0 | 0 | 0 | 0 | 1.3 ± 0.9 | 0 | 0 |
| 54 | 2-Ethylhexanol | 104-76-7 | 1483 | OH | 0.3 ± 0.03 | 1.5 ± 0.1 | 1.8 ± 0.2 | 0.8 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 | 1.3 ± 0.4 | 0.8 ± 0.4 ns | 1.7 ± 0.8 | 0.9 ± 0.1 | 1.1 ± 0.2 | 0.71 ± 0.03 |
| 55 | Acetylfuran | 1192-62-7 | 1496 | FU 14 | 0 | 0.3 ± 0.1 | 0.3 ± 0.2 | 0.6 ± 0.1 | 0.2 ± 0.1 | 0.6 ± 0.3 | 0.7 ± 0.3 | 0 | 0 | 0 | 0.5 ± 0.1 | 0.19 ± 0.02 |
| 56 | Benzaldehyde * | 100-52-7 | 1505 | AR, AL | 9.9 ± 2.7 | 2.0 ± 1.6 | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.28 ± 0.04 | 0.29 ± 0.04 | 0.18 ± 0.02 | 1.0 ± 0.2 | 0.20 ± 0.02 | 0.23 ± 0.01 | 0.20 ± 0.03 | 0.3 ± 0.1 |
| 57 | 2-Nonanol | 628-99-9 | 1524 | OH | 0 | 0.05 ± 0.04 | 0 | 1.1 ± 0.2 | 0 | 0.2 ± 0.1 | 0 | 0 | 0 | 0.3 ± 0.1 | 0.4 ± 0.1 | 0.5 ± 0.1 |
| 58 | Propionic acid | 79-09-4 | 1526 | FA | 0.3 ± 0.04 | 0 | 2.9 ± 2.0 ns | 6.7 ± 1.7 | 2.2 ± 0.1 | 0.7 ± 0.5 ns | 1.0 ± 0.4 | 0.9 ± 0.2 | 1.6 ± 0.3 | 2.3 ± 0.4 | 2.4 ± 0.5 | 0.3 ± 0.1 ns |
| 59 | 2-Methylpropionic acid | 79-31-2 | 1557 | FA | 4.9 ± 0.5 | 0 | 12 ± 6 ns | 34 ± 5 | 26 ± 2 | 13 ± 7 | 9.9 ± 2.2 | 8.3 ± 1.6 | 20 ± 3 | 1.8 ± 1.2 ns | 1.9 ± 0.8 ns | 1.9 ± 1.0 ns |
| 60 | 2-Decanol | 1120-06-5 | 1574 | OH | 0 | 0 | 0 | 0.7 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 61 | 2-Undecanone | 112-12-9 | 1594 | KT | 0 | 1.1 ± 0.6 | 0.6 ± 0.1 | 0 | 0 | 0 | 0 | 0.2 ± 0.1 | 1.1 ± 0.2 | 0.20 ± 0.02 | 0.8 ± 0.2 | 0 |
| 62 | Butanoic acid | 107-92-6 | 1614 | FA | 1.0 ± 0.1 | 0.2 ± 0.04 | 2.6 ± 1.8 ns | 8.5 ± 1.7 | 2.7 ± 0.1 | 0.7 ± 0.5 ns | 9.0 ± 2.1 | 0.3 ± 0.2 | 1.1 ± 0.1 ns | 0.2 ± 0.1 | 0.21 ± 0.04 | 0.3 ± 0.2 |
| 63 | Acetophenone * | 98-86-2 | 1625 | AR, KT | 1.8 ± 0.2 | 0.7 ± 0.1 | 0.7 ± 0.1 | 1.1 ± 0.1 | 0.8 ± 0.04 | 0.4 ± 0.4 | 0.9 ± 0.1 | 1.1 ± 0.8 ns | 1.3 ± 0.1 ns | 1.6 ± 1.0 ns | 1.5 ± 0.1 ns | 0.6 ± 0.1 |
| 64 | Ethyl decanoate | 110-38-3 | 1632 | ES | 0 | 0 | 0 | 4.0 ± 1.2 | 0.9 ± 0.1 | 0.5 ± 0.5 | 0.2 ± 0.1 | 0 | 0 | 2.3 ± 2.2 | 0 | 0 |
| 65 | 2-Furanmethanol * | 98-00-0 | 1649 | OH | 1.8 ± 0.1 | 1.7 ± 1.5 ns | 1.2 ± 0.4 ns | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.7 ± 0.4 | 0 |
| 66 | 3-Methylbutanoic acid | 503-74-2 | 1658 | FA | 20 ± 2 | 0.26 ± 0.02 | 58 ± 39 | 261 ± 66 | 136 ± 9 | 37 ± 25 ns | 44 ± 12 | 21 ± 6 ns | 71 ± 14 | 6.8 ± 5.6 | 4.5 ± 2.9 | 4.6 ± 4.1 |
| 67 | 3-Hydroxypropyl methylsulphide | 505-10-2 | 1700 | SU 15 | 0 | 0.18 ± 0.03 | 0.4 ± 0.1 | 0.33 ± 0.04 | 2.0 ± 1.0 | 1.3 ± 0.7 | 6.6 ± 1.1 | 0.9 ± 0.4 | 0 | 0.8 ± 0.2 | 7.5 ± 2.2 | 0.5 ± 0.2 |
| 68 | Geranyl acetate | 16409-44-2 | 1764 | TE 16, ES | 0 | 0 | 0 | 0.7 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 69 | Methoxy-phenyl-oxime * | 67160-14-9 | 1768 | IM 17 | 3.1 ± 0.7 | 1.2 ± 0.2 | 2.9 ± 0.4 ns | 0.6 ± 0.3 | 1.3 ± 0.1 | 0.7 ± 0.2 | 0.43 ± 0.02 | 1.7 ± 0.4 ns | 1.0 ± 0.3 | 0.6 ± 0.2 | 0.6 ± 0.2 | 1.4 ± 0.4 |
