
Azole-Resistance Development; How the Aspergillus fumigatus Lifecycle Defines the Potential for Adaptation


Abstract
1. Introduction
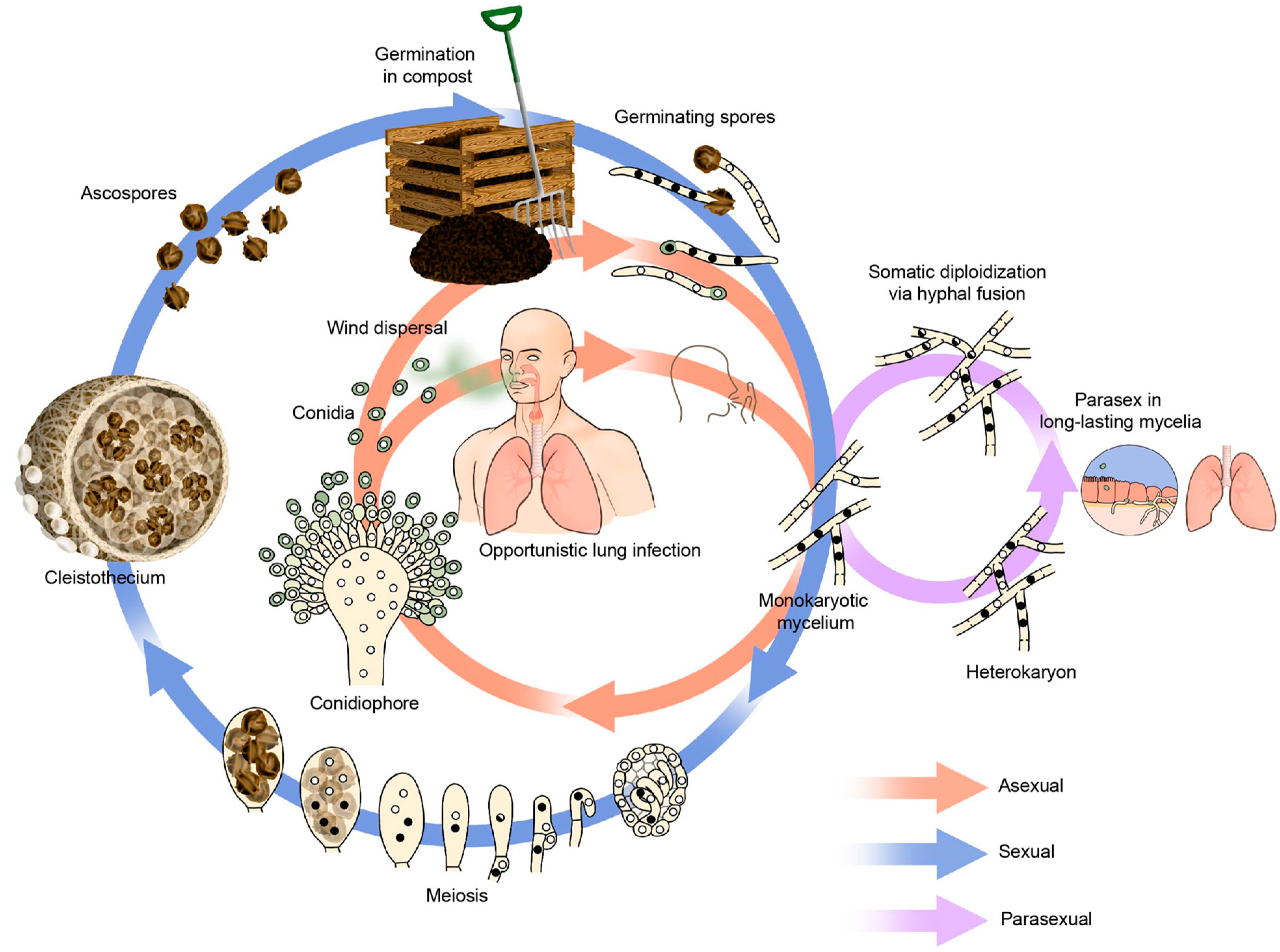
2. A. fumigatus Lifecycle
2.1. Asexual Lifecycle
2.2. Parasexual Lifecycle
2.3. Sexual Lifecycle
3. Mutation and Recombination Contribute to Genetic Variation
3.1. Asexual Genetic Variation
3.2. Parasexual Genetic Varation
3.3. Sexual Genetic Variation
4. Antifungal Resistance Selection and the Role of the A. fumigatus Lifecycle
5. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Latge, J.P. Aspergillus fumigatus and aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [CrossRef]
- Van de Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latge, J.P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of allergic bronchopulmonary aspergillosis with asthma and its complication chronic pulmonary aspergillosis in adults. Med. Mycol. 2013, 51, 361–370. [Google Scholar] [CrossRef]
- Schauwvlieghe, A.; Rijnders, B.J.A.; Philips, N.; Verwijs, R.; Vanderbeke, L.; Van Tienen, C.; Lagrou, K.; Verweij, P.E.; Van de Veerdonk, F.L.; Gommers, D.; et al. Invasive aspergillosis in patients admitted to the intensive care unit with severe influenza: A retrospective cohort study. Lancet Respir. Med. 2018, 6, 782–792. [Google Scholar] [CrossRef]
- Schwartz, I.S.; Friedman, D.Z.P.; Zapernick, L.; Dingle, T.C.; Lee, N.; Sligl, W.; Zelyas, N.; Smith, S.W. High rates of influenza-associated invasive pulmonary aspergillosis may not be universal: A retrospective cohort study from Alberta, Canada. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chong, W.H.; Neu, K.P. The Incidence, Diagnosis, and Outcomes of COVID-19-associated Pulmonary Aspergillosis (CAPA): A Systematic Review. J. Hosp. Infect. 2021. [Google Scholar] [CrossRef]
- Latge, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33. [Google Scholar] [CrossRef]
- Verweij, P.E.; Zhang, J.; Debets, A.J.; Meis, J.F.; van de Veerdonk, F.L.; Schoustra, S.E.; Zwaan, B.J.; Melchers, W.J. In-host adaptation and acquired triazole resistance in Aspergillus fumigatus: A dilemma for clinical management. Lancet Infect. Dis. 2016, 16, 251–260. [Google Scholar] [CrossRef]
- Lestrade, P.P.; Bentvelsen, R.G.; Schauwvlieghe, A.; Schalekamp, S.; van der Velden, W.; Kuiper, E.J.; van Paassen, J.; van der Hoven, B.; van der Lee, H.A.; Melchers, W.J.G.; et al. Voriconazole resistance and mortality in invasive aspergillosis: A multicenter retrospective cohort study. Clin. Infect. Dis. 2019, 68, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Camps, S.M.; Dutilh, B.E.; Arendrup, M.C.; Rijs, A.J.; Snelders, E.; Huynen, M.A.; Verweij, P.E.; Melchers, W.J. Discovery of a HapE mutation that causes azole resistance in Aspergillus fumigatus through whole genome sequencing and sexual crossing. PLoS ONE 2012, 7, e50034. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Arai, T.; Takahashi, H.; Kusuya, Y.; Watanabe, A.; Kamei, K. Non-cyp51A Azole-Resistant Aspergillus fumigatus Isolates with Mutation in HMG-CoA Reductase. Emerg. Infect. Dis. 2018, 24, 1889–1897. [Google Scholar] [CrossRef]
