
Chaetomugilins and Chaetoviridins—Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics




Abstract
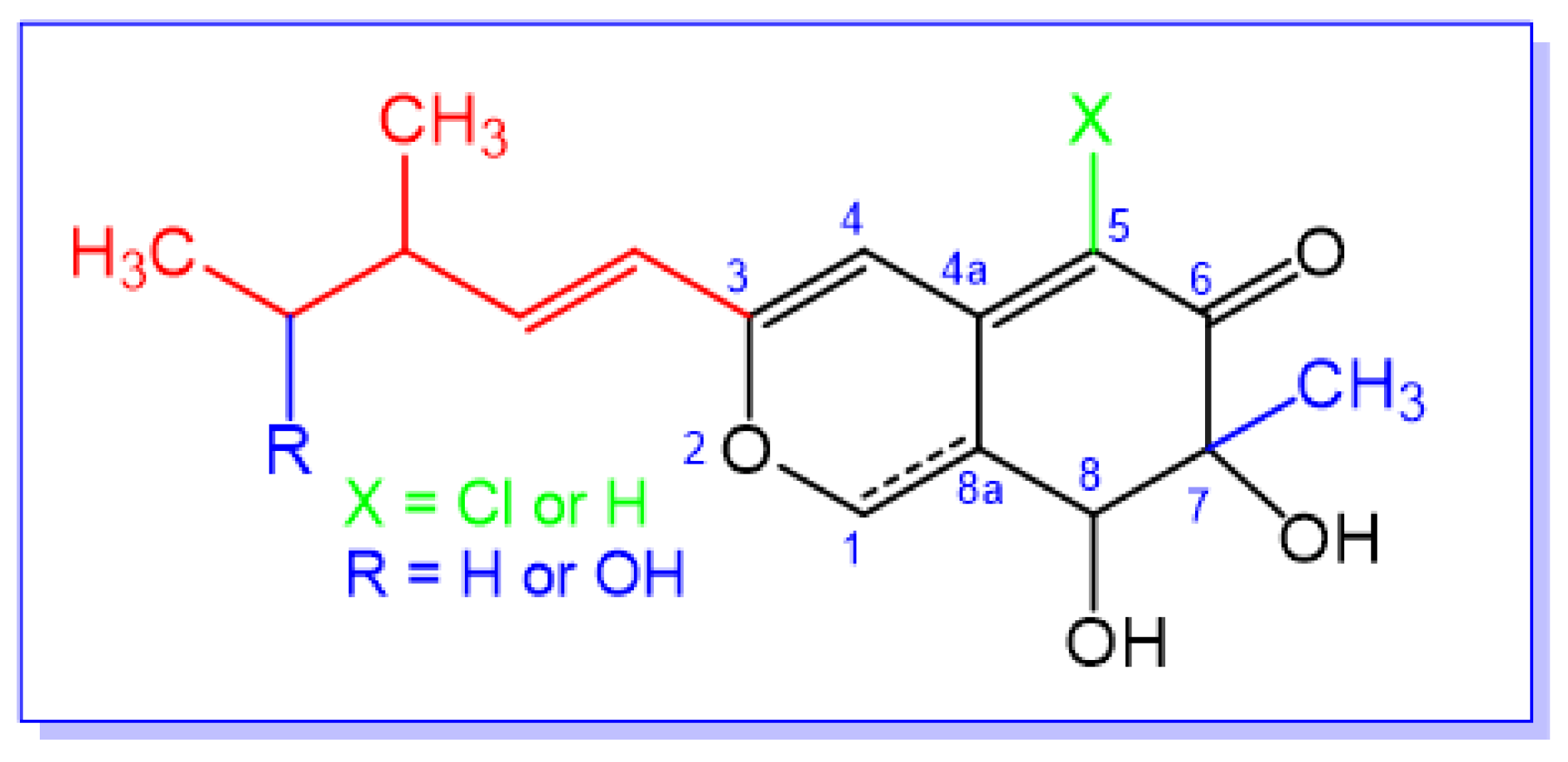
:1. Introduction
2. Extraction, Isolation, and Structural Characterization

| Compound Name | Mol. Wt. | Mol. Formula | Fungal Source | Host (Part, Family) | Place | Refs. |
|---|---|---|---|---|---|---|
| Chaetomugilin 106B-6 XXVIII (1) | 328 | C16H21ClO5 | C. globosum | Mugii cephalus (Fish bora stomach content, Mugilidae) | Katsuura, Nachi, Wakayama, Japan | [37] |
| Chaetomugilin A (2) | 450 | C23H27ClO7 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [38] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [40] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [41] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [35] | |||
| C. globosum | Ginkgo biloba (Leaves, Ginkgoaceae) | Linyi, Shandong, China | [42] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [43] | |||
| C. globosum Z1 | Broussonetia papyrifera (Barks, Moraceae) | Nanjing, Jiangsu, China | [44] | |||
| C. globosum TY1 | Ginkgo biloba (Barks Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| C. globosum CBS148.51 | Cultured | China | [46] | |||
| C. globosum HDN151398 | Sediment sea | South China Sea | [22] | |||
| C. globosum TY-2 | Polygonatum sibiricum, (Root, Convallariaceae) | Linan, Zhejiang, China | [32] | |||
| C. globosum TY1 | Ginkgo biloba (Barks Ginkgoaceae) | Linyi, Shandong, China | [47] | |||
| Seco-chaetomugilin A (3) | 482 | C24H31ClO8 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [48] |
| 11-Epi-chaetomugilin A (4) | 450 | C23H27ClO7 | C. globosum | Mugii cephalus (Fish bora stomach content, Mugilidae) | Katsuura, Nachi, Wakayama, Japan | [37] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [35] | |||
| C. globosum TW1-1 | Armadillidium vulgare (Pillbugs, Armadillidiidae) | Tongji Medical College, Hubei, China | [33] | |||
| C. globosum CBS148.51 | Culture | China | [46] | |||
| 4′-Epi-chaetomugilin A (5) | 450 | C23H27ClO7 | C. globosum | Mugii cephalus (Fish bora stomach content, Mugilidae) | Katsuura, Nachi, Wakayama, Japan | [37] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [35] | |||
| Chaetomugilin B (6) | 464 | C24H29ClO7 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [38] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [40] | |||
| C. globosum Z1 | Broussonetia papyrifera (Barks, Moraceae) | Nanjing, Jiangsu, China | [44] | |||
| Chaetomugilin C (7) | 432 | C23H25ClO6 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [38] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [40] | |||
| C. globosum HDN151398 | Sediment sea | South China Sea | [22] | |||
| Chaetomugilin D (8) | 434 | C23H27ClO6 | C. globosum | Adiantum capillus-veneris (Plant, Pteridaceae) | Saint Katherine Protectorate, Sinai, Egypt | [49] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [40] | |||
| C. globosum | Ginkgo bilob (Leaves, Ginkgoaceae) | Linyi, Shandong province, China | [42] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [41] | |||
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [43] | |||
| C. globosum DAOM 240359 | Indoor air samples or building materials | Ontario, Alberta, Saskatchewan, Nova Scotia, Canada | [30] | |||
| C. globosum DAOM 240359 | Indoor air samples or building materials | Ontario, Alberta, Saskatchewan, Nova Scotia, Canada | [50] | |||
| C. globosum | Amaranthus viridis (Leaves, Amaranthaceae) | Central Province of Sri Lanka | [51] | |||
| C. globosum TW1-1 | Armadillidium vulgare (Pill bugs, Armadillidiidae) | Tongji Medical College, Hubei Province, China | [33] | |||
| C. globosum DAOMC 240359 | Damp building materials | Ontario, Alberta, Saskatchewan, Nova Scotia, Canada | [52] | |||
| C. globosum TY1 | Ginkgo biloba (Barks Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| C. globosum CBS148.51 | Cultured | China | [46] | |||
| Chaetomium sp. NA-S01-R1 | Deep-sea | West Pacific Ocean, China | [25] | |||
| C. cochliodes | Indoor buildings | Finland | [53] | |||
| C. globosum | Seawater and marine deposits | Jeju, Korea | [54] | |||
| C. globosum DAOM 240359 | Damp and moldy buildings | Canada | [55] | |||
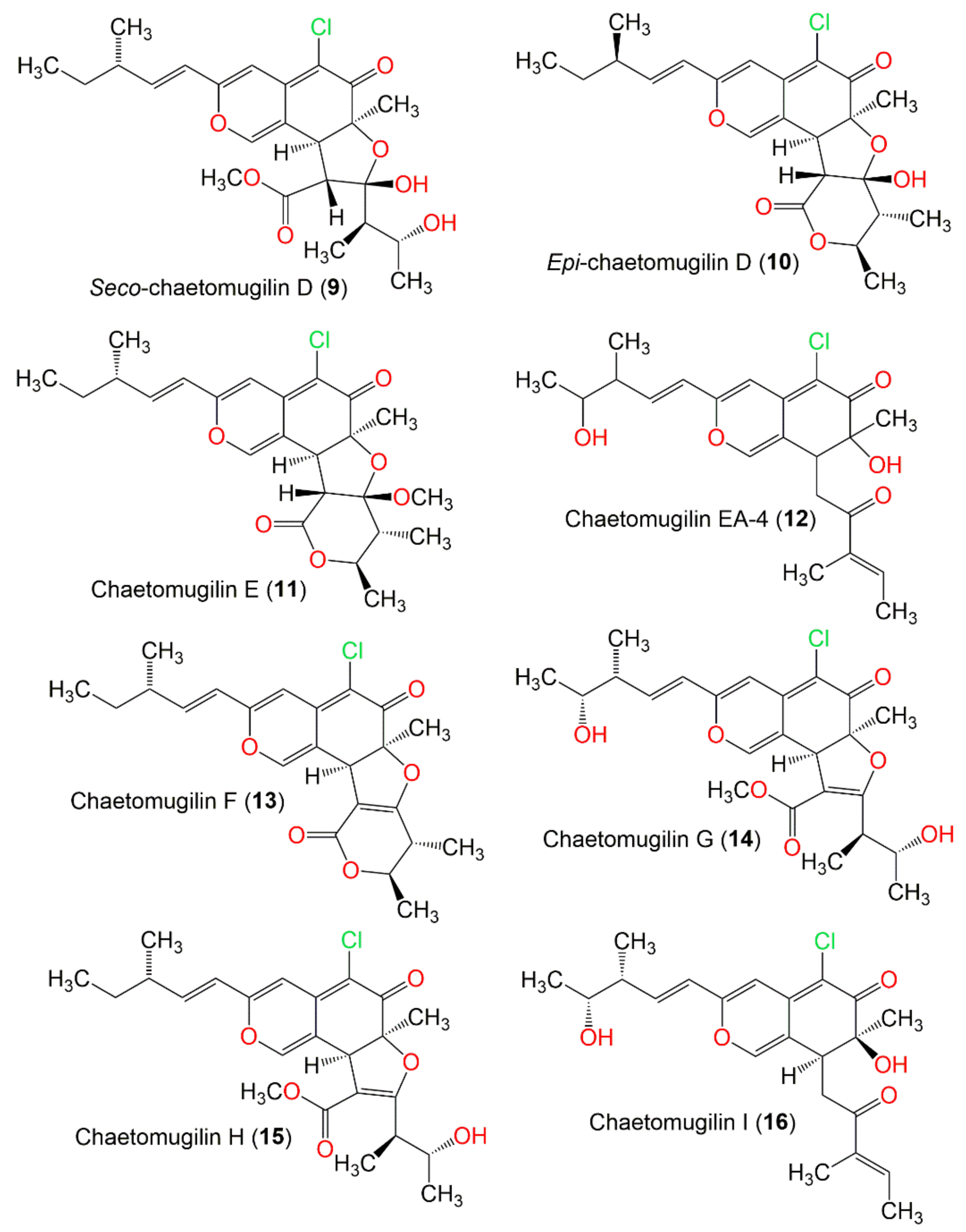
| Seco-chaetomugilin D (9) | 466 | C24H31ClO7 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [48] |
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| C. cupreum | National Fungal Culture Collection | India, Agharkar Research Institute, Pune, India | [29] | |||
| Epi-chaetomugilin D (10) | 434 | C23H27ClO6 | C. globosum | Adiantum capillus-veneris (Plant, Pteridaceae) | Saint Katherine Protectorate, Sinai, Egypt | [49] |
| Chaetomugilin E (11) | 448 | C24H29ClO6 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [40] | |||
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
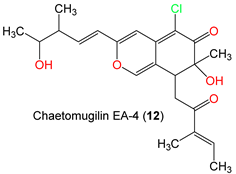
| Chaetomugilin EA-4 (12) | 406 | C22H27ClO5 | C. globosum Kunze ex. 5157 | Soils of wheat field | New Delhi, India | [57] |
| Chaetomugilin F (13) | 416 | C23H25ClO5 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [40] | |||
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| Chaetomugilin G (14) | 464 | C24H29ClO7 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [41] | |||
| Chaetomugilin H (15) | 448 | C24H29ClO6 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [39] |
| C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [41] | |||
| Chaetomugilin I (16) | 406 | C22H27ClO5 | C. globosum | Mugii cephalus (Fish bora stomach content, Mugilidae) | Katsuura, Nachi, Wakayama, Japan | [37] |
| C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] | |||
| C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] | |||
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| C. globosum TY1 | Ginkgo biloba (Barks Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| C. globosum TY-2 | Polygonatum sibiricum, (Roots, Convallariaceae) | Linan, Zhejiang, China | [32] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [47] | |||
| C. globosum Kunze ex. 5157 | Soils of wheat field | New Delhi, India | [57] | |||
| 11-Epi-chaetomugilin I (17) | 406 | C22H27ClO5 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [59] |
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| Chaetomugilin J (18) | 390 | C22H27ClO4 | C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] |
| C. globosum | Adiantum capillus-veneris (Plant, Pteridaceae) | Saint Katherine Protectorate, Sinai, Egypt | [49] | |||
| C. globosum | Mugii cephalus (Fish bora stomach content, Mugilidae) | Katsuura, Nachi, Wakayama, Japan | [37] | |||
| C. globosum | Amaranthus viridis (Leaves, Amaranthaceae) | Central Province of Sri Lanka | [51] | |||
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| C. globosum TY-2 | Polygonatum sibiricum (Root, Convallariaceae) | Linan, Zhejiang, China | [32] | |||
| C. globosum | Polygonatum sibiricum, (Root, Convallariaceae) | China | [34] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [47] | |||
| C. globosum Kunze ex. 5157 | Soils of wheat field | New Delhi, India | [57] | |||
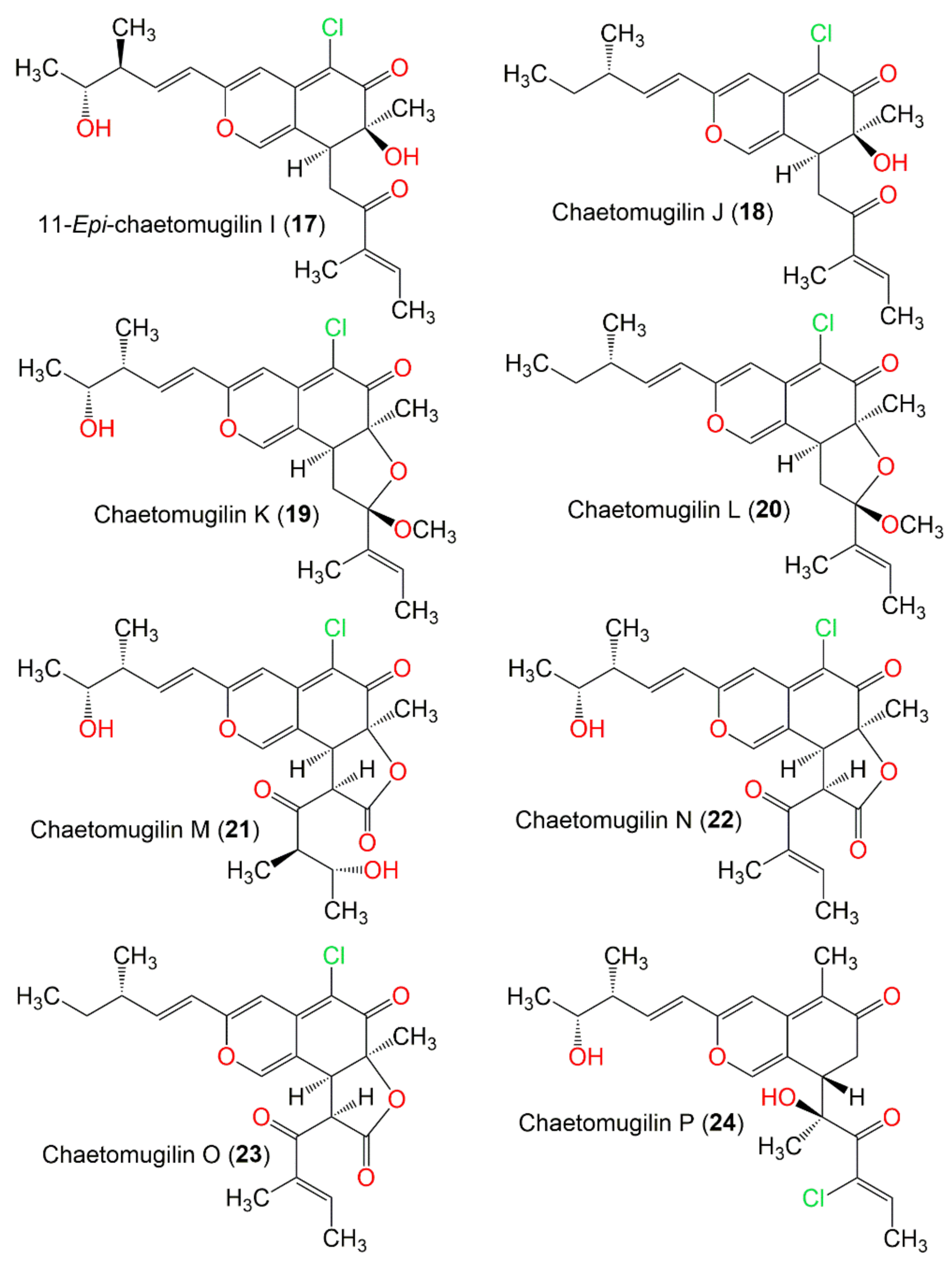
| Chaetomugilin K (19) | 420 | C23H29ClO5 | C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] |
| Chaetomugilin L (20) | 404 | C23H29ClO4 | C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] |
| Chaetomugilin M (21) | 450 | C23H27ClO7 | C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] |
| Chaetomugilin N (22) | 432 | C23H25ClO6 | C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] |
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| Chaetomugilin O (23) | 416 | C23H25ClO5 | C. globosum | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [58] |
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| C. globosum TY-2 | Polygonatum sibiricum, (Root, Convallariaceae) | Linan, Zhejiang, China | [32] | |||
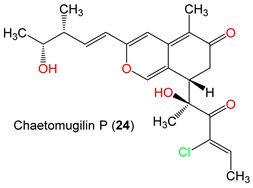
| Chaetomugilin P (24) | 406 | C22H27ClO5 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [59] |
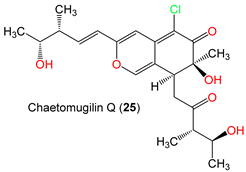
| Chaetomugilin Q (25) | 424 | C22H29ClO6 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [59] |
| C. globosum TW1-1 | Armadillidium vulgare (Pill bugs, Armadillidiidae) | Tongji Medical College, Hubei Province, China | [33] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| C. globosum TY1 | Ginkgo bilob (Leaves, Ginkgoaceae) | Linyi, Shandong province, China | [47] | |||
| C. globosum TY-2 | Polygonatum sibiricum, (Root, Convallariaceae) | Linan, Zhejiang, China | [32] | |||
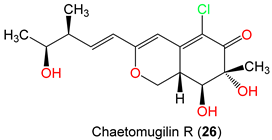
| Chaetomugilin R (26) | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [59] | ||
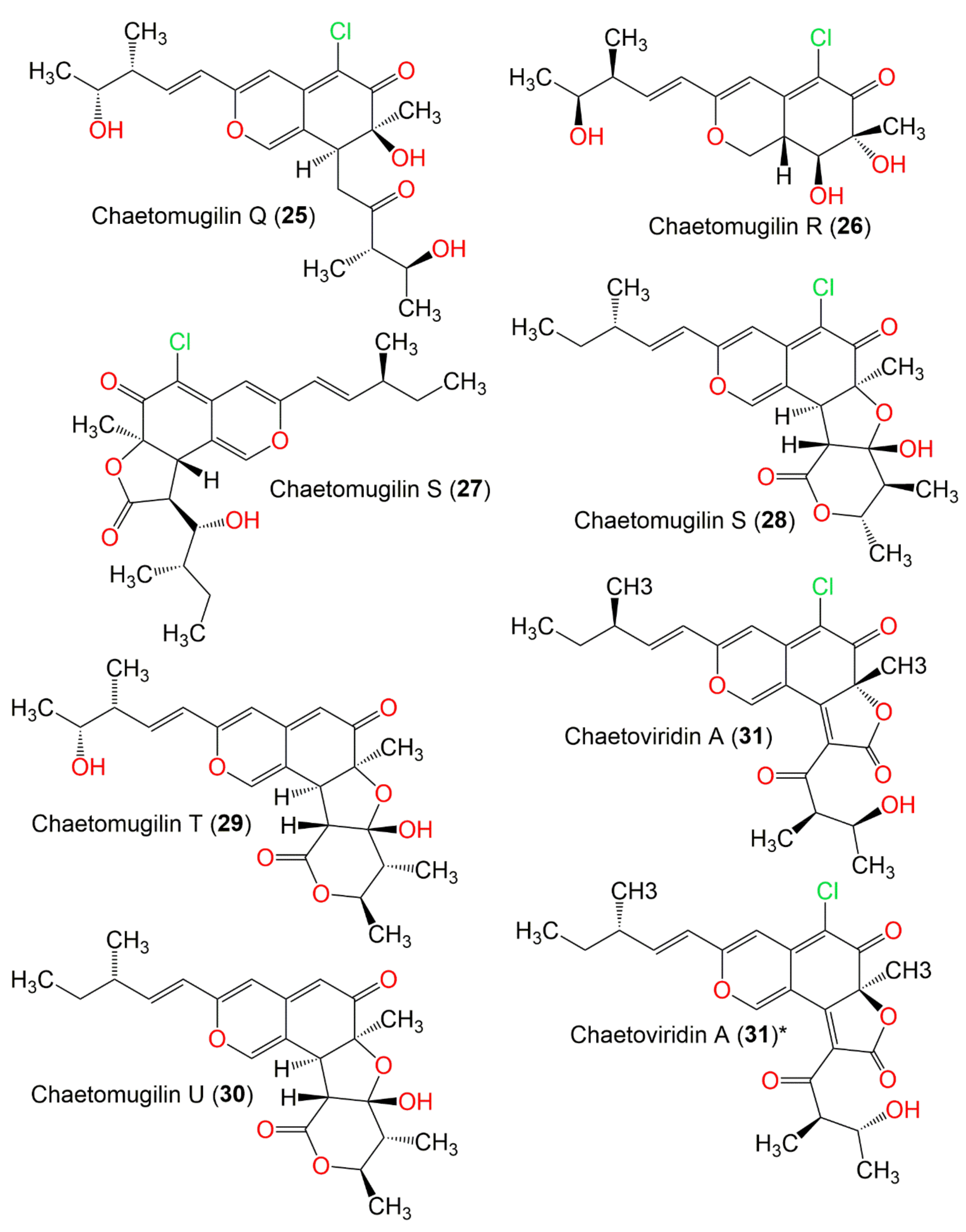
| Chaetomugilin S (27) | 434 | C23H27ClO6 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [43] |
| C. globosum TW1-1 | Armadillidium vulgare (Pillbugs, Armadillidiidae) | Tongji Medical College, Hubei Province, China | [33] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [45] | |||
| Chaetomugilin S (28) | 420 | C23H29ClO5 | C. elatum No. 89-1-3-1 | Ramalina calicaris (Lichen, Ramalinaceae) | Zixishan Mountain, Yunnan, China | [31] |
| Chaetomugilin T (29) | 416 | C23H28O7 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [43] |
