
Bacillus velezensis TSA32-1 as a Promising Agent for Biocontrol of Plant Pathogenic Fungi


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of B. velezensis TSA 32-1
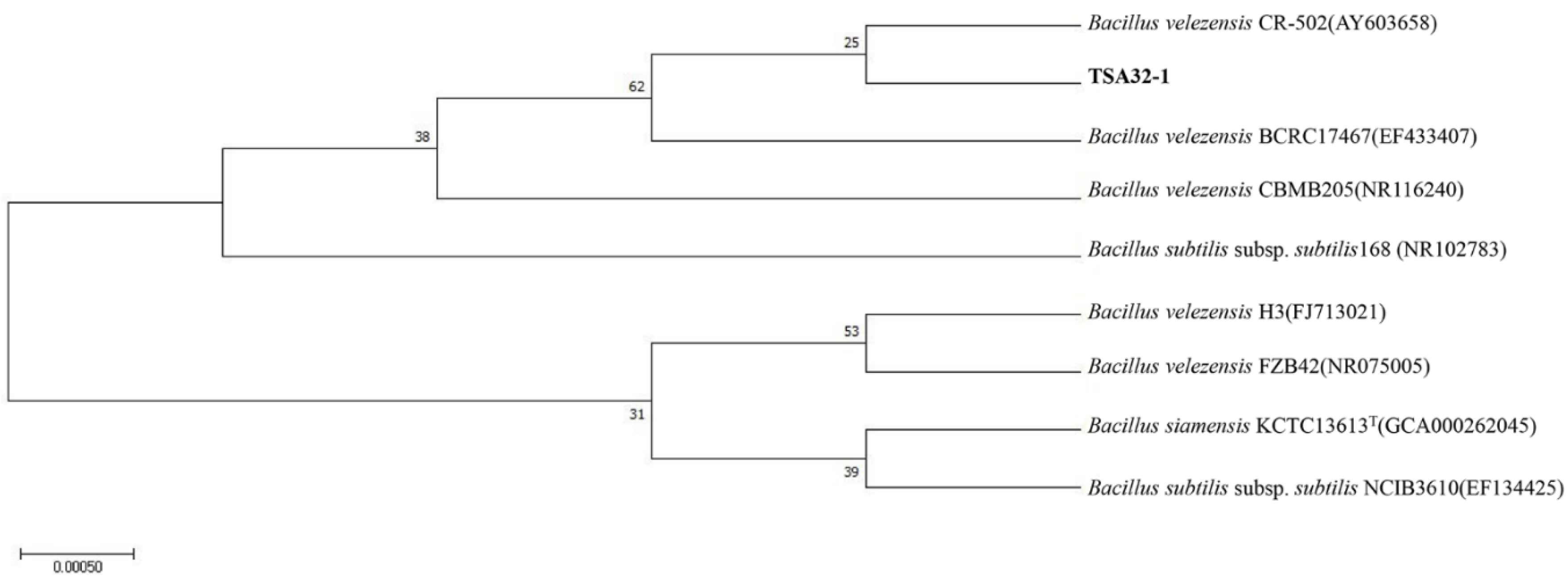
2.2. Phylogenetic and Statistical Analysis
2.3. Enzymatic Activity Analysis
2.3.1. Cellulase Activity
2.3.2. Lysozyme Activity
2.3.3. Protease Activity
2.4. Antifungal Analysis
2.5. Pathogen Inoculum Production and Disease Evaluation
2.5.1. Preparation of Bacillus Strains
2.5.2. Preparation of Seeds
2.5.3. Preparation of Pathogenic Fungus
2.5.4. Seed Germination Rate of Treatments
2.5.5. Disease Suppression Effect of Seed Treatments
2.5.6. DNA Extraction and Whole-Genome Sequencing
2.5.7. Annotation
2.6. Antifungal Metabolite Extraction and Analysis
2.6.1. Preparation of Culture Extract
2.6.2. Ultra-High Performance (UP) LC-QTOF-Mass Spectrometry (MS) Analysis of Metabolites
3. Results
3.1. Identification of Bacterial Strains
3.2. Plant Disease Biocontrol Activity
3.2.1. Enzyme Activity of Isolated Strains
3.2.2. In Vitro Antifungal Effects against Phytopathogenic Fungi
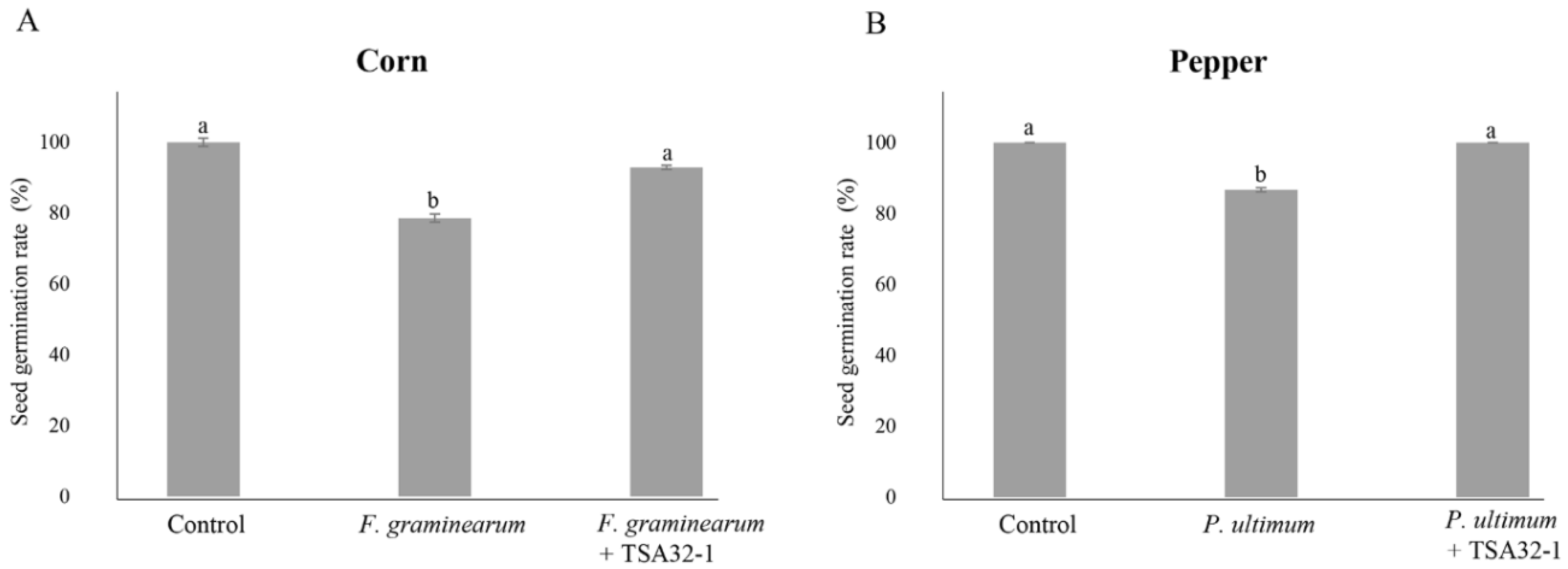
3.2.3. Plant Protection Effects of Seed Treatments with TSA32-1
3.3. Genome Feature of B. velezensis TSA32-1
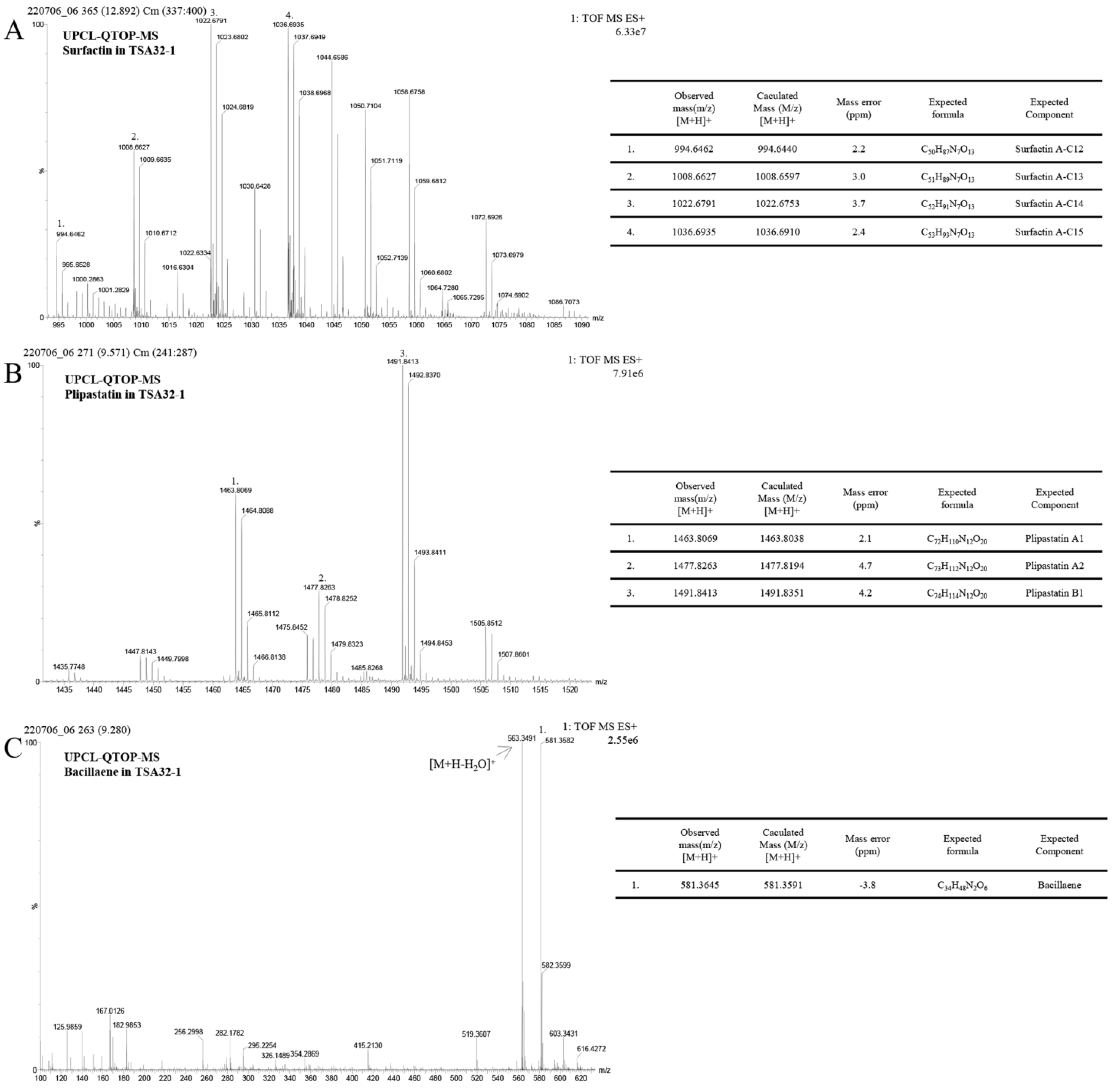
3.4. Antifungal Metabolite of B. velezensis TSA32-1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ji, X.; Ge, Y.; Li, J.; Qi, W.; Qiao, K. Characterization of Antagonistic Bacillus methylotrophicus Isolated From Rhizosphere and Its Biocontrol Effects on Maize Stalk Rot. Phytopathology 2019, 109, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Legein, M.; Smets, W.; Vandenheuvel, D.; Eilers, T.; Muyshondt, B.; Prinsen, E.; Samson, R.; Lebeer, S. Modes of Action of Microbial Biocontrol in the Phyllosphere. Front. Microbiol. 2020, 11, 1619. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, R. Global trends in the crop protection industry. J. Pestic. Sci. 2019, 44, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Muis, S.d.A. Prospect of Bacillus subtilis as a Biological Control Agent of Soilborne Pathogens on Maize. J. Litbang Pert. 2016, 35, 37–45. [Google Scholar]