- Zhang, J.; Debets, A.J.; Verweij, P.E.; Melchers, W.J.; Zwaan, B.J.; Schoustra, S.E. Asexual sporulation facilitates adaptation: The emergence of azole resistance in Aspergillus fumigatus. Evolution 2015, 69, 2573–2586. [Google Scholar] [CrossRef]
- Adams, T.H.; Wieser, J.K.; Yu, J.H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Pontecorvo, G. The parasexual cycle in fungi. Annu. Rev. Microbiol. 1956, 10, 393–400. [Google Scholar] [CrossRef]
- Stromnaes, O.; Garber, E.D. Heterocaryosis and the parasexual cycle in Aspergillus fumigatus. Genetics 1963, 48, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Snelders, E.E.; Zwaan, B.J.; Schoustra, S.E.; Kuijper, E.J.; Arendrup, M.C.; Melchers, W.J.G.; Verweij, P.E.; Debets, A.J.M. Relevance of heterokaryosis for adaptation and azole-resistance development in Aspergillus fumigatus. Proc. Biol. Sci. 2019, 286, 20182886. [Google Scholar] [CrossRef] [PubMed]
- Saupe, S.J. Molecular genetics of heterokaryon incompatibility in filamentous ascomycetes. Microbiol. Mol. Biol. Rev. 2000, 64, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Debets, F.; Swart, K.; Hoekstra, R.F.; Bos, C.J. Genetic maps of eight linkage groups of Aspergillus niger based on mitotic mapping. Curr. Genet. 1993, 23, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Verdoes, J.C.; van Diepeningen, A.D.; Punt, P.J.; Debets, A.J.; Stouthamer, A.H.; van den Hondel, C.A. Evaluation of molecular and genetic approaches to generate glucoamylase overproducing strains of Aspergillus niger. J. Biotechnol. 1994, 36, 165–175. [Google Scholar] [CrossRef]
- O’Gorman, C.M.; Fuller, H.; Dyer, P.S. Discovery of a sexual cycle in the opportunistic fungal pathogen Aspergillus fumigatus. Nature 2009, 457, 471–474. [Google Scholar] [CrossRef]
- Bennett, R.J.; Forche, A.; Berman, J. Rapid mechanisms for generating genome diversity: Whole ploidy shifts, aneuploidy, and loss of heterozygosity. Cold Spring Harb. Perspect. Med. 2014, 4. [Google Scholar] [CrossRef]
- Zhang, J.; Snelders, E.; Zwaan, B.J.; Schoustra, S.E.; Meis, J.F.; van Dijk, K.; Hagen, F.; van der Beek, M.T.; Kampinga, G.A.; Zoll, J.; et al. A Novel Environmental Azole Resistance Mutation in Aspergillus fumigatus and a Possible Role of Sexual Reproduction in Its Emergence. mBio 2017, 8. [Google Scholar] [CrossRef]
- Sugui, J.A.; Losada, L.; Wang, W.; Varga, J.; Ngamskulrungroj, P.; Abu-Asab, M.; Chang, Y.C.; O’Gorman, C.M.; Wickes, B.L.; Nierman, W.C.; et al. Identification and characterization of an Aspergillus fumigatus “supermater” pair. mBio 2011, 2. [Google Scholar] [CrossRef]
- Swilaiman, S.S.; O’Gorman, C.M.; Du, W.; Sugui, J.A.; Del Buono, J.; Brock, M.; Kwon-Chung, K.J.; Szakacs, G.; Dyer, P.S. Global sexual fertility in the opportunistic pathogen Aspergillus fumigatus and identification of new supermater strains. J. Fungi 2020, 6, 258. [Google Scholar] [CrossRef]