| Chaetomugilin U (30) | 406 | C23H28O6 | C. globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [43] |
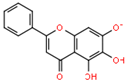
| Chaetoviridin A (31) | 432 | C23H25ClO6 | C. globosum DAOM 240359 | Indoor air samples or building materials | Ontario, Alberta, Saskatchewan, Nova Scotia, Canada | [30,50] |
| Chaetomium sp. NA-S01-R1 | Deep sea | West Pacific Ocean, China | [25] | |||
| C. cochliodes VTh01 C. cochliodes CTh05 | Soil | Ubon Rajathanee province, Bangkok, Thailand Chiangrai province, Bangkok, Thailand | [60] | |||
| C. globosum | Adiantum capillus-veneris (Plant, Pteridaceae) | Saint Katherine Protectorate, Sinai, Egypt | [49] | |||
| C. globosum 5157 | Soils of wheat field | New Delhi, India | [57] | |||
| C. globosum var. flavo-viride | Culture | - | [27] | |||
| C. globosum F0142 | Echinochloa crusgalli (Stems, Poaceae) | Korea | [61] | |||
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| C. siamense | Soil | Bangkok, Thailand | [62] | |||
| C. globosum | Cucumber soil (Rhizosphere) | Egypt | [63] | |||
| C. globosum CIB-160 | - | China | [64] | |||
| C. subafine | Culture | Japan | [65] | |||
| C. globosum DAOM 240359 | Damp and moldy buildings | Canada | [55] | |||
| C. globosum | Sea water and marine deposits | Jeju, Korea | [54] | |||
| C. globosum F211_UMNG | Protium heptaphyllum (Leaves, Burseraceae) | Foothill of the west Colombian Andes mountains, Aguazul, Casanare, Colombia | [66] | |||
| C. globosum 22-10 | Soil | PaLong ZangBu Brook, Tibet, China | [67] | |||
| C. globosum | Artemisia desterorum (Roots, Asteraceae) | Tengger Desert, Ningxia Province, China | [68] | |||
| C. globosum CEF-082 | Gossypium arboreum (Plant, Malvaceae) | China | [69] | |||
| C. globosum E-C-2 | Apostichopus japonicas (Surface muscle, Stichopodidae) | Chengshantou Island, Weihai City, the Yellow Sea, China | [70] | |||
| C. globosum | Indoor buildings | Finland | [53] | |||
| C. cochliodes | indoor buildings | Finland | [53] | |||
| C. globosum MP4-S01-7 | Sea water | West Pacific Ocean, China | [71] | |||
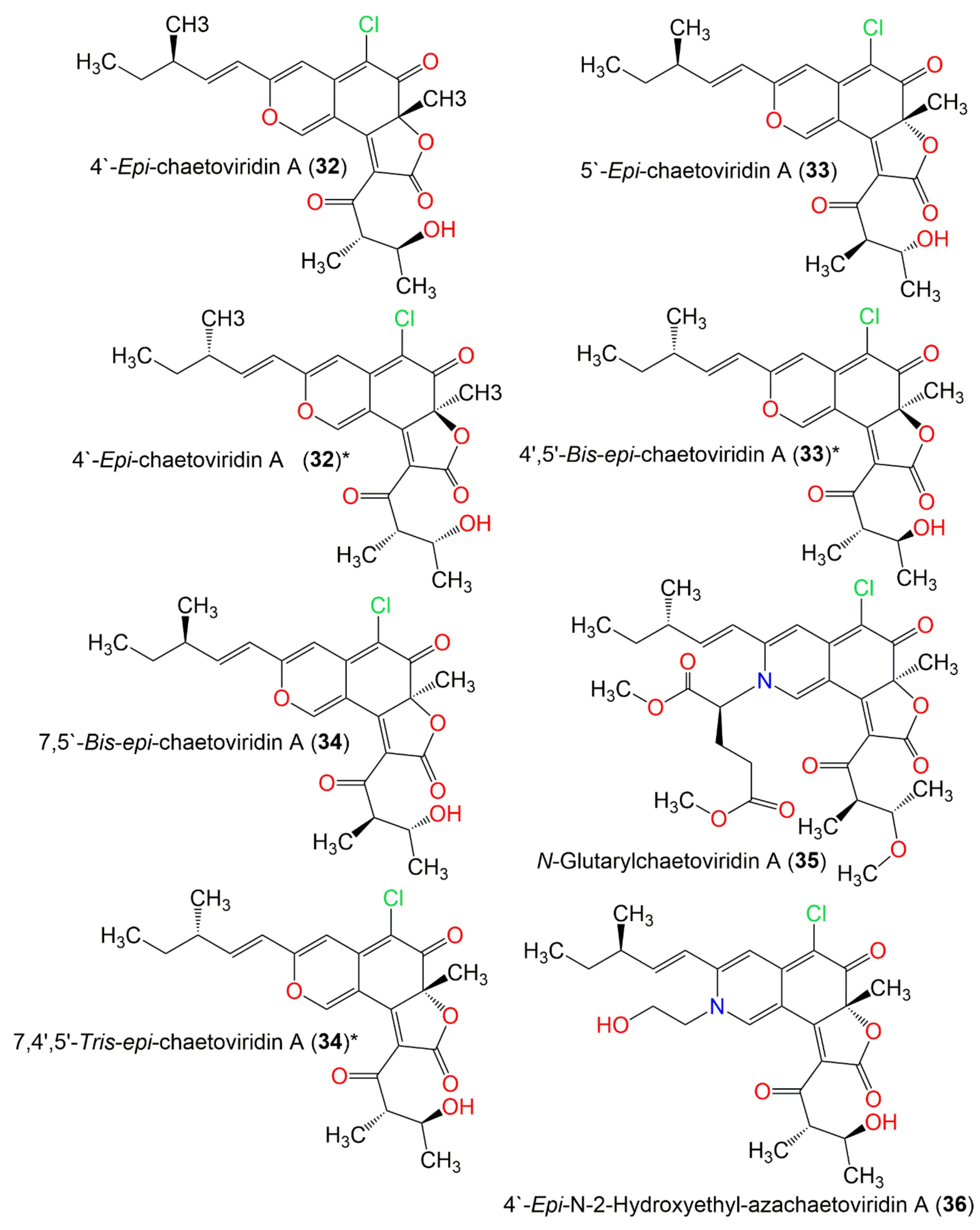
| 4′-Epi-chaetoviridin A (32) | 432 | C23H25ClO6 | C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] |
| C. globosum F211_UMNG | Protium heptaphyllum (Leaves, Burseraceae) | Foothill of the west Colombian Andes mountains, Aguazul, Casanare, Colombia | [66] | |||
| 5′-Epi-chaetoviridin A (33) | 432 | C23H25ClO6 | C. cochliodes VTh01 C. cochliodes CTh05 | Soil | Ubon Rajathanee province, Bangkok, Thailand Chiangrai province, Bangkok, Thailand | [60] |
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong province, China | [72] | |||
| C. globosum CDW7, | Ginkgo biloba (Leaves, Ginkgoaceae) | China | [73] | |||
| C. globosum 22-10 | Soil | Palong Zangbu Brook, Tibet, China | [67] | |||
| C. globosum F211_UMNG | Protium heptaphyllum (Leaves, Burseraceae) | Foothill of the west Colombian Andes mountains, Aguazul, Casanare, Colombia | [66] | |||
| 7,5′-Bis-epi-chaetoviridin A (34) | 432 | C23H25ClO6 | C. elatum No. 89-1-3-1 | Ramalina calicaris (Lichen, Ramalinaceae) | Zixishan Mountain, Yunnan, China | [31] |
| N-Glutarylchaetoviridin A (35) | 603 | C31H38ClNO9 | C. globosum HDN151398 | Sea sediment | South China Sea | [22] |
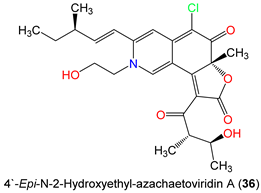
| 4′-Epi-N-2-Hydroxyethyl-azachaetoviridin A (36) | 475 | C25H30ClNO6 | C. globosum DAOM 240359 | Indoor air samples or building materials | Ontario, Alberta, Saskatchewan, Nova Scotia, Canada | [50] |
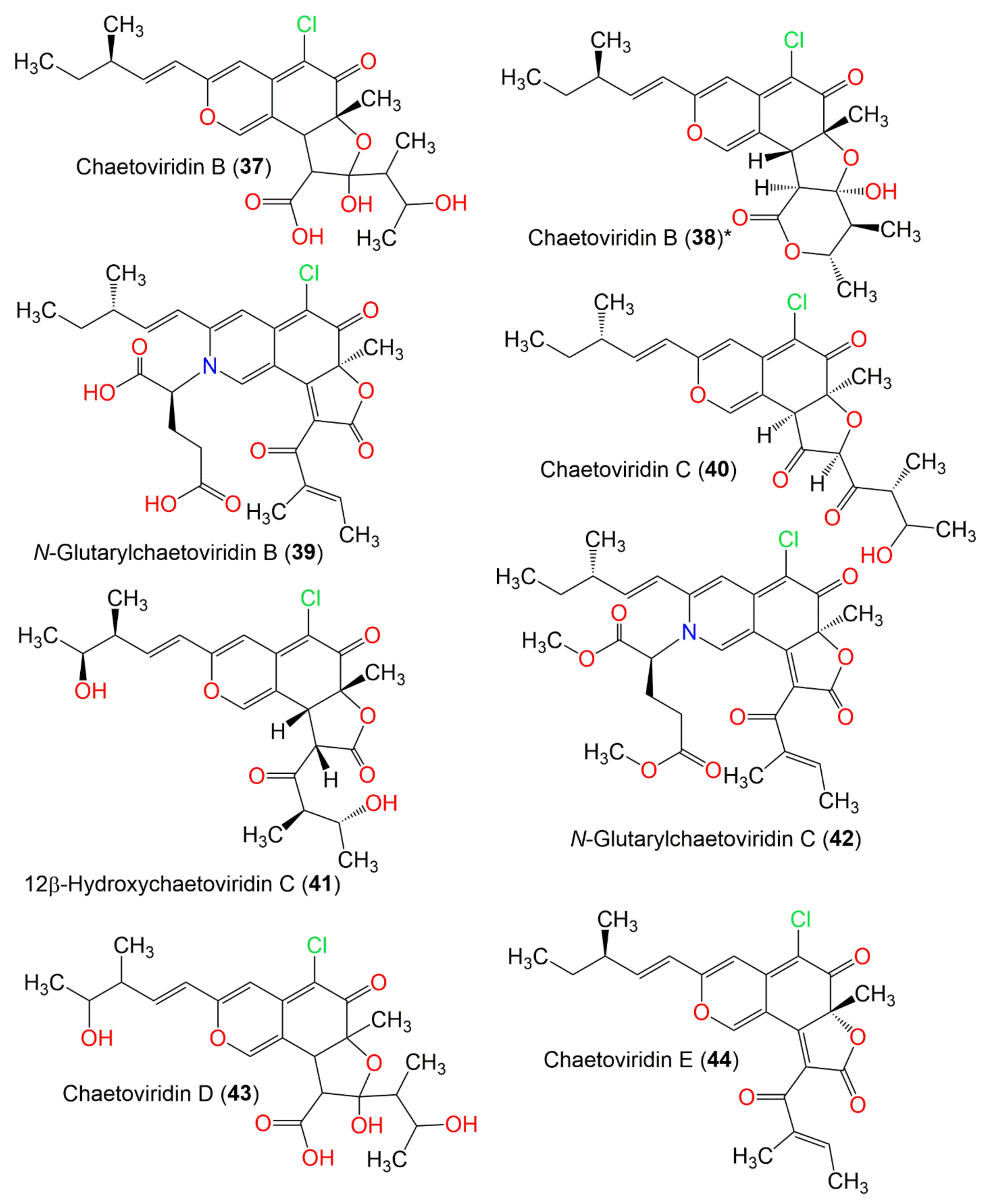
| Chaetoviridin B (37) | 452 | C23H29ClO7 | C. globosum var. flavo-viride | Culture | - | [27] |
| C. globosum F0142 | Echinochloa crusgalli (Stems, Poaceae) | Korea | [61] | |||
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| C. globosum | Cucumber soil (Rhizosphere) | Egypt | [63] | |||
| Chaetoviridin B (38) | C. globosum 5157 | Soils of wheat field | New Delhi, India | [57] | ||
| C. globosum E-C-2 | Apostichopus japonicas (Surface muscle, Stichopodidae) | Chengshantou Island, Weihai City, the Yellow Sea, China | [70] | |||
| C. globosum Z1 | Broussonetia papyrifera (Barks, Moraceae) | Nanjing, Jiangsu, China | [44] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong, China | [47] | |||
| Chaetomium sp. | Dromaius novaehollandiae (Scat, Casuariidae) | Australia | [74] | |||
| C. globosum | Adiantum capillus-veneris (Plant, Pteridaceae) | Saint Katherine Protectorate, Sinai, Egypt | [49] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong province, China | [72] | |||
| N-Glutarylchaetoviridin B (39) | 543 | C28H30ClNO8 | C. globosum HDN151398 | Sea sediment | South China Sea | [22] |
| Chaetoviridin C (40) | 434 | C23H27ClO6 | C. globosum var. flavo-viride | Culture | - | [27] |
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| Chaetomium globosum OUPS-T106B-6 | Mugil cephalus (Marine fish, Mugilidae) | Katsuura Bay, Japan | [41] | |||
| C. globosum | Indoor buildings | Finland | [53] | |||
| 12β-Hydroxychaetoviridin C (41) | 450 | C23H27ClO7 | C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] |
| N-Glutarylchaetoviridin C (42) | 571 | C30H34ClNO8 | C. globosum HDN151398 | Sea sediment | South China Sea | [22] |
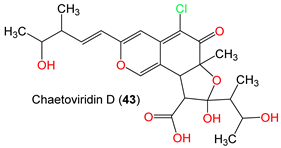
| Chaetoviridin D (43) | 486 | C23H29ClO8 | C. globosum var. flavo-viride | Culture | Spain | [27] |
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| Chaetomium sp. | Dromaius novaehollandiae (Scat, Casuariidae) | Australia | [74] | |||
| Chaetoviridin E (44) | 414 | C23H23ClO5 | C. globosum 5157 | Soils of wheat field | New Delhi, India | [57] |
| C. globosum MP4-S01-7 | Sea water | West Pacific Ocean, China | [71] | |||
| C. globosum | Artemisia desterorum (Roots, Asteraceae) | Tengger Desert in Ningxia, China. | [68] | |||
| C. globosum E-C-2 | Apostichopus japonicas (Surface muscle, Stichopodidae) | Chengshantou Island, Weihai City, the Yellow Sea, China | [70] | |||
| Chaetomium sp. NA-S01-R1 | Deep sea | West Pacific Ocean, China | [25] | |||
| C. globosum 22-10 | Soil | PaLong ZangBu Brook, Tibet, China | [67] | |||
| C. globosum | Sea water and marine deposits | Jeju, Korea | [54] | |||
| C. cochliodes CTh05 | Soil | Ubon Rajathanee province, Bangkok, Thailand | [60] | |||
| C. globosum | Adiantum capillus-veneris (Plant, Pteridaceae) | Saint Katherine Protectorate, Sinai, Egypt | [49] | |||
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| Chaetomium siamense | Soil | Bangkok, Thailand | [62] | |||
| Chaetomium sp. | Dromaius novaehollandiae (Scat, Casuariidae) | Australia | [74] | |||
| C. globosum TY1 | Ginkgo biloba (Barks, Ginkgoaceae) | Linyi, Shandong province, China | [72] | |||
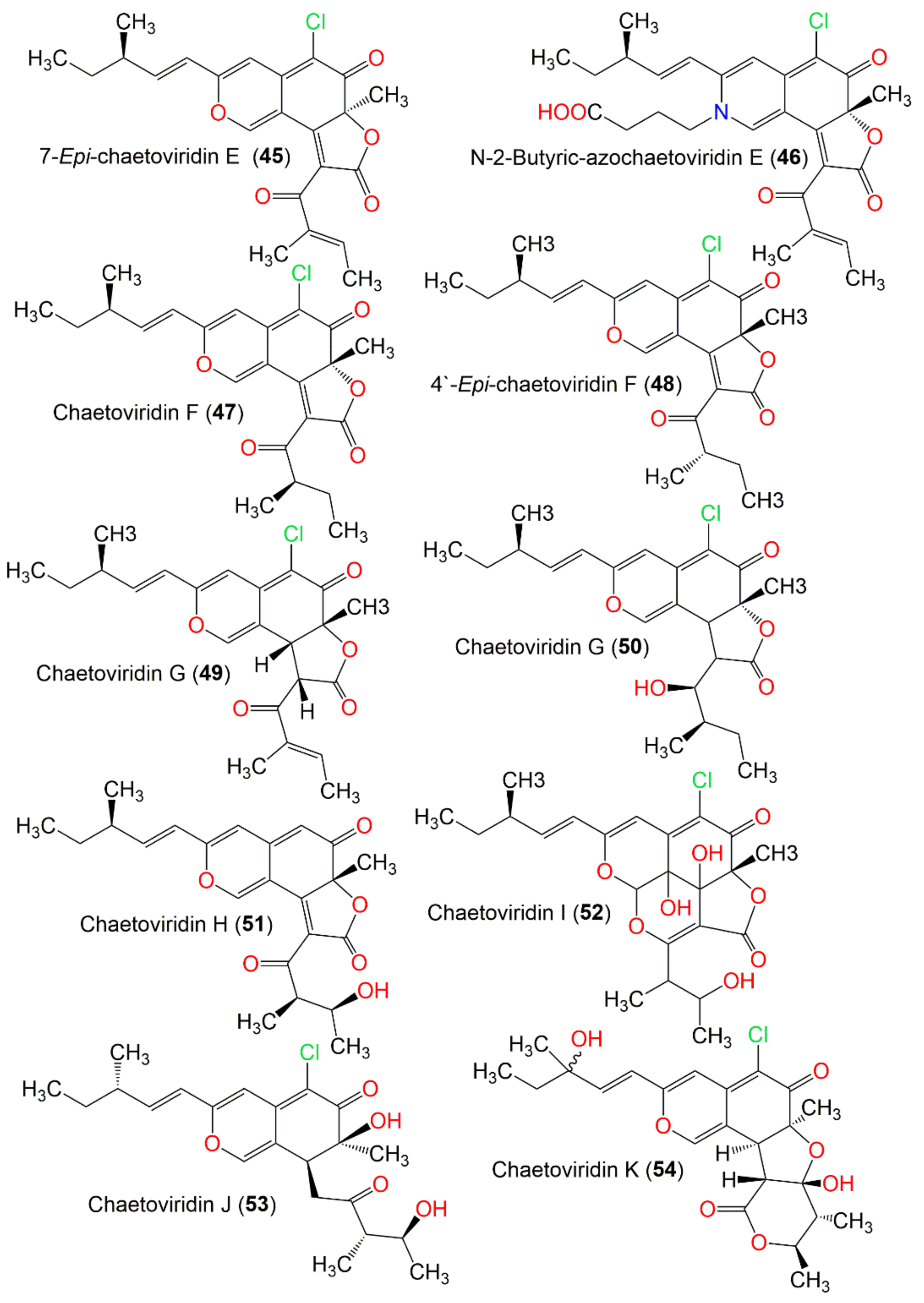
| 7-Epi-chaetoviridin E (45) | 414 | C23H23ClO5 | C. elatum No. 89-1-3-1 | Ramalina calicaris (Lichen, Ramalinaceae) | Zixishan Mountain, Yunnan, China | [31] |
| N-2-Butyric-azochaetoviridin E (46) | 499 | C27H30ClNO6 | C. globosum DAOM 240359 | Indoor air samples or building materials | Ontario, Alberta, Saskatchewan, Nova Scotia, Canada | [50] |
| Chaetoviridin F (47) | 416 | C23H25ClO5 | C. cochliodes VTh01 C. cochliodes CTh05 | Soil | Ubon Rajathanee province, Bangkok, Thailand Chiangrai province, Bangkok, Thailand | [60] |
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| 4′-Epi-chaetoviridin F (48) | 416 | C23H25ClO5 | C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] |
| Chaetoviridin G (49) | 416 | C23H25ClO5 | C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] |
| Chaetoviridin G (50) | 420 | C23H29ClO5 | C. siamense | Soil | Bangkok, Thailand | [62] |
| Chaetoviridin H (51) | 398 | C23H26ClO6 | C. globosum CBS148.51 | Cultured | China | [46] |
| C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] | |||
| Chaetoviridin I (52) | 466 | C23H27ClO8 | C. globosum | Viguiera robusta (Leaves, Asteraceae) | Spain | [36] |
| Chaetoviridin J (53) | 408 | C22H29ClO5 | C. globosum TY1 | Ginkgo biloba (Barks Ginkgoaceae) | Linyi, Shandong, China | [47] |
| C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] | |||
| C. globosum | Seawater and marine deposits | Jeju, Korea | [54] | |||
| Chaetoviridin K (54) | 450 | C23H27ClO7 | C. globosum | Wikstroemia uva-ursi (Leaves, Thymelaeaceae) | Hawaiian Islands, USA | [56] |
| Compound Name | Biological Activity | Assay, Organism, or Cell Line | Biological Results | Positive Control | Refs. |
|---|---|---|---|---|---|
| Chaetomugilin 106B-6 XXVIII (1) | Cytotoxicity | MTT/P388 | 32.0 µM (IC50) | 5-FU 1.7 µM (IC50) | [37,58] |
| MTT/HL-60 | 51.8 µM (IC50) | 5-FU 2.7 µM (IC50) | [37,58] | ||
| MTT/L1210 | 67.1 µM (IC50) | 5-FU 1.1 µM (IC50) | [37,58] | ||
| MTT/KB | 58.8 µM (IC50) | 5-FU 7.7 µM (IC50) | [37,58] | ||
| Chaetomugilin A (2) | Cytotoxicity | MTT/P388 | 8.7 µM (IC50) | 5-FU 1.7 µM (IC50) | [38,40] |
| MTT/HL-60 | 7.3 µM (IC50) | 5-FU 2.7 µM (IC50) | [38,40] | ||
| MTT/HL-60 | 6.4 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] | ||
| MTT/K562 | 11.1 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/BEL-7402 | 17.9 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/HCT-116 | 6.1 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/HeLa | 20.3 µM (IC50) | Adriamycin 0.6 µM (IC50) | [22] | ||