- Foysal, M.J.; Lisa, A.K. Isolation and characterization of Bacillus sp. strain BC01 from soil displaying potent antagonistic activity against plant and fish pathogenic fungi and bacteria. J. Genet. Eng. Biotechnol. 2018, 16, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, Y.; Cheon, W.; Park, J.; Kwon, H.T.; Balaraju, K.; Kim, J.; Yoon, Y.J.; Jeon, Y. Characterization of Bacillus velezensis AK-0 as a biocontrol agent against apple bitter rot caused by Colletotrichum gloeosporioides. Sci. Rep. 2021, 11, 626. [Google Scholar] [CrossRef]
- Santoso, I.; Fadhilah, Q.G.; Maryanto, A.E.; Yasman. The Potency of Bacillus siamensis LDR as Biocontrol Agent Against Fungal Phytopathogen. Adv. Biol. Sci. Res. 2020, 14, 463–468. [Google Scholar]
- Kuta, F.A. Antifungal effect of Calotropis procera stem bark on Epidermophyton flocosum and Trichophyton gypseum. Afr. J. Biotechnol. 2008, 7, 2116–2118. [Google Scholar]
- Mardanova, A.M.; Fanisovna Hadieva, G.; Tafkilevich Lutfullin, M.; Valer’evna Khilyas, I.; Farvazovna Minnullina, L.; Gadelevna Gilyazeva, A.; Bogomolnaya, L.M.; Sharipova, M.R. Bacillus subtilis Strains with Antifungal Activity against the Phytopathogenic Fungi. Agric. Sci. 2017, 8, 1–20. [Google Scholar]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E.; et al. Antagonistic action of Bacillus subtilis strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef] [PubMed]
- Khabbaz, S.E.; Zhang, L.; Cáceres, L.A.; Sumarah, M.; Wang, A.; Abbasi, P.A. Characterisation of antagonistic Bacillus and Pseudomonas strains for biocontrol potential and suppression of damping-off and root rot diseases. Ann. Appl. Biol. 2015, 166, 456–471. [Google Scholar] [CrossRef]
- Kamali, M.; Guo, D.; Naeimi, S.; Ahmadi, J. Perception of Biocontrol Potential of Bacillus inaquosorum KR2-7 against Tomato Fusarium Wilt through Merging Genome Mining with Chemical Analysis. Biology 2022, 11, 137. [Google Scholar] [CrossRef]
- Li, X.; Munir, S.; Xu, Y.; Wang, Y.; He, Y. Combined mass spectrometry-guided genome mining and virtual screening for acaricidal activity in secondary metabolites of Bacillus velezensis W1. RSC Adv. 2021, 11, 25441–25449. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Son, S.; Kim, J.A.; Jung, M.Y.; Choi, Y.J.; Kim, D.-H.; Lee, H.K.; Shin, D.; Kim, Y. Characterization and Functional Test of Canine Probiotics. Front. Microbiol. 2021, 12, 625562. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.C.; Lai, N.C.; Wu, K.C.; Choi, M.C.; Ma, C.H.; Lin, J.; Kuok, C.-N.; Leong, W.-L.; Lam, W.-K.; Hamied, Y.K.; et al. Safety and Immunogenicity of Inactivated Bacillus subtilis Spores as a Heterologous Antibody Booster for COVID-19 Vaccines. Vaccines 2022, 10, 1014. [Google Scholar] [CrossRef] [PubMed]