| 70 | Ethyl 2-phenylacetate | 119-43-7 | 1767 | AR, ES | 0 | 0 | 0 | 0 | 0.6 ± 0.2 | 0 | 0.3 ± 0.1 | 0 | 0 | 0 | 0 | 0 |
| 71 | 2-Phenylethyl acetate | 103-45-7 | 1794 | AR, ES | 0 | 0 | 0 | 148 ± 13 | 353 ± 48 | 2.1 ± 1.6 | 88 ± 8 | 7.8 ± 6.3 | 0 | 8.0 ± 3.4 | 65 ± 17 | 11 ± 8 |
| 72 | Hexanoic acid | 122-70-3 | 1816 | AR, ES | 0 | 0 | 0.8 ± 0.1 | 0 | 0.4 ± 0.1 | 0.3 ± 0.2 | 0 | 0 | 0 | 0.6 ± 0.1 | 0 | 0 |
| 73 | Ethyl dodecanoate | 106-33-2 | 1823 | ES | 0 | 0 | 0 | 1.7 ± 1.0 | 0.1 ± 0.03 | 0.3 ± 0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 74 | Geraniol | 106-24-1 | 1842 | TEOH | 0 | 0 | 0.5 ± 0.1 | 0 | 0 | 0 | 0.5 ± 0.1 | 0 | 0 | 0 | 0 | 0 |
| 75 | 3-Methylbutyl decanoate | 2306-91-4 | 1843 | ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 ± 0.1 | 0 | 0 |
| 76 | Phenyl methanol | 100-51-6 | 1854 | AR, OH | 0.7 ± 0.04 | 0 | 0.7 ± 0.2 ns | 5.1 ± 0.9 | 2.9 ± 0.5 | 1.8 ± 1.0 ns | 1.2 ± 0.3 ns | 1.2 ± 0.2 | 0.6 ± 0.2 ns | 0.7 ± 0.2 ns | 1.1 ± 0.2 ns | 2.2 ± 0.3 |
| 77 | 2-Phenylethyl propionate | 122-70-3 | 1862 | AR, ES | 0 | 0 | 0 | 18 ± 4 | 6.3 ± 0.7 | 0 | 1.1 | 1.1 ± 0.9 | 0 | 0 | 172 ± 38 | 0 |
| 78 | 2-Phenyl ethanol | 60-12-8 | 1890 | AR, OH | 0.2 ± 0.04 | 362 ± 102 | 20 ± 2 | 336 ± 46 | 567 ± 19 | 865 ± 206 | 235 ± 17 | 432 ± 57 | 814 ± 17 | 271 ± 83 | 448 ± 87 | 490 ± 320 |
| 79 | 2-Phenylethyl butanoate | 103-52-6 | 1949 | AR, ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 ± 0.1 | 0 |
| 80 | 2-Phenylethyl 2-methylbutanoate | 24817-51-4 | 1955 | AR, ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4.3 ± 1.8 | 0 |
| 81 | 2-Phenylethyl 3-methylbutanoate | 140-26-1 | 1973 | AR, ES | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.11 ± 0.02 | 0 |
| 82 | Phenol * | 87-66-1 | 1979 | AR, OH | 0.4 ± 0.04 | 0.38 ± 0.01 ns | 0.5 ± 0.1 ns | 0.15 ± 0.01 | 0.07 ± 0.03 | 0.4 ± 0.1 ns | 0.5 ± 0.1 ns | 0.2 ± 0.1 ns | 0.6 ± 0.2 ns | 0.49 ± 0.02 | 0.3 ± 0.1 ns | 0.30 ± 0.04 ns |
| 83 | Pentadecan-2-one | 2345-28-0 | 2010 | KT | 0 | 0 | 0 | 0 | 0.2 ± 0.1 | 0 | 0 | 0 | 1.2 ± 0.3 | 0.08 ± 0.06 | 0 | 0 |
| 84 | Octanoic acid | 124-07-2 | 2065 | FA | 0.3 ± 0.02 | 1.0 ± 0.2 | 0 | 0.3 ± 0.1 ns | 0.6 ± 0.1 | 0 | 0 | 0 | 0.27 ± 0.03 ns | 0.20 ± 0.01 | 0 | 0 |
| 85 | 3-Methylbutyl dodecanoate | 6309-51-9 | 2089 | ES | 0 | 0 | 0 | 2.3 ± 0.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 86 | Hexadecan-2-one | 18787-63-8 | 2118 | KT | 0 | 0 | 0 | 0 | 0.4 ± 0.1 | 0 | 0 | 0 | 0.6 ± 0.2 | 0 | 0 | 0 |
| 87 | gamma-Decalactone | 706-14-9 | 2125 | LA 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.2 ± 0.2 | 5.2 ± 1.2 | 0 | 1.3 ± 0.8 | 0 |