| SRB/L-02 | 15.2 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/MGC-803 | 15.3 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/HO8910 | 12.1 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/SH-SY5Y | 23.4 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/NCI-H1975 | 18.3 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/U87 | 27.1 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] | ||
| SRB/MDA-MB-231 | 22.7 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| 11-Epi-chaetomugilin A (4) | Cytotoxicity | MTT/P388 | 88.9 µM (IC50) | 5-FU 1.7 µM (IC50) | [35] |
| MTT/HL-60 | 66.7 µM (IC50) | 5-FU 2.7 µM (IC50) | [35] | ||
| MTT/P388 | 88.9 µM (IC50) | 5-FU 1.7 µM (IC50) | [37,58] | ||
| MTT/HL-60 | 66.7 µM (IC50) | 5-FU 2.7 µM (IC50) | [37,58] | ||
| MTT/L1210 | 80.2 µM (IC50) | 5-FU 1.1 µM (IC50) | [37,58] | ||
| Chaetomugilin B (6) | Cytotoxicity | MTT/P388 | 18.7 µM (IC50) | 5-FU 1.7 µM (IC50) | [38,40] |
| MTT/HL-60 | 16.5 µM (IC50) | 5-FU 2.7 µM (IC50) | [38,40] | ||
| Chaetomugilin C (7) | Cytotoxicity | MTT/P388 | 3.6 µM (IC50) | 5-FU 1.7 µM (IC50) | [38,40] |
| MTT/HL-60 | 2.7 µM (IC50) | 5-FU 2.7 µM (IC50) | [38,40] | ||
| MTT/HL-60 | 6.6 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] | ||
| MTT/K562 | 12.3 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/BEL-7402 | 16.8 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/HCT-116 | 5.7 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/HeLa | 13.2 µM (IC50) | Adriamycin 0.6 µM (IC50) | [22] | ||
| SRB/L-02 | 9.1 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/MGC-803 | 9.6 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/HO8910 | 8.8 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/SH-SY5Y | 19.4 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/NCI-H1975 | 12.1 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/U87 | 17.6 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] | ||
| SRB/MDA-MB-231 | 26.6 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| Chaetomugilin D (8) | Cytotoxicity | MTT/P388 | 7.5 µM (IC50) | 5-FU 1.7 µM (IC50) | [38,40] |
| MTT/HL-60 | 6.8 µM (IC50) | 5-FU 2.7 µM (IC50) | [38,40] | ||
| Phytotoxic activity | Lettuce seed germination bioassay/Root growth inhibition | 24.2 ppm (IC50) | - | [51] | |
| Lettuce seed germination bioassay/Shoot growth inhibition | 27.8 ppm (IC50) | - | [51] | ||
| Antimicrobial | Microplate assay/Vibrio vulnificus | 32.4 µg/mL (MIC) | Erythromycin 2.0 µg/mL (MIC) | [25] | |
| Microplate assay/Vibrio rotiferianus | 15.3 µg/mL (MIC) | Erythromycin 3.9 µg/mL (MIC) | [25] | ||
| Microplate assay/MRSA 1 | 32.2 µg/mL (MIC) | Chloramphenicol 7.6 µg/mL (MIC) | [25] | ||
| Microplate assay/MRSA 2 | 32.4 µg/mL (MIC) | Chloramphenicol 7.5 µg/mL (MIC) | [25] | ||
| Seco-chaetomugilin D (9) | Cytotoxicity | MTT/P388 | 38.6 µM (IC50) | 5-FU 1.7 µM (IC50) | [48] |
| MTT/HL-60 | 47.2 µM (IC50) | 5-FU 2.7 µM (IC50) | [48] | ||
| MTT/L1210 | 53.6 µM (IC50) | 5-FU 3.0 µM (IC50) | [48] | ||
| MTT/KB | 47.2 µM (IC50) | 5-FU 6.0 µM (IC50) | [48] | ||
| Chaetomugilin E (11) | Cytotoxicity | MTT/P388 | 15.7 µM (IC50) | 5-FU 1.7 µM (IC50) | [38,40] |
| MTT/HL-60 | 13.2 µM (IC50) | 5-FU 2.7 µM (IC50) | [38,40] | ||
| Inhibition TNF-α | TNF-α activated NF-kB assay | 11.6 µM (IC50) | TPCK 3.8 µM (IC50) BAY-11 2.0 µM (IC50) | [56] | |
| Inhibition NO | Nitrite assay | 5.8 µM (IC50) | L-NMMA 25.1 µM (IC50) | [56] | |
| Chaetomugilin F (13) | Cytotoxicity | MTT/P388 | 3.3 µM (IC50) | 5-FU 1.7 µM (IC50) | [38,40] |
| MTT/HL-60 | 1.3 µM (IC50) | 5-FU 2.7 µM (IC50) | [38,40] | ||
| Inhibition TNF-α | TNF-α activated NF-kB assay | 5.1 µM (IC50) | TPCK 3.8 µM (IC50) BAY-11, 2.0 µM (IC50) | [56] | |
| Inhibition NO | Nitrite assay | 1.9 µM (IC50) | L-NMMA 25.1 µM (IC50) | [56] | |
| Chaetomugilin G (14) | Cytotoxicity | MTT/P388 | 24.1 µM (IC50) | 5-FU 1.7 µM (IC50) | [38] |
| MTT/HL-60 | 19.8 µM (IC50) | 5-FU 2.7 µM (IC50) | [38] | ||
| MTT/P388 | 24.1 µM (IC50) | 5-FU 1.7 µM (IC50) | [41] | ||
| MTT/HL-60 | 19.8 µM (IC50) | 5-FU 2.7 µM (IC50) | [41] | ||
| MTT/L1210 | 123.6 µM (IC50) | 5-FU 3.0 µM (IC50) | [41] | ||
| MTT/KB | 137.8 µM (IC50) | 5-FU 6.0 µM (IC50) | [41] | ||
| Chaetomugilin H (15) | Cytotoxicity | MTT/P388 | 12.3 µM (IC50) | 5-FU 1.7 µM (IC50) | [38] |
| MTT/HL-60 | 10.3 µM (IC50) | 5-FU 2.7 µM (IC50) | [38] | ||
| MTT/P388 | 12.3 µM (IC50) | 5-FU 1.7 µM (IC50) | [41] | ||
| MTT/HL-60 | 10.3 µM (IC50) | 5-FU 2.7 µM (IC50) | [41] | ||
| MTT/L1210 | 93.3 µM (IC50) | 5-FU 3.0 µM (IC50) | [41] | ||
| MTT/KB | 18.8 µM (IC50) | 5-FU 6.0 µM (IC50) | [41] | ||
| Chaetomugilin I (16) | Cytotoxicity | MTT/P388 | 1.1 µM (IC50) | 5-FU 1.7 µM (IC50) | [37,58,59] |
| MTT/HL-60 | 1.1 µM (IC50) | 5-FU 2.7 µM (IC50) | [37,58,59] | ||
| MTT/L1210 | 1.9 µM (IC50) | 5-FU 1.1 µM (IC50) | [37,58,59] | ||
| MTT/KB | 2.3 µM (IC50) | 5-FU 7.7 µM (IC50) | [37,58,59] | ||
| Inhibition TNF-α | TNF-α activated NF-kB assay | 0.9 µM (IC50) | TPCK 3.8 µM (IC50) BAY-11 2.0 µM (IC50) | [56] | |
| Inhibition NO | Nitrite assay | 0.3 µM (IC50) | L-NMMA 25.1 µM (IC50) | [56] | |
| 11-Epi-chaetomugilin I (17) | Cytotoxicity | MTT/P388 | 0.7 µM (IC50) | 5-FU 1.7 µM (IC50) | [59] |
| MTT/HL-60 | 1.0 µM (IC50) | 5-FU 2.7 µM (IC50) | [59] | ||
| MTT/L1210 | 1.6 µM (IC50) | 5-FU 1.1 µM (IC50) | [59] | ||
| MTT/KB | 1.2 µM (IC50) | 5-FU 7.7 µM (IC50) | [59] | ||
| Inhibition TNF-α | TNF-α activated NF-kB assay | 0.9 µM (IC50) | TPCK 3.8 µM (IC50) BAY-11 2.0 µM (IC50) | [56] | |
| Inhibition NO | Nitrite assay | 0.8 µM (IC50) | L-NMMA 25.1 µM (IC50) | [56] | |
| Chaetomugilin J (18) | Phytotoxic activity | Lettuce seed germination bioassay/Root growth inhibition | 22.6 ppm (IC50) | - | [51] |
| Lettuce seed germination bioassay/Shoot growth inhibition | 21.9 ppm (IC50) | - | [51] | ||
| Cytotoxicity | MTT/P388 | 12.6 µM (IC50) | 5-FU 1.7 µM (IC50) | [37,58] | |
| MTT/HL-60 | 12.6 µM (IC50) | 5-FU 2.7 µM (IC50) | [37,58] | ||
| MTT/L1210 | 2.8 µM (IC50) | 5-FU 1.1 µM (IC50) | [37,58] | ||
| MTT/KB | 8.5 µM (IC50) | 5-FU 7.7 µM (IC50) | [37,58] | ||
| Inhibition TNF-α | TNF-α activated NF-kB assay | 7.6 µM (IC50) | TPCK 3.8 µM (IC50) BAY-11 2.0 µM (IC50) | [56] | |
| Inhibition NO | Nitrite assay | 4.2 µM (IC50) | L-NMMA 25.1 µM (IC50) | [56] | |
| Chaetomugilin K (19) | Cytotoxicity | MTT/P388 | 8.2 µM (IC50) | 5-FU 1.7 µM (IC50) | [58] |
| MTT/HL-60 | 14.1 µM (IC50) | 5-FU 2.7 µM (IC50) | [58] | ||
| MTT/L1210 | 11.2 µM (IC50) | 5-FU 1.1 µM (IC50) | [58] | ||
| MTT/KB | 18.7 µM (IC50) | 5-FU 7.7 µM (IC50) | [58] | ||
| Chaetomugilin L (20) | Cytotoxicity | MTT/P388 | 10.9 µM (IC50) | 5-FU 1.7 µM (IC50) | [58] |
| MTT/HL-60 | 13.1 µM (IC50) | 5-FU 2.7 µM (IC50) | [58] | ||
| MTT/L1210 | 15.6 µM (IC50) | 5-FU 1.1 µM (IC50) | [58] | ||
| MTT/KB | 20.1 µM (IC50) | 5-FU 7.7 µM (IC50) | [58] | ||
| Chaetomugilin N (22) | Cytotoxicity | MTT/P388 | 2.3 µM (IC50) | 5-FU 1.7 µM (IC50) | [58] |
| MTT/HL-60 | 2.3 µM (IC50) | 5-FU 2.7 µM (IC50) | [58] | ||
| MTT/L1210 | 10.6 µM (IC50) | 5-FU 1.1 µM (IC50) | [58] | ||
| MTT/KB | 10.6 µM (IC50) | 5-FU 7.7 µM (IC50) | [58] | ||
| Chaetomugilin O (23) | Cytotoxicity | MTT/P388 | 11.1 µM (IC50) | 5-FU 1.7 µM (IC50) | [58] |
| MTT/HL-60 | 11.1 µM (IC50) | 5-FU 2.7 µM (IC50) | [58] | ||
| MTT/L1210 | 10.1 µM (IC50) | 5-FU 1.1 µM (IC50) | [58] | ||
| MTT/KB | 7.2 µM (IC50) | 5-FU 7.7 µM (IC50) | [58] | ||
| Chaetomugilin P (24) | Cytotoxicity | MTT/P388 | 0.7 µM (IC50) | 5-FU 1.7 µM (IC50) | [59] |
| MTT/HL-60 | 1.2 µM (IC50) | 5-FU 2.7 µM (IC50) | [59] | ||
| MTT/L1210 | 1.5 µM (IC50) | 5-FU 1.1 µM (IC50) | [59] | ||
| MTT/KB | 1.8 µM (IC50) | 5-FU 7.7 µM (IC50) | [59] | ||
| Chaetomugilin Q (25) | Cytotoxicity | MTT/P388 | 49.5 µM (IC50) | 5-FU 1.7 µM (IC50) | [59] |
| MTT/HL-60 | 47.2 µM (IC50) | 5-FU 2.7 µM (IC50) | [59] | ||
| MTT/L1210 | 80.2 µM (IC50) | 5-FU 1.1 µM (IC50) | [59] | ||
| Chaetomugilin R (26) | Cytotoxicity | MTT/P388 | 32.0 µM (IC50) | 5-FU 1.7 µM (IC50) | [59] |
| MTT/HL-60 | 51.8 µM (IC50) | 5-FU 2.7 µM (IC50) | [59] | ||
| MTT/L1210 | 67.1 µM (IC50) | 5-FU 1.1 µM (IC50) | [59] | ||
| MTT/KB | 67.1 µM (IC50) | 5-FU 7.7 µM (IC50) | [59] | ||
| Chaetomugilin S (27) | Caspase-3 inhibitory | Caspase-3 enzymatic assay | 20.6 µM (IC50) | Ac-DEVD-CHO 13.7 nM (IC50) | [31] |
| Chaetomugilin T (29) | Cytotoxicity | MTT/P388 | 62.4 µM (IC50) | 5-FU 1.9 µM (IC50) | [43] |
| MTT/HL-60 | 67.2 µM (IC50) | 5-FU 2.3 µM (IC50) | [43] | ||
| Chaetomugilin U (30) | Cytotoxicity | MTT/P388 | 57.4 µM (IC50) | 5-FU 1.9 µM (IC50) | [43] |
| MTT/HL-60 | 57.4 µM (IC50) | 5-FU 2.3 µM (IC50) | [43] | ||
| MTT/L1210 | 94.8 µM (IC50) | 5-FU 2.2 µM (IC50) | [43] | ||
| Chaetoviridin A (31) | Antifungal | Alternaria mali | 33.3 µg/mL (MIC) | - | [61] |
| Botrytis cinerea | 33.3 µg/mL (MIC) | - | [61] | ||
| Colletotrichum gloeosporioides | 33.3 µg/mL (MIC) | - | [61] | ||
| Fusarium oxysporum | 33.3 µg/mL (MIC) | - | [61] | ||
| Phytophthora capsici | 33.3 µg/mL (MIC) | - | [61] | ||
| Phytophthora infestans | 33.3 µg/mL (MIC) | - | [61] | ||
| Pythium ultimum | 1.23 µg/mL (MIC) | - | [61] | ||
| Magnaporthe grisea | 1.23 µg/mL (MIC) | - | [61] | ||
| Sclerotinia sclerotiorum | 97.5 (% inhibition) | - | [73] | ||
| Botrytis cinerea | 69.1 (% inhibition) | - | [73] | ||