- Sauer, D.B.; Burroughs, R. Disinfection of Seed Surfaces with Sodium Hypochlorite. Techniques 1986, 76, 745–749. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Hoboken, NJ, USA, 2008. [Google Scholar]
- Bongiorno, G.; Postma, J.; Bünemann, E.K.; Brussaard, L.; de Goede, R.G.M.; Mäder, P.; Tamm, L.; Thuerig, B. Soil suppressiveness to Pythium ultimum in ten European long-term field experiments and its relation with soil parameters. Soil Biol. Biochem. 2019, 133, 174–187. [Google Scholar] [CrossRef]
- St Arnaud, M.; Hamel, C.; Fortin, J.A. Inhibition of Pythium ultimum in roots and growth substrate of mycorrhizal Tagetes patula colonized with Glomus intraradices. Can. J. Plant Pathol. 2009, 16, 187–194. [Google Scholar] [CrossRef]
- Shang, H.; Chen, J.; Handelsman, J.; Goodman, R.M. Behavior of Pythium torulosum Zoospores During Their Interaction with Tobacco Roots and Bacillus cereus. Curr. Microbol. 1999, 38, 199–204. [Google Scholar] [CrossRef]
- Kaur, K.; Kaur, J.; Puyam, A.; Singh, K. Cultural, Morphological and Molecular Characterization of Fusarium verticillioides causing Maize Ear Rot under Punjab Condition. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 1698–1706. [Google Scholar] [CrossRef]
- Teng, J.L.L.; Yeung, M.L.; Chan, E.; Jia, L.; Lin, C.H.; Huang, Y.; Tse, H.; Wong, S.S.Y.; Sham, P.C.; Lau, S.K.P.; et al. PacBio But Not Illumina Technology Can Achieve Fast, Accurate and Complete Closure of the High GC, Complex Burkholderia pseudomallei Two-Chromosome Genome. Front. Microbiol. 2017, 8, 1448. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, Z.; Fu, L.; Niu, B.; Li, W. WebMGA: A customizable web server for fast metagenomic sequence analysis. BMC Genom. 2011, 12, 444. [Google Scholar] [CrossRef]
- Kim, P.I.; Ryu, J.; Kim, Y.H.; Chi, Y.T. Production of Biosurfactant Lipopeptides Iturin A, Fengycin and Surfactin A from Bacillus subtilis CMB32 for Control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol. 2010, 20, 138–145. [Google Scholar] [CrossRef]