| 88 | 6-Pentyl-5,6 dihydro-2H-pyran-2-one | 54814-64-1 | 2222 | LA | 0 | 0 | 0 | 0 | 0 | 2.3 ± 1.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 89 | Heptadecan-2-one | 2922-51-2 | 2230 | KT | 0 | 0 | 0 | 0 | 2.1 ± 0.6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 90 | Farnesyl acetate | 4128-17-0 | 2247 | TE, ES | 0 | 0 | 0 | 1.1 ± 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 91 | Farnesol | 4602-84-0 | 2287 | TE, OH | 0 | 0 | 0 | 1.7 ± 0.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 92 | 4-Hydroxy-6-pentyloxan-2-one | 36555-25-6 | 2456 | LA | 0 | 0 | 0 | 0 | 0 | 1.2 ± 0.4 | 0 | 0 | 0 | 0 | 0 | 0 |
| Yeast Species | 3-MBA 1 | 3-MBP 2 | 2-MBOH 3 | 3-MBOH 4 | ||||
|---|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | Male | Female | |
| Aureobasidium pullulans | - 5 | 3 (3) | 3 (3) | 3 (3) | 3 (3) | |||
| Cryptococcus wieringae | 4 (4) | 3 (3) | 4 (4) | 3 (3) | ||||
| Hanseniaspora uvarum | 4 6 (4) 7 | 4 (4) | 4 (4) | 3 (4) | 4 (4) | 4 (4) | 4 (4) | 4 (4) |
| Pichia kudriavzevii | 5 (7) | 4 (4) | 5 (7) | 4 (4) | 7 (7) | 4 (4) | 7 (7) | 4 (4) |
| Metschnikowia pulcherrima | 1 (3) | 3 (4) | 3 (3) | 4 (4) | 3 (3) | 4 (4) | ||
| Total | 10 (14) | 11 (12) | 9 (11) | 7 (8) | 21 (21) | 18 (18) | 21 (21) | 18 (18) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mozūraitis, R.; Apšegaitė, V.; Radžiutė, S.; Aleknavičius, D.; Būdienė, J.; Stanevičienė, R.; Blažytė-Čereškienė, L.; Servienė, E.; Būda, V. Volatiles Produced by Yeasts Related to Prunus avium and P. cerasus Fruits and Their Potentials to Modulate the Behaviour of the Pest Rhagoletis cerasi Fruit Flies. J. Fungi 2022, 8, 95. https://doi.org/10.3390/jof8020095
Mozūraitis R, Apšegaitė V, Radžiutė S, Aleknavičius D, Būdienė J, Stanevičienė R, Blažytė-Čereškienė L, Servienė E, Būda V. Volatiles Produced by Yeasts Related to Prunus avium and P. cerasus Fruits and Their Potentials to Modulate the Behaviour of the Pest Rhagoletis cerasi Fruit Flies. Journal of Fungi. 2022; 8(2):95. https://doi.org/10.3390/jof8020095
Chicago/Turabian StyleMozūraitis, Raimondas, Violeta Apšegaitė, Sandra Radžiutė, Dominykas Aleknavičius, Jurga Būdienė, Ramunė Stanevičienė, Laima Blažytė-Čereškienė, Elena Servienė, and Vincas Būda. 2022. "Volatiles Produced by Yeasts Related to Prunus avium and P. cerasus Fruits and Their Potentials to Modulate the Behaviour of the Pest Rhagoletis cerasi Fruit Flies" Journal of Fungi 8, no. 2: 95. https://doi.org/10.3390/jof8020095
APA StyleMozūraitis, R., Apšegaitė, V., Radžiutė, S., Aleknavičius, D., Būdienė, J., Stanevičienė, R., Blažytė-Čereškienė, L., Servienė, E., & Būda, V. (2022). Volatiles Produced by Yeasts Related to Prunus avium and P. cerasus Fruits and Their Potentials to Modulate the Behaviour of the Pest Rhagoletis cerasi Fruit Flies. Journal of Fungi, 8(2), 95. https://doi.org/10.3390/jof8020095