| Fusarium graminearum | 77.0 (% inhibition) | - | [73] | ||
| Phytophthora capsici | 60.7 (% inhibition) | - | [73] | ||
| Fusarium moniliforme | 59.2 (% inhibition) | - | [73] | ||
| 5′-Epi-chaetoviridin A (33) | Cytotoxicity | SRB assay/HepG-2 | 35.3 µM (IC50) | Camptothecin 32.3 µM (IC50) | [72] |
| 7,5′- Bis-epi-chaetoviridin A (34) | Caspase-3 inhibitory | Caspase-3 enzymatic assay | 10.9 µM (IC50) | Ac-DEVD-CHO 13.7 nM (IC50) | [31] |
| N-glutarylchaetoviridin A (35) | Cytotoxicity | MTT/HL-60 | 10.3 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] |
| MTT/K562 | 20.3 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/BEL-7402 | 23.9 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| Chaetoviridin B (37) | Antifungal | Pythium ultimum | 33.3 µg/mL (MIC) | - | [61] |
| Magnaporthe grisea | 33.3 µg/mL (MIC) | - | [61] | ||
| Chaetoviridin B (38) | α-Glucosidase inhibiory | Spectrophotometric assay | 6.328 µM (IC50) | Acarbose 54.74 µM (IC50) | [47] |
| N-Glutarylchaetoviridin C (42) | Cytotoxicity | MTT/HL-60 | 11.1 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] |
| MTT/K562 | 11.7 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/BEL-7402 | 10.9 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/HCT-116 | 11.3 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/HeLa | 22.1 µM (IC50) | Adriamycin 0.6 µM (IC50) | [22] | ||
| SRB/L-02 | 18.2 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/MGC-803 | 6.6 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| SRB/HO8910 | 9.7 µM (IC50) | Adriamycin 0.4 µM (IC50) | [22] | ||
| SRB/NCI-H1975 | 11.2 µM (IC50) | Adriamycin 0.3 µM (IC50) | [22] | ||
| SRB/U87 | 18.3 µM (IC50) | Adriamycin 0.1 µM (IC50) | [22] | ||
| SRB/MDA-MB-231 | 13.2 µM (IC50) | Adriamycin 0.2 µM (IC50) | [22] | ||
| Chaetoviridin E (44) | Cytotoxicity | SRB assay/BC1 | 5.6 µg/mL (IC50) | Ellipticine 0.26 µg/mL (IC50) | [60] |
| Cytotoxicity | SRB assay/NCl-H187 | 3.5 µg/mL (IC50) | Ellipticine 0.32 µg/mL (IC50) | [60] | |
| Cytotoxicity | SRB assay/HepG-2 | 40.6 µM (IC50) | Camptothecin 32.3 µM (IC50) | [72] | |
| Antimalarial | Microculture radioisotope assay/P. falciparum (K1, MDR) | 2.9 µg/mL (IC50) | - | [60] | |
| 7-Epi-chaetoviridin E (45) | Caspase-3 inhibitory | Caspase-3 enzymatic assay | 7.9 µM (IC50) | Ac-DEVD-CHO 13.7 nM (IC50) | [31] |
| Chaetoviridin F (47) | Cytotoxicity | SRB assay/NCl-H187 | 4.5 µg/mL (IC50) | Ellipticine 0.32 µg/mL (IC50) | [60] |
| Cytotoxicity | MTT/P388 | 46.0 µM (IC50) | 5-FU 1.9 µM (IC50) | [43] | |
| MTT/HL-60 | 39.1 µM (IC50) | 5-FU 2.3 µM (IC50) | [43] | ||
| MTT/L1210 | 43.7 µM (IC50) | 5-FU 2.2 µM (IC50) | [43] | ||
| MTT/KB | 34.5 µM (IC50) | 5-FU 20.6 µM (IC50) | [43] |
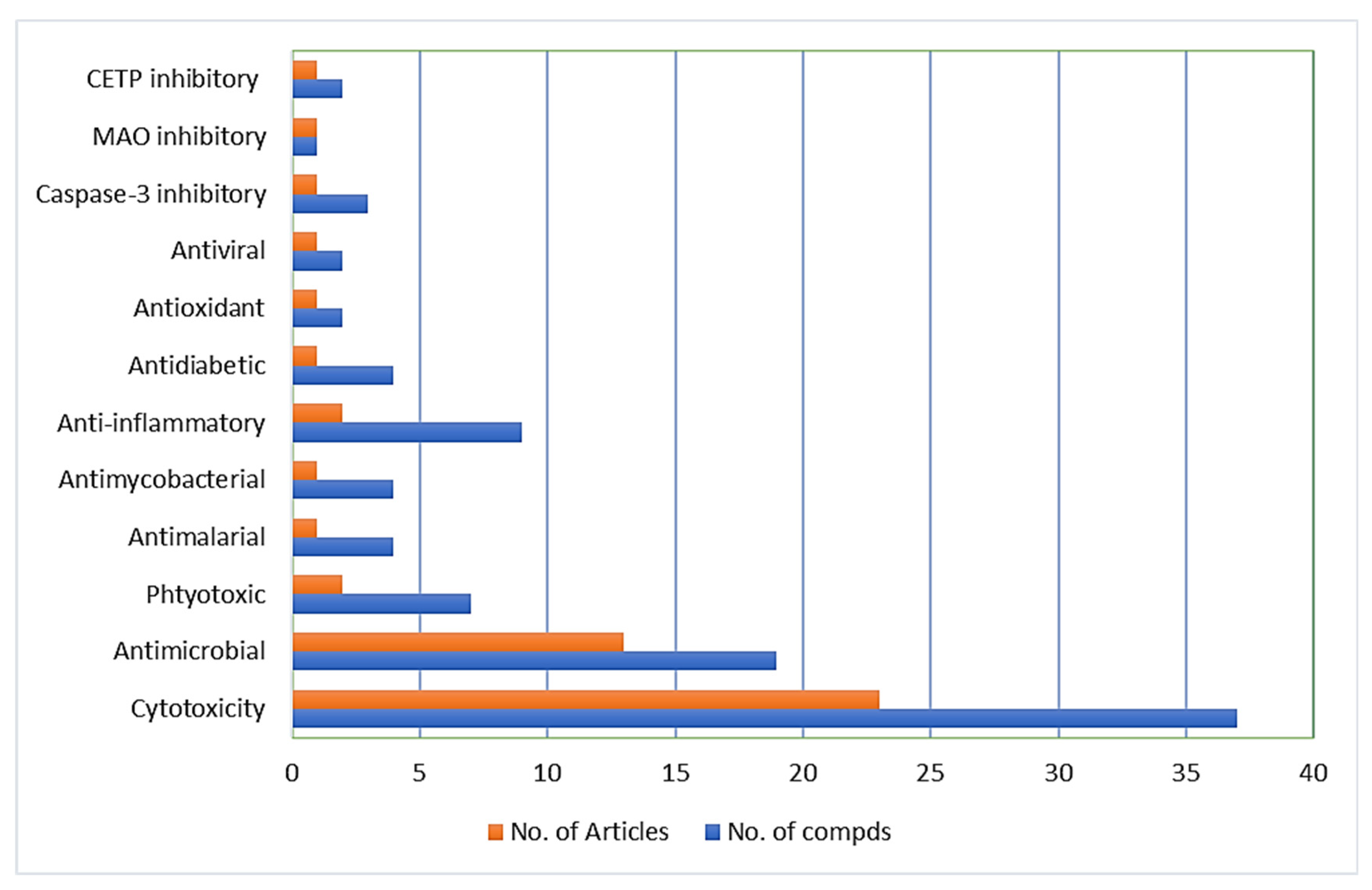
3. Biological Activities
3.1. Cytotoxic Activity
3.2. Antimicrobial Activity
3.3. Phytotoxic Activity
3.4. Antimalarial and Antimycobacterial Activities
3.5. Anti-Inflammatory Activity
3.6. Antidiabetic, Antioxidant, and Antiviral Activities
3.7. Caspase-3 and Monoamine Oxidase (MAO) Inhibitory Activities
3.8. Cholesteryl Ester Transfer Protein Inhibitory Activity
3.9. Anti-SARS-CoV-2 Activity
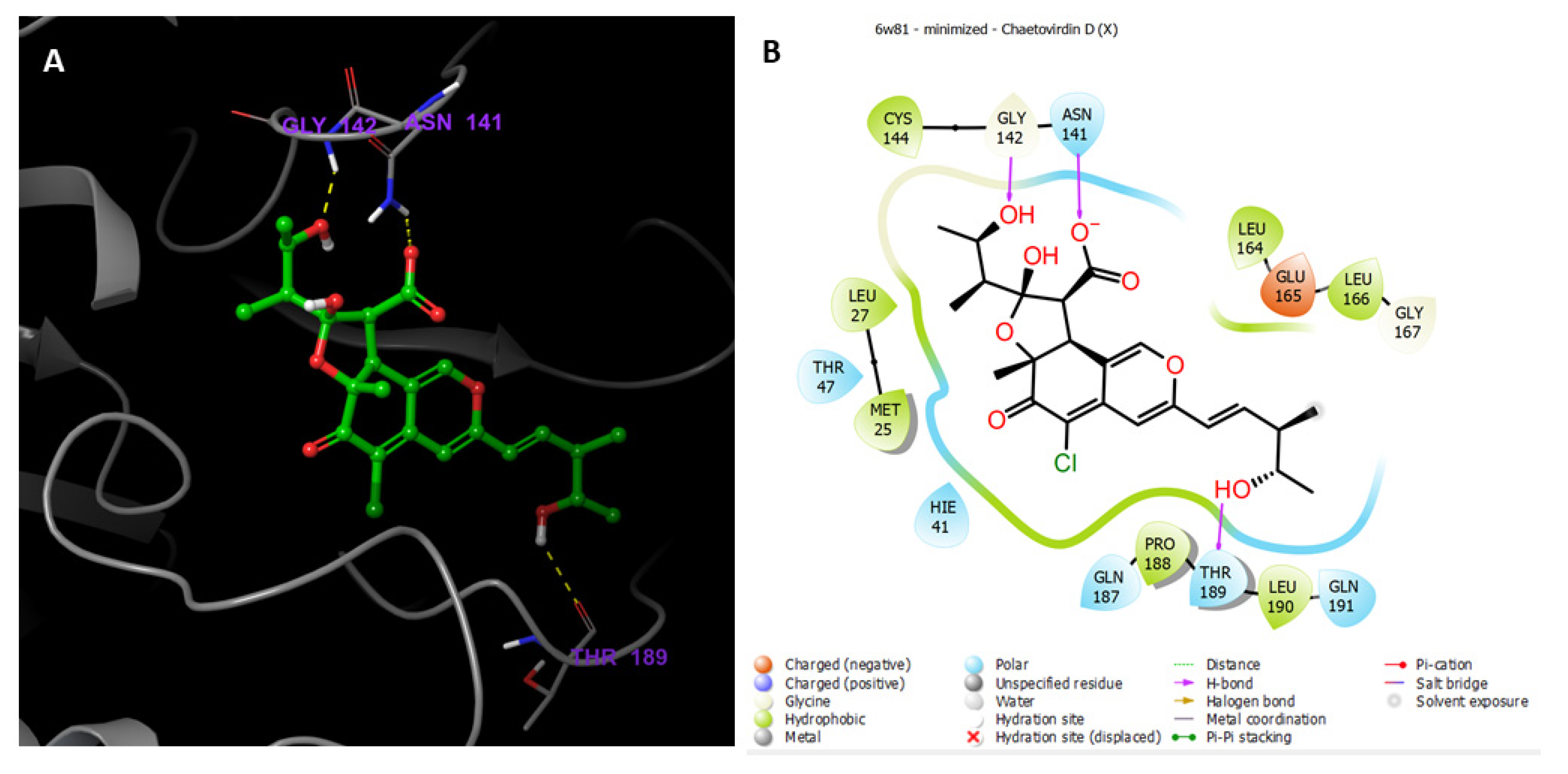
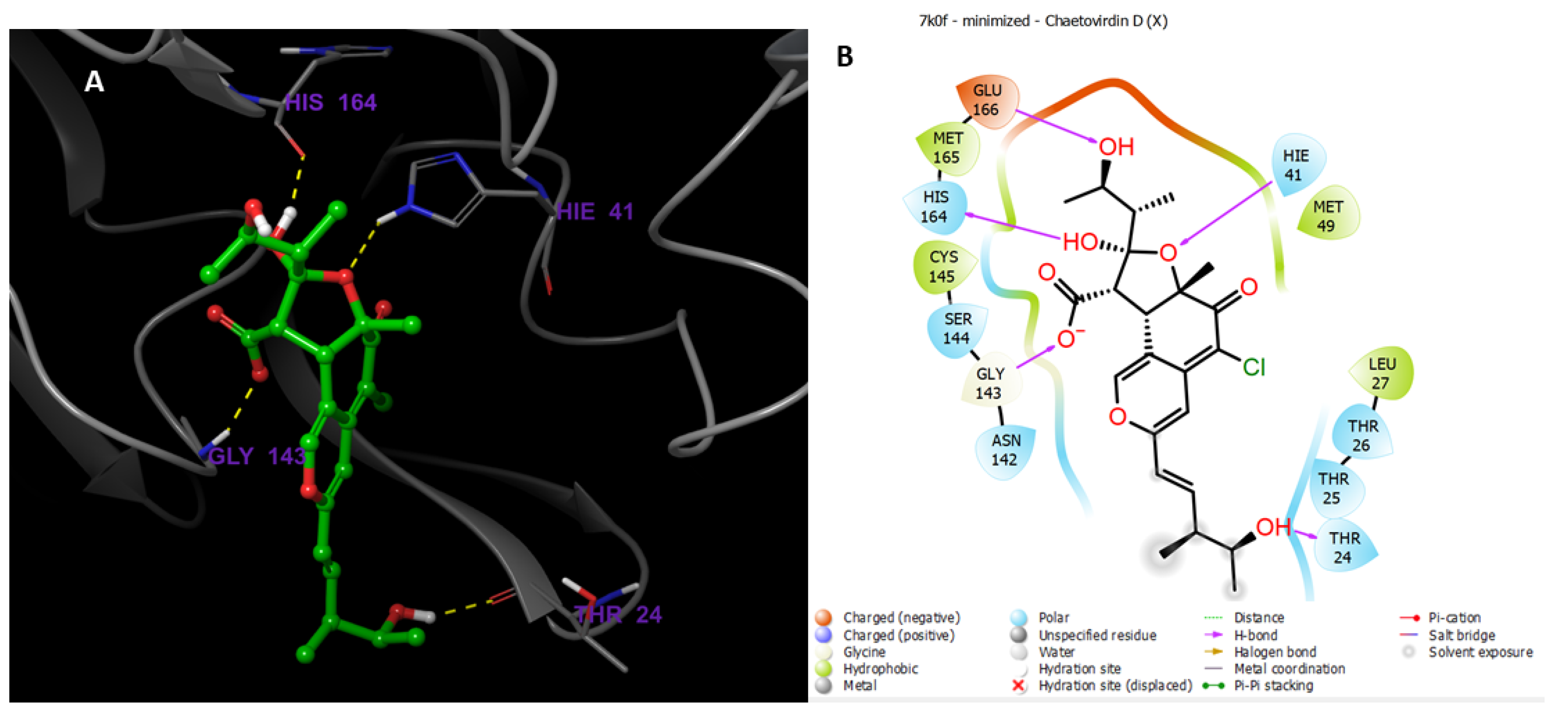
3.9.1. Preparations of Ligands and Proteins
3.9.2. Molecular Docking Studies
3.9.3. In Silico ADMET Properties of Selected Ligands
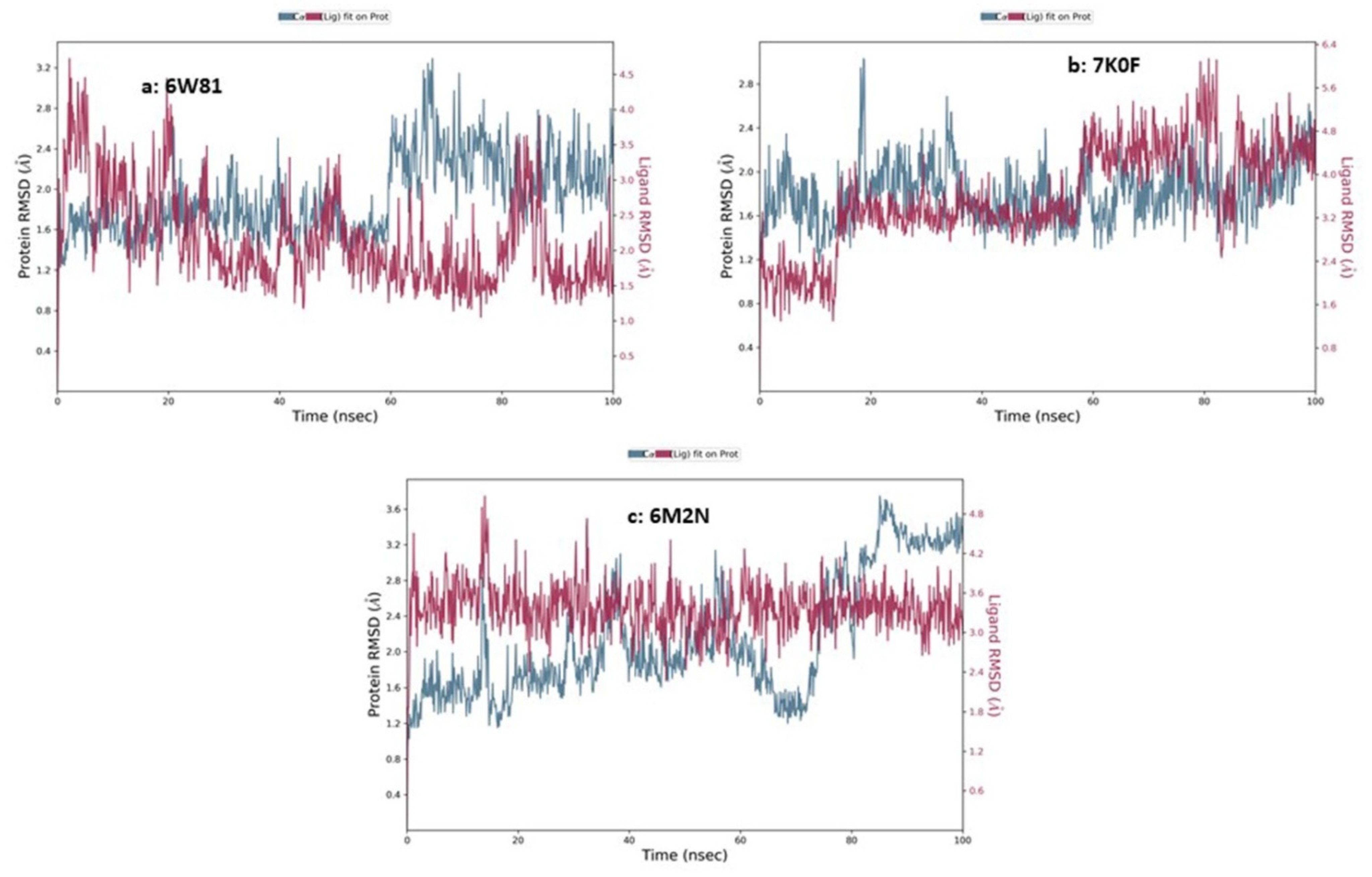
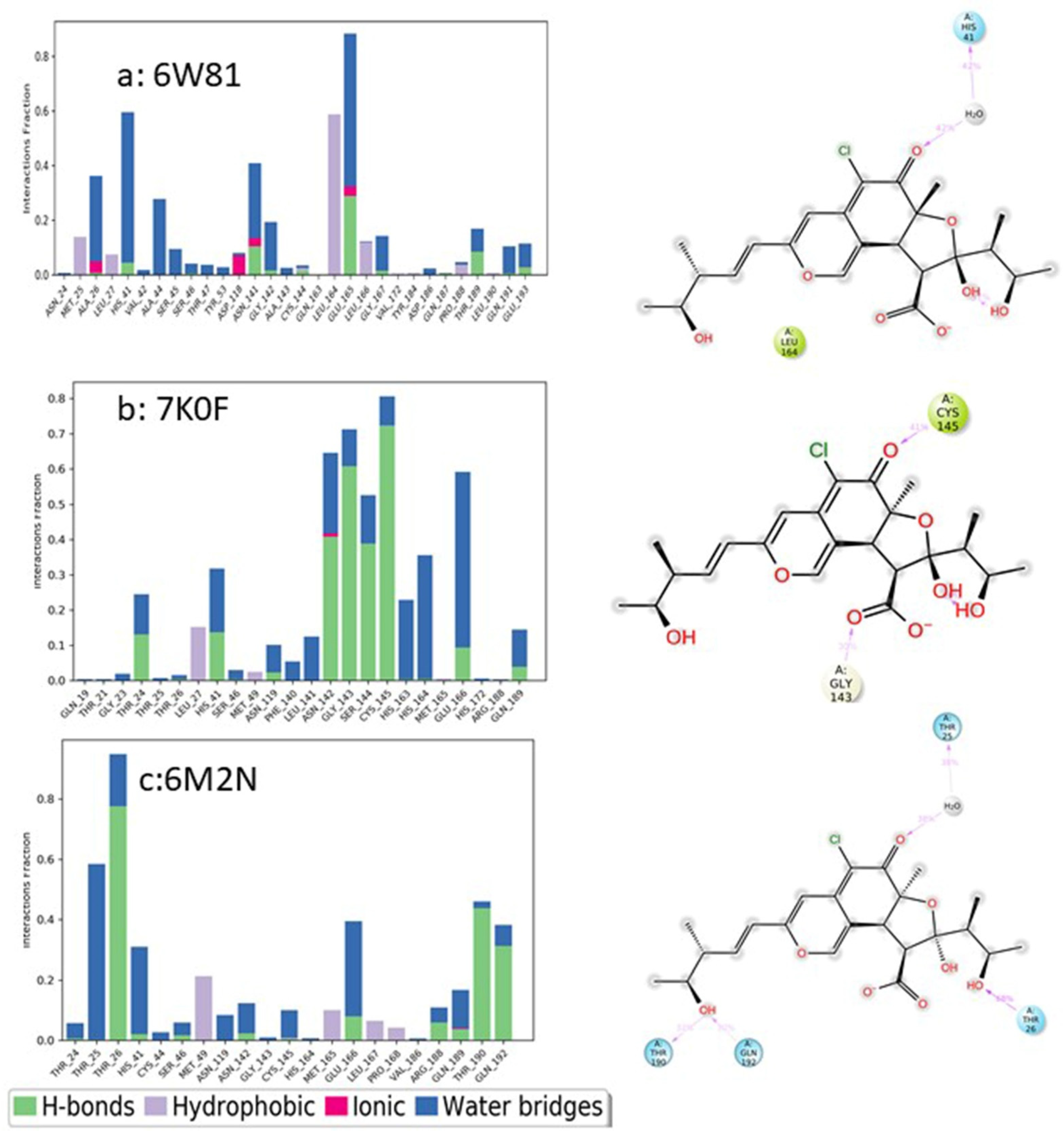
3.9.4. Molecular Dynamics Simulation
3.9.5. Materials and Methods
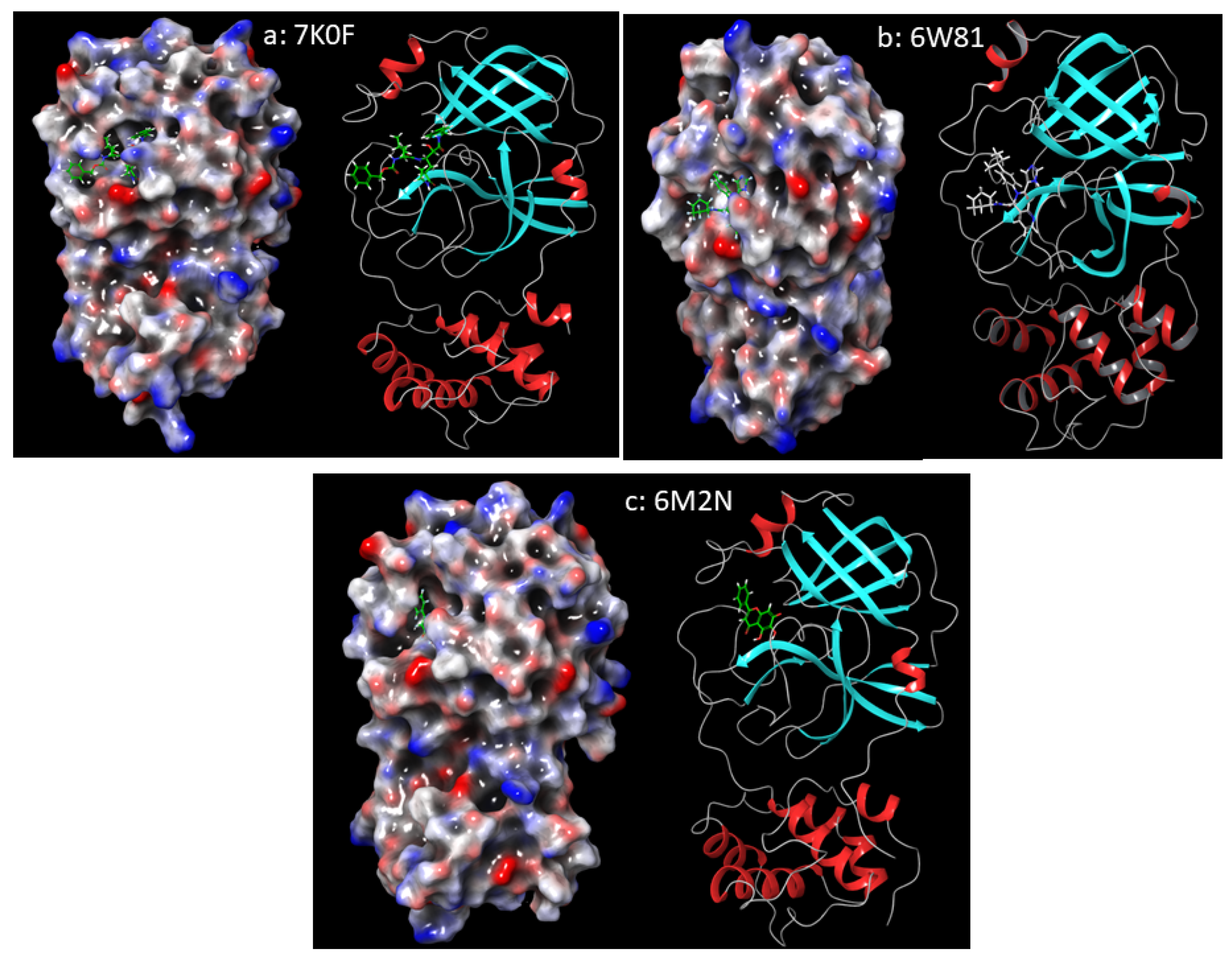
Preparation of PDB Structures
Predictions of ADME Properties
Receptor Grids Generation and Docking
MD Simulations of Chaetovirdin D in Complexes with 6M2N, 6W81, and 7K0f
4. Biosynthesis of Chaetomugilins and Chaetoviridins
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohamed, G.A.; Ibrahim, S.; El-Agamy, D.S.; Elsaed, W.M.; Sirwi, A.; Asfour, H.Z.; Koshak, A.E.; Elhady, S.S. Terretonin as a new protective agent against sepsis-induced acute lung injury: Impact on SIRT1/Nrf2/NF-κBp65/NLRP3 signaling. Biology 2021, 10, 1219. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.A.; Ibrahim, S.R.M. Untapped potential of marine associated Cladosporium species: An overview on secondary metabolites, biotechnological relevance, and biological activities. Mar. Drugs 2021, 19, 645. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Altyar, A.E.; Mohamed, S.G.A.; Mohamed, G.A. Genus Thielavia: Phytochemicals, industrial importance, and biological relevance. Nat. Prod. Res. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Sindi, I.A.; Mohamed, G.A. Biologically active secondary metabolites and biotechnological applications of species of the family Chaetomiaceae (Sordariales): An updated review from 2016 to 2021. Mycol. Prog. 2021, 20, 595–639. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Mohamed, G.A. Humicola genus: Chemical constituents, industrial importance, and biological activities. Curr. Microbiol. 2021. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Bright Side of Fusarium oxysporum: Secondary Metabolites Bioactivities and Industrial Relevance in Biotechnology and Nanotechnology. J. Fungi 2021, 7, 943. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.; Fat’hi, S.M.; Ross, S.A. Fusarithioamide A, a new antimicrobial and cytotoxic benzamide derivative from the endophytic fungus Fusarium chlamydosporium. Biochem. Biophys. Res. Commun. 2016, 479, 211–216. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Abdallah, H.M.; Mohamed, G.A.; Ross, S.A. Integracides H.-New tetracyclic triterpenoids from the endophytic fungus Fusarium sp. Fitoterapia 2016, 112, 161–167. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Ross, S.A. Integracides F and G: New tetracyclic triterpenoids from the endophytic fungus Fusarium sp. Phytochem. Lett. 2016, 15, 125–130. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F.; Khayat, M.T. Biologically active fungal depsidones: Chemistry, biosynthesis, structural characterization, and bioactivities. Fitoterapia 2018, 129, 317–365. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; Zayed, M.F.; El-Kholy, A.A.; Elkhayat, E.S.; Ross, S.A. Fusarithioamide B, a new benzamide derivative from the endophytic fungus Fusarium chlamydosporium with potent cytotoxic and antimicrobial activities. Bioorg. Med. Chem. 2018, 26, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Kamal, H.M.K.; Mohamed, S.G.A.; Khedr, A.I.M. Terretonins from Aspergillus Genus: Structures, biosynthesis, bioactivities, and structural elucidation. Mini Rev. Org. Chem. 2021. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Fungal depsides naturally inspiring molecules: Biosynthesis, structural characterization, and biological activities. Metabolites 2021, 11, 683. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khedr, A.M.I. γ-Butyrolactones from Aspergillus species: Structures, biosynthesis, and biological activities. Nat. Prod. Commun. 2017, 12, 791–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bills, G.F.; Gloer, J.B. Biologically active secondary metabolites from the fungi. Microbiol. Spectr. 2016, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Kornienko, A.; Cimmino, A.; Andolfi, A.; Lefranc, F.; Mathieu, V.; Kiss, R. Fungal metabolites with anticancer activity. Nat. Prod. Rep. 2014, 31, 617–627. [Google Scholar] [CrossRef]
- Cox, R.J.; Simpson, T.J. Fungal type I polyketide synthases. Methods Enzymol. 2009, 459, 49–78. [Google Scholar]
- Simpson, T.J. Fungal polyketide biosynthesis—A personal perspective. Nat. Prod. Rep. 2014, 31, 1247–1252. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef]
- Schümann, J.; Hertweck, C. Advances in cloning, functional analysis and heterologous expression of fungal polyketide synthase genes. J. Biotechnol. 2006, 124, 690–703. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J. Fungal Pigments and Their Roles Associated with Human Health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Ge, X.; Mudassir, S.; Zhou, L.; Yu, G.; Che, Q.; Zhang, G.; Peng, J.; Gu, Q.; Zhu, T.; et al. New glutamine-containing azaphilone alkaloids from deep-sea-derived fungus Chaetomium globosum HDN151398. Mar. Drugs 2019, 17, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapari, S.A.; Meyer, A.S.; Thrane, U. Colorimetric characterization for comparative analysis of fungal pigments and natural food colorants. J. Agric. Food Chem. 2006, 54, 7027–7035. [Google Scholar] [CrossRef] [PubMed]
- Zu, W.Y.; Tang, J.W.; Hu, K.; Zhou, Y.F.; Gou, L.L.; Su, X.Z.; Lei, X.; Sun, H.D.; Puno, P.T. Chaetolactam A, an azaphilone derivative from the endophytic fungus Chaetomium sp. g1. J. Org. Chem. 2021, 86, 475–483. [Google Scholar] [CrossRef]
- Wang, W.; Liao, Y.; Chen, R.; Hou, Y.; Ke, W.; Zhang, B.; Gao, M.; Shao, Z.; Chen, J.; Li, F. Chlorinated azaphilone pigments with antimicrobial and cytotoxic activities isolated from the deep sea derived fungus Chaetomium sp. NA-S01-R1. Mar. Drugs 2018, 16, 61. [Google Scholar] [CrossRef] [Green Version]
- Makrerougras, M.; Coffinier, R.; Oger, S.; Chevalier, A.; Sabot, C.; Franck, X. Total Synthesis and Structural Revision of Chaetoviridins, A. Org. Lett. 2017, 19, 4146–4149. [Google Scholar] [CrossRef]