- Guo, S.; Li, X.; He, P.; Ho, H.; Wu, Y.; He, Y. Whole-genome sequencing of Bacillus subtilis XF-1 reveals mechanisms for biological control and multiple beneficial properties in plants. J. Ind. Microbiol. Biotechnol. 2015, 42, 925–937. [Google Scholar] [CrossRef]
- Fonseca, M.C.D.; Bossolani, J.W.; de Oliveira, S.L.; Moretti, L.G.; Portugal, J.R.; Scudeletti, D.; de Oliveira, E.F.; Crusciol, C.A.C. Bacillus subtilis Inoculation Improves Nutrient Uptake and Physiological Activity in Sugarcane under Drought Stress. Microorganisms 2022, 10, 809. [Google Scholar] [CrossRef]
- Poulose, A.J. Biotechnology and fungal control. In Target Sites of Fungicide Action; CRC Press: Boca Raton, FL, USA, 1992; pp. 311–318. [Google Scholar]
- Hernández, C. Chitisan/Carrageenan/Lysozyme particles: Synthesis, Characterization and Antifungal activity against Aspergillus parasiticus. Rev. Mex. Ing. Química 2018, 17, 897–912. [Google Scholar] [CrossRef]
- Kim, J.A.; Bayo, J.; Cha, J.; Choi, Y.J.; Jung, M.Y.; Kim, D.H.; Kim, Y. Investigating the probiotic characteristics of four microbial strains with potential application in feed industry. PLoS ONE 2019, 14, e0218922. [Google Scholar] [CrossRef]
- El-Sersawy, M.M.; Hassan, S.E.; El-Ghamry, A.A.; El-Gwad, A.M.A.; Fouda, A. Implication of plant growth-promoting rhizobacteria of Bacillus spp. as biocontrol agents against wilt disease caused by Fusarium oxysporum Schlecht. in Vicia faba L. Biomol. Concepts 2021, 12, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, B.; Sun, X.; Qi, X.; Zhao, C.; Chang, X.; Khaskheli, M.I.; Gong, G. Symptoms and pathogens diversity of Corn Fusarium sheath rot in Sichuan Province, China. Sci. Rep. 2021, 11, 2835. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.P.; Casal, O.A.; Padin, M.C.; Martinez, V.F.; Oses, M.J.S.; Scauflaire, J.; Munaut, F.; Castro, M.J.B.; Vázquez, J.P.M. First Report of Fusarium temperatum Causing Seedling Blight and Stalk Rot on Maize in Spain. Plant Dis. 2013, 97, 1252. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Dunlap, C.A.; Bowman, M.J.; Chulze, S.N. Bacillus velezensis RC 218 as a biocontrol agent to reduce Fusarium head blight and deoxynivalenol accumulation: Genome sequencing and secondary metabolite cluster profiles. Microbiol. Res. 2016, 192, 30–36. [Google Scholar] [CrossRef]
- Hyder, S.; Gondal, A.S.; Rizvi, Z.F.; Atiq, R.; Haider, M.I.S.; Fatima, N.; Inam-Ul-Haq, M. Biological Control of Chili Damping-Off Disease, Caused by Pythium myriotylum. Front. Microbiol. 2021, 12, 587431. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.-J.M.; Flors, V.; Garcia-Agustin, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting Ready for Battle. Am. Phytopathol. Soc. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Xue, A.G.; Tanbong, J.T. Evaluation of Seed and Soil Treatments with Novel Bacillus subtilis Strains for Control of Soybean Root Rot Caused by Fusarium oxysporum and F. graminearum. Plant Dis. 2009, 93, 1317–1323. [Google Scholar] [CrossRef]
- Kim, K.; Lee, Y.; Ha, A.; Kim, J.I.; Park, A.R.; Yu, N.H.; Son, H.; Choi, G.J.; Park, H.W.; Lee, C.W.; et al. Chemosensitization of Fusarium graminearum to Chemical Fungicides Using Cyclic Lipopeptides Produced by Bacillus amyloliquefaciens Strain JCK-12. Front. Plant Sci. 2017, 8, 2010. [Google Scholar] [CrossRef]
- Leclere, V.; Bechet, M.; Adam, A.; Guez, J.S.; Wathelet, B.; Ongena, M.; Thonart, P.; Gancel, F.; Chollet-Imbert, M.; Jacques, P. Mycosubtilin overproduction by Bacillus subtilis BBG100 enhances the organism’s antagonistic and biocontrol activities. Appl. Environ. Microbiol. 2005, 71, 4577–4584. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.K.; Savard, M.E.; Reid, L.M.; Cyr, T.; McCormick, W.A.; Seguin, C. Identification of lipopeptide antibiotics of a Bacillus subtilis isolate and their control of Fusarium graminearum diseases in maize and wheat. BioControl 2009, 54, 567–574. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Vahidinasab, M.; Lilge, L.; Reinfurt, A.; Pfannstiel, J.; Henkel, M.; Morabbi Heravi, K.; Hausmann, R. Construction and description of a constitutive plipastatin mono-producing Bacillus subtilis. Microb. Cell Fact. 2020, 19, 205. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.F.B.; Gudiña, E.J.; Costa, R.; Vitorino, R.; Teixeira, J.A.; Coutinho, J.A.P.; Rodrigues, L.R. Optimization and characterization of biosurfactant production by Bacillus subtilis isolates towards microbial enhanced oil recovery applications. Fuel 2013, 111, 259–268. [Google Scholar] [CrossRef]