- Takahashi, M.; Koyama, K.; Natori, S. Four New Azaphilones from Chaetomium globosum var. flavo-viridae. Chem. Pharm. Bull. 1990, 38, 625. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.M.; Yang, S.X.; Qin, J.C. Azaphilones: Chemistry and biology. Chem. Rev. 2013, 113, 4755–4811. [Google Scholar] [CrossRef]
- Wani, N.A.; Khanday, W.I.; Tirumale, S. Evaluation of anticancer activity of Chaetomium cupreum extracts against human breast adenocarcinoma cell lines. Matrix Sci. Pharma. 2020, 4, 31–40. [Google Scholar]
- McMullin, D.R.; Sumarah, M.W.; Miller, J.D. Chaetoglobosins and azaphilones produced by Canadian strains of Chaetomium globosum isolated from the indoor environment. Mycotoxin Res. 2013, 29, 47–54. [Google Scholar] [CrossRef]
- Chen, G.D.; Li, Y.J.; Gao, H.; Chen, Y.; Li, X.X.; Li, J.; Guo, L.D.; Cen, Y.Z.; Yao, X.S. New azaphilones and chlorinated phenolic glycosides from Chaetomium elatum with caspase-3 inhibitory activity. Planta Med. 2012, 78, 1683–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Ding, G.; Wu, G.; Yang, J.; Zhang, M.; Wang, H.; Wei, D.; Qin, J.; Guo, L. Identification of a unique azaphilone produced by Chaetomium globosum isolated from Polygonatum sibiricum. Chem. Biodivers. 2020, 17, e1900744. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, J.; Zhu, H.; Wang, J.; Xue, Y.; Wei, G.; Guo, Y.; Tan, D.; Zhang, J.; Yin, C.; et al. Chaephilones A and B, two new azaphilone derivatives isolated from Chaetomium globosum. Chem. Biodivers. 2016, 13, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, J.; Chai, J.; Yu, X.; Zhang, Y.; Feng, Y.; Qin, J.; Yu, H. Chaetomugilin J enhances apoptosis in human ovarian cancer A2780 cells induced by cisplatin through inhibiting pink1/parkin mediated mitophagy. Onco. Targets Ther. 2020, 13, 9967–9976. [Google Scholar] [CrossRef]
- Muroga, Y.; Yamada, T.; Numata, A.; Tanaka, R. 11-and 4-epimers of chaetomugilin a, novel cytostatic metabolit marine fish-derived fungus Chaetomium globosum. Helv. Chim. Acta 2010, 93, 542–549. [Google Scholar] [CrossRef]
- Borges, W.S.; Mancilla, G.; Guimarães, D.O.; Durán-Patrón, R.; Collado, I.G.; Pupo, M.T. Azaphilones from the endophyte Chaetomium globosum. J. Nat. Prod. 2011, 74, 1182–1187. [Google Scholar] [CrossRef]
- Jinno, M.; Muroga, Y.; Yamada, F.; Tanaka, R. Absolute stereostructures of novel azaphiloness produced by a marine fish-derived fungus. Symp. Chem. Nat. Prod. 2009, 51, 563–568. [Google Scholar]
- Yamada, T.; Doi, M.; Shigeta, H.; Muroga, Y.; Hosoe, S.; Numata, A.; Tanaka, R. Absolute stereostructures of cytotoxic metabolites, chaetomugilins A–C, produced by a Chaetomium species separated from a marine fish. Tetrahedron Lett. 2008, 49, 4192–4195. [Google Scholar] [CrossRef]
- Yamada, T.; Muroga, Y.; Doi, M.; Shiget, H.; Numata, A.; Tanaka, R. The structures of the cytotoxic metabolites produced by a marine fish-derived fungus. Symp. Chem. Nat. Prod. 2008, 50, 345–350. [Google Scholar]
- Yasuhide, M.; Yamada, T.; Numata, A.; Tanaka, R. Chaetomugilins, new selectively cytotoxic metabolites, produced by a marine fish derived Chaetomium species. J. Antibiot. 2008, 61, 615–622. [Google Scholar] [CrossRef]
- Yamada, T.; Yasuhide, M.; Shigeta, H.; Numata, A.; Tanaka, R. Absolute stereostructures of chaetomugilins G and H produced by a marine-fish-derived Chaetomium species. J. Antibiot. 2009, 62, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.C.; Zhang, Y.M.; Gao, J.M.; Bai, M.S.; Yang, S.X.; Laatsch, H.; Zhang, A.L. Bioactive metabolites produced by Chaetomium globosum, an endophytic fungus isolated from Ginkgo biloba. Bioorg. Med. Chem. Lett. 2009, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Jinno, M.; Kikuchi, T.; Kajimoto, T.; Numata, A.; Tanaka, R. Three new azaphilones produced by a marine fish derived Chaetomium globosum. J. Antibiot. 2012, 65, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Y.; Ji, X.; Gui, X.; Huang, B.-K. Chemical constituents from an endophytic fungus Chaetomium globosum Z1. Nat. Prod. Commun. 2013, 8, 1217–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, Y.; Li, X.; Pan, H.; Chang, M.; Zheng, T.; Sun, J.; Qiu, D.; Zhang, M.; Wei, D.; et al. Potential allelopathic azaphilones produced by the endophytic Chaetomium globosum TY1 inhabited in Ginkgo biloba using the one strain-many compounds method. Nat. Prod. Res. 2017, 31, 724–728. [Google Scholar] [CrossRef]
- Wang, M.H.; Jiang, T.; Ding, G.; Niu, S.B.; Wang, X.W.; Yu, M.; Gu, Y.C.; Zhang, Q.B.; Chen, J.H.; Jia, H.M.; et al. Molecular epigenetic approach activates silent gene cluster producing dimeric bis-spiro-azaphilones in Chaetomium globosum CBS148.51. J. Antibiot. 2017, 70, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Wang, D.; Yin, X.; Zhang, Q.; Gao, J.-M. New metabolite with inhibitory activity against α-glucosidase and α-amylase from endophytic Chaetomium globosum. Nat. Prod. Commun. 2020, 15, 1–9. [Google Scholar] [CrossRef]
- Yamada, T.; Muroga, Y.; Tanaka, R. New azaphilones, seco-chaetomugilins A and D, produced by a marine-fish-derived Chaetomium globosum. Mar. Drugs 2009, 7, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Selim, K.A.M.A. Biological Evaluation and Chemical Identification of Secondary Metabolites of Endophytic Fungi from Egyptian Flora. Master’s Thesis, Faculty of Science, Cairo University, Cairo, Egypt, 2008. [Google Scholar]
- McMullin, D.R.; Sumarah, M.W.; Blackwell, B.A.; Miller, J.D. New azaphilones from Chaetomium globosum isolated from the built environment. Tetrahedron Lett. 2013, 54, 568–572. [Google Scholar] [CrossRef]
- Piyasena, K.G.N.P.; Wickramarachchi, W.A.R.T.; Kumar, N.S.; Jayasinghe, L.; Fujimoto, Y. Two phytotoxic azaphilone derivatives from Chaetomium globosum, a fungal endophyte isolated from Amaranthus viridis leaves. Mycology 2015, 6, 158–160. [Google Scholar] [CrossRef] [Green Version]
- Rand, T.G.; Chang, C.T.; McMullin, D.R.; Miller, J.D. Inflammation-associated gene expression in RAW 264.7 macrophages induced by toxins from fungi common on damp building materials. Toxicol. In Vitro 2017, 43, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Salo, J.M.; Kedves, O.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Kurnitski, J.; Salonen, H. Detection of Chaetomium globosum, Ch. Cochliodes and Ch. Rectangulare during the diversity tracking of mycotoxin-producing Chaetomium-like isolates obtained in buildings in Finland. Toxins 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y. Cytochalasan and Azaphilone Derivatives from a Marine-Derived Fungus Chaetomium globosum. Master’s Thesis, Seoul National University, Seoul, Korea, 2017. [Google Scholar]
- McMullin, D.R. Structural Characterization of Secondary Metabolites Produced by Fungi Obtained from Damp Canadian Buildings. Ph.D. Thesis, Carleton University, Ottawa, ON, Canada, 2014. [Google Scholar]
- Youn, U.J.; Sripisut, T.; Park, E.J.; Kondratyuk, T.P.; Fatima, N.; Simmons, C.J.; Wall, M.M.; Sun, D.; Pezzuto, J.M.; Chang, L.C. Determination of the absolute configuration of chaetoviridins and other bioactive azaphilones from the endophytic fungus Chaetomium globosum. Bioorg. Med. Chem. Lett. 2015, 25, 4719–4723. [Google Scholar] [CrossRef]
- Kumar, R.; Kundu, A.; Dutta, A.; Saha, S.; Das, A.; Bhowmik, A. Chemo-profiling of bioactive metabolites from Chaetomium globosum for biocontrol of Sclerotinia rot and plant growth promotion. Fungal Biol. 2021, 125, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Muroga, Y.; Yamada, T.; Numata, A.; Tanaka, R. Chaetomugilins I–O, new potent cytotoxic metabolites from a marine-fish-derived Chaetomium species. Stereochemistry and biological activities. Tetrahedron 2009, 65, 7580–7586. [Google Scholar] [CrossRef]
- Yamada, T.; Muroga, Y.; Jinno, M.; Kajimoto, T.; Usami, Y.; Numata, A.; Tanaka, R. New class azaphilone produced by a marine fish derived Chaetomium globosum. The stereochemistry and biological activities. Bioorg. Med. Chem. 2011, 19, 4106–4113. [Google Scholar] [CrossRef]
- Phonkerd, N.; Kanokmedhakul, S.; Kanokmedhakul, K.; Soytong, K.; Prabpai, S.; Kongsearee, P. Bio-spiro-azaphilones and azaphilones from the fungi Chaetomium cochliodes VTh01 and C. cochliodes Cth05. Tetrahedron 2008, 64, 9636–9645. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, G.J.; Jang, K.S.; Lim, H.K.; Kim, H.T.; Cho, K.Y.; Kim, J.C. Antifungal activity against plant pathogenic fungi of chaetoviridins isolated from Chaetomium globosum. FEMS Microbiol. Lett. 2005, 252, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Pornsuriya, C.; Soytong, K.; Poeaim, S.; Kanokmedhakul, S.; Khumkomkhet, P.; Lin, F.-C.; Wang, H.K.; Hyde, K.D. Chaetomium siamense sp. nov., a soil isolate from Thailand, produces a new chaetoviridin, G. Mycotaxon 2011, 115, 19–27. [Google Scholar] [CrossRef]
- Awad, N.E.; Kassem, H.A.; Hamed, M.A.; El-Naggar, M.A.; El-Feky, A.M. Bioassays guided isolation of compounds from Chaetomium globosum. J. Mycol. Med. 2014, 24, e35–e42. [Google Scholar] [CrossRef]
- Xu, G.-B.; Liu, J.; Xiao, X.; Zhou, M.; Yan, T.; Li, G.-Y. Isolation and identification of secondary metabolites from Chaetomium globosum CIB-160 and their immunological activity. Nat. Prod. Res. Dev. 2016, 28, 1562–1567. [Google Scholar]
- Koyama, K.; Takahashi, K.; Natori, S.; Udagawa, S. Production of mycotoxins by Chaetomium species. Proc. Jpn. Assoc. Mycotoxicol. 1991, 33, 40–43. [Google Scholar] [CrossRef]
- Fierro-Cruz, J.E.; Jiménez, P.; Coy-Barrera, E. Fungal endophytes isolated from Protium heptaphyllum and Trattinnickia rhoifolia as antagonists of Fusarium oxysporum. Rev. Argent. Microbiol. 2017, 49, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.M.; Wang, M.; Gong, W.F.; Zhang, L.Q. The screening and identification of the biological control fungi Chaetomium spp. against wheat common root rot. FEMS Microbiol. Lett. 2018, 365, 242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Tan, X.M.; Yu, M.; Yang, J.; Sun, B.D.; Qin, J.C.; Guo, L.P.; Ding, G. Bioactive metabolites from the desert plant-associated endophytic fungus Chaetomium globosum (Chaetomiaceae). Phytochemistry 2021, 185, 112701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, H.; Ye, Y.; Tang, C. Antifungal activity of chaetoviridin A from Chaetomium globosum CEF-082 metabolites against Verticillium dahliae in cotton. Mol. Plant Microbe Interact. 2021, 34, 758–769. [Google Scholar] [CrossRef]
- Qi, J.; Jiang, L.; Zhao, P.; Chen, H.; Jia, X.; Zhao, L.; Dai, H.; Hu, J.; Liu, C.; Shim, S.H.; et al. Chaetoglobosins and azaphilones from Chaetomium globosum associated with Apostichopus japonicus. Appl. Microbiol. Biotechnol. 2020, 104, 1545–1553. [Google Scholar] [CrossRef]
- Wang, W.; Yang, J.; Liao, Y.Y.; Cheng, G.; Chen, J.; Cheng, X.D.; Qin, J.J.; Shao, Z. Cytotoxic nitrogenated azaphilones from the deep-sea-derived fungus Chaetomium globosum MP4-S01-7. J. Nat. Prod. 2020, 83, 1157–1166. [Google Scholar] [CrossRef]
- Li, X.; Tian, Y.; Yang, S.X.; Zhang, Y.M.; Qin, J.C. Cytotoxic azaphilone alkaloids from Chaetomium globosum TY1. Bioorg. Med. Chem. Lett. 2013, 23, 2945–2947. [Google Scholar] [CrossRef]
- Yan, W.; Cao, L.L.; Zhang, Y.Y.; Zhao, R.; Zhao, S.S.; Khan, B.; Ye, Y.H. New metabolites from endophytic fungus Chaetomium globosum CDW7. Molecules 2018, 23, 2873. [Google Scholar] [CrossRef] [Green Version]
- Kingsland, S.R.; Barrow, R.A. Identification of chaetoviridin E from a cultured microfungus, Chaetomium sp. and structural reassignment of chaetoviridins B and D. Aust. J. Chem. 2009, 62, 269–274. [Google Scholar] [CrossRef]
- Steyn, P.S.; Vleggaar, R. The Structure of Dihydrodeoxy-8-epi-austdiol and the Absolute Configuration of the Azaphilones. J. Chem. Soc. Perkin Trans. 1976, 7, 204–206. [Google Scholar] [CrossRef]
- Yasukawa, K.; Takahashi, M.; Natori, S.; Kawai, K.; Yamazaki, M.; Takeuchi, M.; Takido, M. Azaphilones inhibit tumor promotion by 12-O-tetradecanoylphorbol-13-acetate in two-stage carcinogenesis in mice. Oncology 1994, 51, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Mehmet, H. Caspases find a new place to hide. Nature 2000, 403, 29–30. [Google Scholar] [CrossRef]
- Howley, B.; Fearnhead, H.O. Caspases as therapeutic targets. J. Cell. Mol. Med. 2008, 12, 1502–1516. [Google Scholar] [CrossRef] [Green Version]
- Taheri, H.; Filion, K.B.; Windle, S.B.; Reynier, P.; Eisenberg, M.J. Cholesteryl ester transfer protein inhibitors and cardiovascular outcomes: A systematic review and meta-analysis of randomized controlled trials. Cardiology 2020, 145, 236–250. [Google Scholar] [CrossRef]
- Tomoda, H.; Matsushima, C.; Tabata, N.; Namatame, I.; Tanaka, H.; Bamberger, M.J.; Arai, H.; Fukazawa, M.; Inoue, K.; Omura, S. Structure-specific inhibition of cholesteryl ester transfer protein by azaphilones. J. Antibiot. 1999, 52, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Acter, T.; Uddin, N.; Das, J.; Akhter, A.; Choudhury, T.R.; Kim, S. Evolution of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) as coronavirus disease 2019 (COVID-19) pandemic: A global health emergency. Sci. Total Environ. 2020, 730, 138996. [Google Scholar] [CrossRef]
- Abdallah, H.M.; El-Halawany, A.M.; Sirwi, A.; El-Araby, A.M.; Mohamed, G.A.; Ibrahim, S.R.M.; Koshak, A.E.; Asfour, H.Z.; Awan, Z.A.; Elfaky, M.A. Repurposing of some natural product isolates as SARS-COV-2 main protease inhibitors via in vitro cell-free and cell-based antiviral assessments and molecular modeling approaches. Pharmaceuticals 2021, 14, 213. [Google Scholar] [CrossRef]
- Vellingiri, B.K.; Jayaramayya, M.; Iyer, A.; Narayanasamy, V.; Govindasamy, B.; Giridharan, S.; Ganesan, A.; Venugopal, D.; Venkatesan, H.; Ganesan, K.; et al. Subramaniam, COVID-19: A promising cure for the global panic. Sci. Total Environ. 2020, 725, 138277. [Google Scholar] [CrossRef]
- Zarei, A.; Fardood, S.T.; Moradnia, F.; Ramazani, A. A review on coronavirus family persistency and considerations of novel type, COVID-19 features. Eurasian Chem. Commun. 2020, 2, 798–811. [Google Scholar] [CrossRef]
- Anand, K.; Ziebuhr, J.; Wadhwani, P.; Mesters, J.R.; Hilgenfeld, R. Coronavirus main proteinase (3CLpro) structure: Basis for design of anti-SARS drugs. Science 2003, 300, 1763–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandyck, K.; Abdelnabi, R.; Gupta, K.; Jochmans, D.; Jekle, A.; Deval, J.; Misner, D.; Bardiot, D.; Foo, C.S.; Liu, C.; et al. ALG-097111, a potent and selective SARS-CoV-2 3-chymotrypsin-like cysteine protease inhibitor exhibits in vivo efficacy in a Syrian Hamster model. Biochem. Biophys. Res. Commun. 2021, 555, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xie, W.; Xue, X.; Yang, K.; Ma, J.; Liang, W. Design of wide-spectrum inhibitors targeting coronavirus main proteases. PLoS Biol. 2005, 3, e324. [Google Scholar]
- Fiorucci, D.; Milletti, E.; Orofino, F.; Brizzi, A.; Mugnaini, C.; Corelli, F. Computational drug repurposing for the identification of SARS-CoV-2 main protease inhibitors. J. Biomol. Struct. Dyn. 2021, 39, 6242–6248. [Google Scholar] [CrossRef]
- Linnakoski, R.; Reshamwala, D.; Veteli, P.; Cortina-Escribano, M.; Vanhanen, H.; Marjomäki, V. Antiviral agents from fungi: Diversity, mechanisms and potential applications. Front. Microbiol. 2018, 9, 2325. [Google Scholar] [CrossRef] [Green Version]
- Sastry, M.G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Olsson, M.H.M.; Søndergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical pK a Predictions. J. Chem. Theory Comput. 2011, 7, 525–537. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Pavesi, C.; Flon, V.; Mann, S.; Leleu, S.; Prado, S.; Franck, X. Biosynthesis of azaphilones: A review. Nat. Prod. Rep. 2021, 38, 1058–1071. [Google Scholar] [CrossRef]
- Winter, J.M.; Sato, M.; Sugimoto, S.; Chiou, G.; Garg, N.K.; Tang, Y.; Watanabe, K. Identification and characterization of the chaetoviridin and chaetomugilin gene cluster in Chaetomium globosum reveals dual functions of an iterative highly reducing polyketide synthase. J. Am. Chem. Soc. 2012, 134, 17900–17903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, J.M.; Cascio, D.; Dietrich, D.; Sato, M.; Watanabe, K.; Sawaya, M.R.; Vederas, J.C.; Tang, Y. Biochemical and structural basis for controlling chemical modularity in fungal polyketide biosynthesis. J. Am. Chem. Soc. 2015, 137, 9885–9893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.; Winter, J.M.; Kishimoto, S.; Noguchi, H.; Tang, Y.; Watanabe, K. Combinatorial generation of chemical diversity by redox enzymes in chaetoviridin biosynthesis. Org. Lett. 2016, 18, 1446–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Name & Structure | For 6W81 | For 6M2N | For 7K0F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Docking Score | Glide Gscore | Glide Emodel | XP GScore | Docking Score | Glide Gscore | Glide Emodel | XP Gscore | Docking Score | Glide Gscore | Glide Emodel | XP Gscore | |
 | −6.901 | −6.918 | −45.952 | −6.918 | −6.862 | −6.879 | −40.382 | −6.879 | −6.428 | −6.445 | −45.402 | −6.445 |
 | −6.634 | −6.933 | −53.592 | −6.933 | −6.657 | −7.205 | −59.649 | −7.205 | −6.214 | −6.762 | −57.163 | −6.762 |
 | −7.293 | −7.294 | −52.497 | −7.294 | −7.851 | −7.851 | −46.384 | −7.851 | −6.886 | −6.904 | −44.124 | −6.904 |
 | −6.838 | −7.269 | −59.268 | −7.269 | −7.4 | −7.791 | −61.032 | −7.791 | −6.599 | −7.03 | −50.901 | −7.03 |
 | −6.94 | −6.958 | −40.421 | −6.958 | −6.921 | −6.939 | −42.35 | −6.939 | −6.886 | −6.904 | −44.124 | −6.904 |
 | −6.48 | −6.48 | −57.388 | −6.48 | −7.519 | −7.519 | −67.872 | −7.519 | −7.311 | −7.311 | −65.197 | −7.311 |
 | −7.944 | −7.944 | −51.069 | −7.944 | −8.141 | −8.141 | −48.92 | −8.141 | −6.615 | −6.615 | −60.199 | −6.615 |
 Ref_6W81 | −5.377 | −5.377 | −86.001 | −5.377 | ||||||||
 Ref_6M2N | −6.995 | −6.995 | −52.4 | −6.995 | ||||||||
 Ref_K0F | −8.159 | −8.159 | −113.296 | −8.159 | ||||||||
| Fungal Chaetomugilins and Chaetoviridins | Docking Score | Glide Gscore | Glide Emodel | XP Gscore |
|---|---|---|---|---|
| Chaetovirdin D (43) | −7.944 | −7.944 | −51.069 | −7.944 |
| Chaetomugilin P (24) | −7.293 | −7.294 | −52.497 | −7.294 |
| N-Glutrylchaetovirdin A (35) | −7.101 | −7.101 | −66.739 | −7.101 |
| Chaetovirdin B (37) | −6.959 | −6.959 | −37.203 | −6.959 |
| Chaetomugilin R (26) | −6.94 | −6.958 | −40.421 | −6.958 |
| Chaetomugilin 106B-6 XXVIII (1) | −6.901 | −6.918 | −45.952 | −6.918 |
| N-Glutarylchaetovirdin C (42) | −6.881 | −6.881 | −60.87 | −6.881 |
| Chaetomugilin Q (25) | −6.838 | −7.269 | −59.268 | −7.269 |