- Harwood, C.R.; Mouillon, J.M.; Pohl, S.; Arnau, J. Secondary metabolite production and the safety of industrially important members of the Bacillus subtilis group. FEMS Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef]
- Aleti, G.; Sessitsch, A.; Brader, G. Genome mining: Prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Comput. Struct. Biotechnol. J. 2015, 13, 192–203. [Google Scholar] [CrossRef]
- Patel, P.S.; Huang, S.; Fisher, S.; Pornik, D.; Aklonis, C.; Dean, L.; Meyers, E.; Fernandes, P.; Mayerl, F. Bacillaene, a Novel Inhibitor of Procaryotic Protein Synthesis Produced by Bacillus subtilis: Production, Taxonomy, Isolation, Physico-chemical Characterization and Biological Activity. J. Antibiot. 1995, 48, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, X.H.; Wu, M.B.; Ge, S. Molecular insights into the antifungal mechanism of bacilysin. J. Mol. Model. 2018, 24, 118. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Zhang, Q.; Yin, L.; Zhang, Y.; Yang, Q.; Liu, K.; Wang, Y.; Han, S.; Zhao, H.; Zhao, H. Isolation and characterization of a mycosubtilin homologue antagonizing Verticillium dahliae produced by Bacillus subtilis strain Z15. PLoS ONE 2022, 17, e0269861. [Google Scholar] [CrossRef] [PubMed]
- Peypoux, F.; Bonmatin, J.M.; Wallach, J. Recent trends in the biochemistry of surfactin. Appl. Microbiol. Biotechnol. 1999, 51, 533–563. [Google Scholar] [CrossRef] [PubMed]
- Moyne, A.L.; Shelby, R.; Cleveland, T.E.; Tuzun, S. Bacillomycin D: An iturin with antifungal activity against Aspergillus flavus. J. Appl. Microbiol. 2001, 90, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Tsalgatidou, P.C.; Thomloudi, E.E.; Baira, E.; Papadimitriou, K.; Skagia, A.; Venieraki, A.; Katinakis, P. Integrated Genomic and Metabolomic Analysis Illuminates Key Secreted Metabolites Produced by the Novel Endophyte Bacillus halotolerans Cal.l.30 Involved in Diverse Biological Control Activities. Microorganisms 2022, 10, 399. [Google Scholar] [CrossRef] [PubMed]
- Fira, D.; Dimkic, I.; Beric, T.; Lozo, J.; Stankovic, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Han, J.; Liu, H.; Qu, X.; Lu, Z.; Bie, X. Plipastatin and surfactin coproduction by Bacillus subtilis pB2-L and their effects on microorganisms. Antonie Van Leeuwenhoek 2017, 110, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Nam, J.; Kim, J.; Song, J.; Kim, P.I.; Min, H.J.; Lee, C.W. Structure and Mechanism of Surfactin Peptide from Bacillus velezensis Antagonistic to Fungi Plant Pathogens. Bull. Korean Chem. Soc. 2019, 40, 704–709. [Google Scholar] [CrossRef]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.-B.; Liao, Y.-C. Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar]