| Chaetomugilin EA-4 (12) | −6.634 | −6.933 | −53.592 | −6.933 |
| 7,4′,5′-Tris-Epi-chaetovirdin A (34) | −6.519 | −6.519 | −56.68 | −6.519 |
| 4′-Epi-N-2-Hydroxyethyl-azachaetovirdin A (36) | −6.48 | −6.48 | −57.388 | −6.48 |
| Chaetomugilin T (26) | −6.407 | −6.407 | −45.87 | −6.407 |
| Chaetovirdin J (53) | −6.295 | −6.673 | −54.688 | −6.673 |
| 12β-Hydroxychaetovirdin C (41) | −6.074 | −6.08 | −54.03 | −6.08 |
| Chaetomugilin M (21) | −5.985 | −6.143 | −53.589 | −6.143 |
| Chaetovirdin G (50) | −5.925 | −5.925 | −50.419 | −5.925 |
| Chaetomugilin I (16) | −5.919 | −6.467 | −53.609 | −6.467 |
| N-2-Butyric-azochaetovirdin E (46) | −5.862 | −5.863 | −58.955 | −5.863 |
| 11-Epi-chaetomugilin A (4) | −5.771 | −5.774 | −45.965 | −5.774 |
| Chaetovirdin C (40) | −5.752 | −5.752 | −48.221 | −5.752 |
| Chaetomugilin H (15) | −5.741 | −5.741 | −40.295 | −5.741 |
| N-Glutarylchaetovirdin B (39) | −5.733 | −5.735 | −57.615 | −5.735 |
| Chaetomugilin U (30) | −5.622 | −5.622 | −40.195 | −5.622 |
| 4′-Epi-chaetovirdin A (5) | −5.561 | −5.561 | −47.473 | −5.561 |
| Chaetomugilin S (27) | −5.539 | −5.539 | −45.867 | −5.539 |
| 4′,5′-Bis-chaetovirdin A (33) | −5.507 | −5.507 | −48.38 | −5.507 |
| Chaetovirdin A (31) | −5.493 | −5.493 | −51.732 | −5.493 |
| Ref_6W81 | −5.377 | −5.377 | −86.001 | −5.377 |
| Chaetovirdin I (52) | −5.354 | −5.419 | −42.206 | −5.419 |
| Seco-Chaetomugilin D (9) | −5.345 | −5.347 | −45.22 | −5.347 |
| Chaetomugilin L (20) | −5.25 | −5.25 | −39.261 | −5.25 |
| Chaetomugilin B (6) | −5.248 | −5.248 | −41.211 | −5.248 |
| Chaetomugilin A (2) | −5.241 | −5.243 | −44.852 | −5.243 |
| 11-Epi-chaetomugilin I (17) | −5.099 | −5.647 | −54.765 | −5.647 |
| Chaetomugilin D (8) | −5.093 | −5.095 | −41.171 | −5.095 |
| 7-Epi-chaetovirdin E (45) | −5.089 | −5.089 | −40.702 | −5.089 |
| Chaetovirdin K (54) | −4.987 | −4.989 | −45.166 | −4.989 |
| Chaetomugilin N (22) | −4.977 | −4.977 | −3454.68 | −4.977 |
| Seco-chaetomugilin A (3) | −4.926 | −4.928 | −54.168 | −4.928 |
| Chaetomugilin K (19) | −4.903 | −4.903 | −38.494 | −4.903 |
| Chaetovirdin H (51) | −4.898 | −4.898 | −51.406 | −4.898 |
| 4′-Epi-chaetovirdin F (X48) | −4.873 | −4.873 | −47.708 | −4.873 |
| Chaetovirdin B (38) | −4.792 | −4.794 | −42.458 | −4.794 |
| Chaetovirdin E (44) | −4.61 | −4.61 | −48.663 | −4.61 |
| Chaetomugilin G (14) | −4.516 | −4.516 | −50.66 | −4.516 |
| Chaetomugilin S (28) | −4.391 | −4.393 | −48.292 | −4.393 |
| Chaetomugilin F (13) | −4.28 | −4.28 | −40.132 | −4.28 |
| Chaetovirdin F (47) | −4.271 | −4.271 | −50.793 | −4.271 |
| Epi-chaetomugilin D (10) | −4.137 | −4.139 | −36.43 | −4.139 |
| Chaetomugilin E (11) | −3.597 | −3.597 | −40.424 | −3.597 |
| Chaetomugilin C (7) | −3.509 | −3.509 | −44.435 | −3.509 |
| Chaetovirdin G (49) | −3.492 | −3.492 | −48.133 | −3.492 |
| Fungal Chaetomugilins and Chaetoviridins | Docking Score | Glide Gscore | Glide Emodel | XP Gscore |
|---|---|---|---|---|
| Chaetovirdin D (43) | −8.141 | −8.141 | −48.92 | −8.141 |
| Chaetovirdin B (37) | −7.851 | −7.851 | −46.384 | −7.851 |
| 4′-Epi-N-2-Hydroxyethyl-azachaetovirdin A (36) | −7.519 | −7.519 | −67.872 | −7.519 |
| Chaetomugilin Q (25) | −7.4 | −7.791 | −61.032 | −7.791 |
| 12β-Hydroxychaetovirdin C (41) | −7.045 | −7.051 | −58.607 | −7.051 |
| Ref_6M2N | −6.995 | −6.995 | −52.4 | −6.995 |
| Chaetomugilin R (26) | −6.921 | −6.939 | −42.35 | −6.939 |
| Chaetomugilin P (24) | −6.9 | −6.901 | −48.711 | −6.901 |
| Chaetomugilin 106B-6 XXVIII (1) | −6.862 | −6.879 | −40.382 | −6.879 |
| Chaetomugilin S (27) | −6.785 | −6.785 | −50.138 | −6.785 |
| Chaetomugilin EA-4 (12) | −6.657 | −7.205 | −59.649 | −7.205 |
| Chaetomugilin M (21) | −6.655 | −6.813 | −56.59 | −6.813 |
| 7,4′,5′-Tris-Epi-chaetovirdin A (34) | −6.44 | −6.44 | −55.163 | −6.44 |
| Chaetovirdin I (52) | −6.439 | −6.504 | −53.296 | −6.504 |
| Chaetovirdin J (53) | −6.362 | −6.741 | −54.219 | −6.741 |
| Chaetovirdin G (50) | −6.312 | −6.312 | −48.248 | −6.312 |
| Chaetomugilin G (49) | −6.272 | −6.272 | −56.423 | −6.272 |
| Chaetovirdin C (40) | −6.219 | −6.219 | −59.734 | −6.219 |
| N-Glutarylchaetovirdin B (39) | −6.178 | −6.179 | −53.868 | −6.179 |
| 11-Epi-chaetomugilin I (17) | −6.077 | −6.625 | −61.469 | −6.625 |
| Chaetovirdin H (51) | −6.071 | −6.071 | −45.006 | −6.071 |
| Chaetovirdin A (31) | −5.93 | −5.93 | −47.164 | −5.93 |
| 4′,5′-Bis-chaetovirdin A (33) | −5.896 | −5.896 | −46.971 | −5.896 |
| Seco-chaetomugilin A (3) | −5.89 | −5.892 | −44.484 | −5.892 |
| Chaetovirdin E (44) | −5.871 | −5.871 | −48.647 | −5.871 |
| Chaetomugilin I (16) | −5.87 | −6.418 | −55.891 | −6.418 |
| Chaetovirdin F (47) | −5.866 | −5.866 | −54.419 | −5.866 |
| 4′-Epi-chaetovirdin F (48) | −5.859 | −5.859 | −51.775 | −5.859 |
| Chaetomugilin F (13) | −5.759 | −5.759 | −43.54 | −5.759 |
| N-2-Butyric-azochaetovirdin E (46) | −5.453 | −5.454 | −54.064 | −5.454 |
| N-Glutrylchaetovirdin A (35) | −5.405 | −5.405 | −59.359 | −5.405 |
| Seco-Chaetomugilin D (9) | −5.376 | −5.379 | −41.076 | −5.379 |
| Chaetomugilin H (15) | −5.293 | −5.293 | −52.628 | −5.293 |
| Chaetovirdin B (38) | −5.217 | −5.219 | −38.105 | −5.219 |
| 11-Epi-chaetomugilin A (4) | −5.166 | −5.168 | −41.465 | −5.168 |
| Chaetovirdin K (54) | −5.11 | −5.113 | −38.902 | −5.113 |
| Chaetomugilin A (2) | −5.09 | −5.092 | −36.177 | −5.092 |
| Chaetomugilin T (29) | −5.012 | −5.012 | −39.774 | −5.012 |
| Chaetomugilin C (7) | −4.987 | −4.987 | −44.839 | −4.987 |
| 4′-Epi-chaetovirdin A (32) | −4.905 | −4.905 | −58.103 | −4.905 |
| Chaetomugilin S (28) | −4.627 | −4.629 | −36.966 | −4.629 |
| Chaetovirdin G (49) | −4.612 | −4.612 | −42.176 | −4.612 |
| 7-Epi-chaetovirdin E (45) | −4.485 | −4.485 | −51.758 | −4.485 |
| Epi-chaetomugilin D (10) | −4.341 | −4.343 | −31.912 | −4.343 |
| Chaetomugilin U (30) | −4.248 | −4.249 | −40.187 | −4.249 |
| Chaetomugilin O (23) | −4.208 | −4.208 | −1767.6 | −4.208 |
| Chaetomugilin N (22) | −4.07 | −4.07 | −31.442 | −4.07 |
| Chaetomugilin B (6) | −4.068 | −4.068 | −41.99 | −4.068 |
| Chaetomugilin K (19) | −3.867 | −3.867 | −34.284 | −3.867 |
| Chaetomugilin D (8) | −3.667 | −3.669 | −35.613 | −3.669 |
| Chaetomugilin L (20) | −3.622 | −3.622 | −38.371 | −3.622 |
| Chaetomugilin E (11) | −3.506 | −3.506 | −41.116 | −3.506 |
| N-Glutarylchaetovirdin C (42) | −1.412 | −1.412 | −62.951 | −1.412 |
| Fungal Chaetomugilins and Chaetoviridins | Docking Score | Glide Gscore | Glide Emodel | XP Gscore |
|---|---|---|---|---|
| Ref_7K0F | −8.159 | −8.159 | −113.296 | −8.159 |
| 4′-Epi-N-2-Hydroxyethyl-azachaetovirdin A (36) | −7.311 | −7.311 | −65.197 | −7.311 |
| Chaetomugilin R (26) | −6.886 | −6.904 | −44.124 | −6.904 |
| Chaetovirdin D (43) | −6.615 | −6.615 | −60.199 | −6.615 |
| Chaetomugilin Q (25) | −6.599 | −7.03 | −50.901 | −7.03 |
| Chaetomugilin 106B-6 XXVIII (1) | −6.428 | −6.445 | −45.402 | −6.445 |
| Chaetovirdin I (52) | −6.381 | −6.446 | −60.443 | −6.446 |
| Chaetomugilin EA-4 (12) | −6.214 | −6.762 | −57.163 | −6.762 |
| Seco-chaetomugilin A (3) | −6.164 | −6.166 | −54.533 | −6.166 |
| Chaetomugilin T (29) | −6.003 | −6.003 | −44.936 | −6.003 |
| Chaetomugilin A (2) | −5.937 | −5.939 | −49.214 | −5.939 |
| 4′-Epi-chaetovirdin F (48) | −5.781 | −5.781 | −53.249 | −5.781 |
| Chaetovirdin G (50) | −5.73 | −5.73 | −46.915 | −5.73 |
| 11-Epi-chaetomugilin A (4) | −5.634 | −5.636 | −45.502 | −5.636 |
| Chaetomugilin M (21) | −5.56 | −5.718 | −41.62 | −5.718 |
| Chaetovirdin K (54) | −5.521 | −5.523 | −43.983 | −5.523 |
| Chaetovirdin H (51) | −5.487 | −5.487 | −53.211 | −5.487 |
| Chaetovirdin J (53) | −5.475 | −5.853 | −56.559 | −5.853 |
| Chaetomugilin D (8) | −5.457 | −5.459 | −40.705 | −5.459 |
| Chaetomugilin S (28) | −5.409 | −5.411 | −37.945 | −5.411 |
| Chaetovirdin A (31) | −5.356 | −5.356 | −55.812 | −5.356 |
| Chaetomugilin J (18) | −5.262 | −5.758 | −44.768 | −5.758 |
| Chaetovirdin F (47) | −5.214 | −5.214 | −51.016 | −5.214 |
| Chaetovirdin E (44) | −5.207 | −5.207 | −57.958 | −5.207 |
| Chaetomugilin U (30) | −5.17 | −5.17 | −39.245 | −5.17 |
| Chaetomugilin G (14) | −5.154 | −5.154 | −52.872 | −5.154 |
| 4′,5′-Bis-chaetovirdin A (33) | −5.147 | −5.147 | −56.898 | −5.147 |
| Chaetomugilin H (15) | −5.144 | −5.144 | −51.211 | −5.144 |
| Chaetovirdin C (40) | −5.119 | −5.119 | −51.53 | −5.119 |
| Epi-chaetomugilin D (10) | −5.11 | −5.112 | −41.394 | −5.112 |
| Chaetomugilin P (24) | −5.083 | −5.085 | −47.434 | −5.085 |
| Chaetomugilin S (27) | −5.005 | −5.005 | −53.874 | −5.005 |
| N-2-Butyric-azochaetovirdin E (46) | −4.926 | −4.927 | −58.939 | −4.927 |
| 7,4′,5′-Tris-Epi-chaetovirdin A (34) | −4.883 | −4.883 | −50.65 | −4.883 |
| 11-Epi-chaetomugilin I (17) | −4.785 | −5.084 | −57.348 | −5.084 |
| Chaetomugilin B (6) | −4.732 | −4.732 | −44.687 | −4.732 |
| Chaetomugilin I (16) | −4.534 | −4.834 | −53.036 | −4.834 |
| Chaetomugilin C (7) | −4.469 | −4.469 | −45.833 | −4.469 |
| 12β-Hydroxychaetovirdin C (41) | −4.352 | −4.357 | −51.515 | −4.357 |
| Chaetomugilin E (11) | −4.268 | −4.268 | −39.678 | −4.268 |
| N-Glutarylchaetovirdin C (42) | −4.247 | −4.247 | −59.534 | −4.247 |
| Chaetomugilin K (19) | −4.245 | −4.245 | −47.953 | −4.245 |
| Chaetovirdin G (59) | −4.218 | −4.218 | −46.464 | −4.218 |
| Seco-Chaetomugilin D (9) | −4.205 | −4.207 | −48.935 | −4.207 |