- Chowdhury, S.P.; Dietel, K.; Randler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on lettuce growth and health under pathogen pressure and its impact on the rhizosphere bacterial community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef] [PubMed]
- Falardeau, J.; Wise, C.; Novitsky, L.; Avis, T.J. Ecological and mechanistic insights into the direct and indirect antimicrobial properties of Bacillus subtilis lipopeptides on plant pathogens. J. Chem. Ecol. 2013, 39, 869–878. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Synthetase Type | Genes | Size (kb) | Bioactivity |
|---|---|---|---|---|
| Surfactin | NRPS | srfAA, AB, AC, AD | 26.1 | multiple |
| Bacilysin | NRPS | ywfA, bacA, B, C, D, E, F, ywfH | 6.7 | antibacterial |
| Plipastatin | NRPS | ppsA, ppsC, ppsD, ppsE | 37.6 | antifungal |
| Bacillaene | PKS-NRPS | baeB, C, D, E, acpK, baeG, H, I, J, L, M, N, R, S baeC, L, M, ppsC, baeL, M baeC, G, I, L, M | 71.7 43.3 64.6 | antibacterial |
| Mycosubtilin | PKS-NRPS | mycA, B, C | 35.9 | antifungal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-A.; Song, J.-S.; Kim, P.I.; Kim, D.-H.; Kim, Y. Bacillus velezensis TSA32-1 as a Promising Agent for Biocontrol of Plant Pathogenic Fungi. J. Fungi 2022, 8, 1053. https://doi.org/10.3390/jof8101053
Kim J-A, Song J-S, Kim PI, Kim D-H, Kim Y. Bacillus velezensis TSA32-1 as a Promising Agent for Biocontrol of Plant Pathogenic Fungi. Journal of Fungi. 2022; 8(10):1053. https://doi.org/10.3390/jof8101053
Chicago/Turabian StyleKim, Jung-Ae, Jeong-Sup Song, Pyoung Il Kim, Dae-Hyuk Kim, and Yangseon Kim. 2022. "Bacillus velezensis TSA32-1 as a Promising Agent for Biocontrol of Plant Pathogenic Fungi" Journal of Fungi 8, no. 10: 1053. https://doi.org/10.3390/jof8101053
APA StyleKim, J.-A., Song, J.-S., Kim, P. I., Kim, D.-H., & Kim, Y. (2022). Bacillus velezensis TSA32-1 as a Promising Agent for Biocontrol of Plant Pathogenic Fungi. Journal of Fungi, 8(10), 1053. https://doi.org/10.3390/jof8101053