| N-Glutrylchaetovirdin A (39) | −4.171 | −4.171 | −55.598 | −4.171 |
| Chaetomugilin F (13) | −3.88 | −3.88 | −42.311 | −3.88 |
| Chaetomugilin R (26) | −3.808 | −5.942 | −41.217 | −5.942 |
| 7-Epi-chaetovirdin E (45) | −3.699 | −3.699 | −46.38 | −3.699 |
| Chaetomugilin L (20) | −3.612 | −3.612 | −43.218 | −3.612 |
| 4′-Epi-chaetovirdin A (32) | −3.552 | −3.552 | −58.379 | −3.552 |
| Chaetomugilin O (23) | −2.951 | −2.951 | 31.679 | −2.951 |
| Chaetovirdin B (38) | −2.642 | −2.644 | −45.702 | −2.644 |
| Chaetomugilin N (22) | −0.921 | −0.921 | −3319.51 | −0.921 |
| N-Glutarylchaetovirdin B (39) | −0.451 | −0.453 | −57.257 | −0.453 |
| Title | Mol MW | #Stars | Dipole | SASA | Donor HB | accptHB | QPlogPo/w | QPlogS | QPlogKhsa | #Metab | QPlogBB | Percent Human Oral Absorption | QPlogHERG | CNS | #rtvFG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Recommended Range | (130–725) | (0.0–5.0) | (1–12.50) | (300–1000) | (0–6) | (2.0–20.0) | (−2–6.5) | (−6.5–0.5) | (−1.5–1.5) | (1–8) | (−3–1.2) | (<25% Poor; >80% High) | Concern Below −5 | (−2 Inactive) (+2 Active) | (0–2) |
| Chaetovirdin K (54) | 450.915 | 0 | 6.534 | 733.38 | 2 | 7.75 | 3.725 | −6.195 | 0.507 | 3 | −0.978 | 100 | −5.072 | −1 | 2 |
| Chaetovirdin J (53) | 408.921 | 0 | 9.823 | 723.174 | 1 | 5.95 | 4.444 | −6.019 | 0.636 | 6 | −1.162 | 100 | −4.971 | −2 | 1 |
| Chaetovirdin I (52) | 466.914 | 0 | 8.017 | 729.53 | 3 | 9.7 | 2.323 | −5.013 | 0.029 | 5 | −1.839 | 77.834 | −4.801 | −2 | 1 |
| Chaetovirdin H (51) | 398.455 | 0 | 10.648 | 693.648 | 0 | 8.2 | 2.632 | −3.929 | −0.18 | 3 | −1.388 | 87.791 | −4.859 | −2 | 0 |
| Chaetovirdin G (50) | 420.932 | 0 | 11.089 | 718.605 | 0 | 6.2 | 4.554 | −5.704 | 0.494 | 4 | −0.639 | 100 | −4.912 | 0 | 1 |
| Chaetovirdin G (49) | 416.9 | 0 | 9.346 | 698.669 | 0 | 4.5 | 4.914 | −6.381 | 0.89 | 5 | −0.882 | 100 | −4.698 | −1 | 1 |
| 4′-Epi-chaetovirdin F (48) | 416.9 | 0 | 11.445 | 723.973 | 0 | 7.5 | 3.722 | −5.295 | 0.155 | 2 | −0.793 | 100 | −5.072 | −1 | 0 |
| Chaetovirdin F (47) | 416.9 | 0 | 11.992 | 704.483 | 0 | 7.5 | 3.502 | −4.926 | 0.089 | 2 | −0.858 | 100 | −4.864 | −1 | 0 |
| N-2-Butyric-azochaetovirdin E (46) | 499.99 | 1 | 13.922 | 827.776 | 1 | 9.5 | 4.264 | −6.466 | 0.317 | 6 | −2.11 | 77.59 | −3.438 | −2 | 1 |
| 7-Epi-chaetovirdin E (45) | 414.885 | 0 | 11.745 | 707.051 | 0 | 7.5 | 3.482 | −5.136 | 0.123 | 3 | −0.816 | 100 | −4.941 | −1 | 1 |
| Chaetovirdin E (44) | 414.885 | 0 | 11.253 | 703.096 | 0 | 7.5 | 3.481 | −5.061 | 0.115 | 3 | −0.776 | 100 | −4.877 | −1 | 1 |
| Chaetovirdin D (43) | 468.93 | 0 | 11.587 | 715.44 | 3 | 8.4 | 3.631 | −4.999 | 0.09 | 5 | −1.572 | 78.707 | −2.816 | −2 | 1 |
| N-Glutarylchaetovirdin C (42) | 572.053 | 0 | 12.347 | 840.463 | 0 | 11.5 | 3.737 | −4.755 | 0.074 | 7 | −1.845 | 73.79 | −4.594 | −2 | 3 |
| 12β-Hydroxychaetovirdin C (41) | 450.915 | 0 | 2.581 | 723.069 | 1 | 6.9 | 3.938 | −5.652 | 0.514 | 6 | −1.528 | 92.976 | −4.818 | −2 | 0 |
| Chaetovirdin C (40) | 434.916 | 1 | 9.724 | 758.733 | 0 | 5.7 | 4.847 | −6.568 | 0.718 | 5 | −1.201 | 100 | −5.436 | −2 | 0 |
| Chaetovirdin C (40) | 434.916 | 0 | 8.19 | 737.905 | 0 | 5.7 | 4.746 | −6.172 | 0.662 | 5 | −1.084 | 100 | −5.134 | −2 | 0 |
| N-Glutarylchaetovirdin B (39) | 544 | 0 | 12.329 | 793.894 | 2 | 11.5 | 3.491 | −5.111 | −0.161 | 7 | −2.718 | 36.417 | −0.961 | −2 | 1 |
| Chaetovirdin B (38) | 434.916 | 0 | 5.891 | 722.451 | 1 | 7 | 4.223 | −6.419 | 0.644 | 3 | −0.608 | 100 | −5.051 | 0 | 2 |
| Chaetovirdin B (37) | 452.931 | 0 | 11.095 | 702.548 | 2 | 6.7 | 4.584 | −5.524 | 0.416 | 4 | −1.112 | 90.886 | −2.679 | −2 | 1 |
| 4′-Epi-N-2-Hydroxyethyl-azachaetovirdin A (36) | 475.968 | 1 | 14.774 | 776.474 | 1 | 9.9 | 3.056 | −5.251 | 0.144 | 6 | −2.061 | 80.997 | −4.863 | −2 | 0 |
| N-Glutrylchaetovirdin A (35) | 604.095 | 0 | 10.792 | 927.971 | 0 | 13.2 | 3.721 | −5.198 | −0.147 | 7 | −2.026 | 76.425 | −5.269 | −2 | 2 |
| 7,4′,5′-Tris-Epi-chaetovirdin A (34) | 432.9 | 0 | 12.018 | 729.829 | 0 | 8.2 | 3.205 | −4.877 | −0.033 | 3 | −1.26 | 92.089 | −5.078 | −2 | 0 |
| 4′,5′-Bis-chaetovirdin A (33) | 432.9 | 0 | 10.874 | 710.174 | 0 | 8.2 | 3.028 | −4.504 | −0.097 | 3 | −1.245 | 90.472 | −4.825 | −2 | 0 |
| 4′-Epi-chaetovirdin A (32) | 432.9 | 1 | 10.365 | 711.04 | 0 | 8.2 | 3.114 | −4.52 | −0.071 | 3 | −1.186 | 91.808 | −4.763 | −2 | 0 |
| Chaetovirdin A (31) | 432.9 | 0 | 11.776 | 730.571 | 0 | 8.2 | 3.204 | −4.891 | −0.033 | 3 | −1.263 | 92.089 | −5.091 | −2 | 0 |
| Chaetomugilin U (30) | 400.471 | 0 | 4.358 | 680.73 | 1 | 7 | 3.638 | −5.427 | 0.501 | 3 | −0.724 | 100 | −4.736 | −1 | 2 |
| Chaetomugilin T (29) | 416.47 | 0 | 5.838 | 708.003 | 2 | 8.7 | 2.784 | −5.198 | 0.232 | 4 | −1.243 | 89.439 | −5.03 | −2 | 2 |
| Chaetomugilin S (28) | 434.916 | 1 | 3.205 | 728.386 | 1 | 7 | 4.315 | −6.524 | 0.659 | 3 | −0.522 | 100 | −5.07 | 0 | 2 |
| Chaetomugilin S (27) | 420.932 | 0 | 10.429 | 713.238 | 0 | 6.2 | 4.401 | −5.611 | 0.469 | 4 | −0.769 | 100 | −4.83 | −1 | 1 |
| Chaetomugilin R (26) | 328.792 | 0 | 6.746 | 578.157 | 3 | 6.9 | 1.802 | −3.7 | −0.18 | 5 | −1.075 | 83.055 | −4.136 | −2 | 0 |
| Chaetomugilin Q (25) | 424.92 | 0 | 10.445 | 738.393 | 2 | 7.65 | 3.474 | −5.495 | 0.295 | 7 | −1.67 | 90.232 | −5.126 | −2 | 1 |
| Chaetomugilin P (24) | 406.905 | 0 | 7.876 | 701.865 | 2 | 6.95 | 3.532 | −5.371 | 0.361 | 7 | −1.223 | 95.462 | −4.879 | −2 | 1 |
| Chaetomugilin O (23) | 416.9 | 1 | 15.406 | 699.173 | 0 | 5.25 | 4.493 | −6.001 | 0.677 | 4 | −0.949 | 100 | −4.786 | −1 | 0 |
| Chaetomugilin N (22) | 432.9 | 1 | 14.072 | 676.058 | 1 | 6.95 | 3.558 | −5.135 | 0.461 | 5 | −1.296 | 90.388 | −4.184 | −2 | 0 |
| Chaetomugilin M (21) | 450.915 | 1 | 12.895 | 720.323 | 1 | 9.9 | 2.623 | −4.609 | −0.057 | 6 | −1.517 | 84.164 | −4.712 | −2 | 2 |
| Chaetomugilin L (20) | 404.933 | 1 | 8.629 | 737.101 | 0 | 4 | 6.159 | −7.539 | 1.184 | 4 | 0.149 | 100 | −5.329 | 1 | 1 |
| Chaetomugilin K (19) | 420.932 | 1 | 6.053 | 732.931 | 1 | 5.7 | 5.186 | −6.78 | 0.887 | 5 | −0.327 | 100 | −5.087 | 0 | 1 |
| Chaetomugilin J (18) | 390.906 | 0 | 7.403 | 711.543 | 1 | 5.25 | 4.761 | −6.404 | 0.766 | 6 | −0.712 | 100 | −5.115 | −1 | 1 |
| 11-Epi-chaetomugilin I (17) | 406.905 | 0 | 8.987 | 705.067 | 2 | 6.95 | 3.831 | −5.476 | 0.389 | 7 | −0.942 | 100 | −4.938 | −1 | 1 |
| Chaetomugilin I (16) | 406.905 | 0 | 8.14 | 722.387 | 2 | 6.95 | 3.744 | −5.781 | 0.415 | 7 | −1.23 | 96.783 | −5.14 | −2 | 1 |
| Chaetomugilin H (15) | 448.942 | 1 | 5.803 | 788.745 | 1 | 6.95 | 4.895 | −7.125 | 0.798 | 4 | −0.859 | 100 | −5.48 | −1 | 0 |
| Chaetomugilin G (14) | 464.942 | 0 | 2.574 | 796.89 | 2 | 8.65 | 3.897 | −6.478 | 0.455 | 5 | −1.373 | 96.768 | −5.491 | −2 | 0 |
| Chaetomugilin F (13) | 416.9 | 0 | 5.154 | 721.74 | 0 | 6.25 | 4.481 | −6.393 | 0.6 | 3 | −0.338 | 100 | −5.109 | 0 | 0 |
| Chaetomugilin EA-4 (12) | 406.905 | 0 | 7.867 | 720.199 | 2 | 6.95 | 3.729 | −5.743 | 0.405 | 7 | −1.214 | 96.92 | −5.132 | −2 | 1 |
| Chaetomugilin E (11) | 448.942 | 0 | 6.895 | 737.896 | 0 | 7 | 4.607 | −6.146 | 0.54 | 3 | −0.279 | 100 | −5.022 | 0 | 2 |
| Epi-chaetomugilin D (10) | 434.916 | 1 | 5.857 | 727.72 | 1 | 7 | 4.222 | −6.512 | 0.645 | 3 | −0.622 | 100 | −5.102 | 0 | 2 |
| Seco-Chaetomugilin D (9) | 466.958 | 1 | 8.323 | 773.281 | 1 | 6.7 | 4.99 | −6.784 | 0.844 | 4 | −0.967 | 100 | −5.112 | −1 | 2 |
| Chaetomugilin D (8) | 434.916 | 1 | 5.813 | 727.059 | 1 | 7 | 4.221 | −6.501 | 0.645 | 3 | −0.621 | 100 | −5.091 | 0 | 2 |
| Chaetomugilin C (7) | 432.9 | 0 | 6.592 | 728.877 | 1 | 7.95 | 3.666 | −6.164 | 0.441 | 4 | −0.828 | 100 | −5.118 | −1 | 0 |
| Chaetomugilin B (6) | 464.942 | 0 | 8.241 | 745.54 | 1 | 8.7 | 3.831 | −6.002 | 0.415 | 4 | −0.756 | 100 | −5.038 | −1 | 2 |
| 11-Epi-chaetomugilin A (4) | 450.915 | 0 | 4.613 | 735.43 | 2 | 8.7 | 3.255 | −5.915 | 0.322 | 4 | −1.091 | 93.153 | −5.126 | −2 | 2 |
| Seco-chaetomugilin A (3) | 482.957 | 0 | 7.988 | 774.455 | 2 | 8.4 | 3.951 | −6.021 | 0.479 | 5 | −1.427 | 95.73 | −5.066 | −2 | 2 |
| Chaetomugilin A (2) | 450.915 | 0 | 4.464 | 734.849 | 2 | 8.7 | 3.266 | −5.905 | 0.335 | 4 | −1.106 | 92.834 | −5.084 | −2 | 2 |
| Chaetomugilin 106B-6 XXVIII (1) | 328.792 | 0 | 6.746 | 578.786 | 3 | 6.9 | 1.79 | −3.71 | −0.186 | 5 | −1.081 | 82.981 | −4.167 | −2 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omar, A.M.; Mohamed, G.A.; Ibrahim, S.R.M. Chaetomugilins and Chaetoviridins—Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics. J. Fungi 2022, 8, 127. https://doi.org/10.3390/jof8020127
Omar AM, Mohamed GA, Ibrahim SRM. Chaetomugilins and Chaetoviridins—Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics. Journal of Fungi. 2022; 8(2):127. https://doi.org/10.3390/jof8020127
Chicago/Turabian StyleOmar, Abdelsattar M., Gamal A. Mohamed, and Sabrin R. M. Ibrahim. 2022. "Chaetomugilins and Chaetoviridins—Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics" Journal of Fungi 8, no. 2: 127. https://doi.org/10.3390/jof8020127
APA StyleOmar, A. M., Mohamed, G. A., & Ibrahim, S. R. M. (2022). Chaetomugilins and Chaetoviridins—Promising Natural Metabolites: Structures, Separation, Characterization, Biosynthesis, Bioactivities, Molecular Docking, and Molecular Dynamics. Journal of Fungi, 8(2), 127. https://doi.org/10.3390/jof8020127