- Zhu, Y.O.; Siegal, M.L.; Hall, D.W.; Petrov, D.A. Precise estimates of mutation rate and spectrum in yeast. Proc. Natl. Acad. Sci. USA 2014, 111, E2310–E2318. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Snelders, E.; Kema, G.H.; Mellado, E.; Melchers, W.J. Azole resistance in Aspergillus fumigatus: A side-effect of environmental fungicide use? Lancet Infect. Dis. 2009, 9, 789–795. [Google Scholar] [CrossRef]
- Tartof, K.D. Unequal mitotic sister chromatid exchange and disproportionate replication as mechanisms regulating ribosomal RNA gene redundancy. Cold Spring Harb. Symp. Quant. Biol. 1974, 38, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zoll, J.; Engel, T.; van den Heuvel, J.; Verweij, P.E.; Debets, A.J.M. The Medical Triazole Voriconazole Can Select for Tandem Repeat Variations in Azole-Resistant Aspergillus Fumigatus Harboring TR34/L98H Via Asexual Reproduction. J. Fungi 2020, 6, 277. [Google Scholar] [CrossRef] [PubMed]
- Pontecorvo, G. Non-random distribution of multiple mitotic crossing-over among nuclei of heterozygous diploid Aspergillus. Nature 1952, 170, 204–205. [Google Scholar] [CrossRef] [PubMed]
- Pontecorvo, G.; Roper, J.A.; Forbes, E. Genetic recombination without sexual reproduction in Aspergillus niger. J. Gen. Microbiol. 1953, 8, 198–210. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Debets, A.J.; Slakhorst, M.; Hoekstra, R.F. Mitotic recombination accelerates adaptation in the fungus Aspergillus nidulans. PLoS Genet. 2007, 3, 68. [Google Scholar] [CrossRef]
- Engel, T.; Verweij, P.E.; van den Heuvel, J.; Wangmo, D.; Zhang, J.; Debets, A.J.M.; Snelders, E. Parasexual recombination enables Aspergillus fumigatus to persist in cystic fibrosis. ERJ Open Res. 2020, 6. [Google Scholar] [CrossRef]
- Heitman, J. Sexual reproduction and the evolution of microbial pathogens. Curr. Biol. 2006, 16, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Melchers, W.J.; Verweij, P.E. Azole resistance in Aspergillus fumigatus: A new challenge in the management of invasive aspergillosis? Future Microbiol. 2011, 6, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Camps, S.M.; Karawajczyk, A.; Rijs, A.J.; Zoll, J.; Verweij, P.E.; Melchers, W.J. Genotype-phenotype complexity of the TR46/Y121F/T289A cyp51A azole resistance mechanism in Aspergillus fumigatus. Fungal Genet. Biol. 2015, 82, 129–135. [Google Scholar] [CrossRef]
- Snelders, E.; Karawajczyk, A.; Verhoeven, R.J.; Venselaar, H.; Schaftenaar, G.; Verweij, P.E.; Melchers, W.J. The structure-function relationship of the Aspergillus fumigatus cyp51A L98H conversion by site-directed mutagenesis: The mechanism of L98H azole resistance. Fungal Genet. Biol. 2011, 48, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Camps, S.M.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.; Verweij, P.E. Triazole fungicides can induce cross-resistance to medical triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef]
- Hare, R.K.; Gertsen, J.B.; Astvad, K.M.T.; Degn, K.B.; Lokke, A.; Stegger, M.; Andersen, P.S.; Kristensen, L.; Arendrup, M.C. In Vivo selection of a unique tandem repeat mediated Azole resistance mechanism (TR120) in Aspergillus fumigatus cyp51A, Denmark. Emerg. Infect. Dis. 2019, 25, 577–580. [Google Scholar] [CrossRef]
- Risum, M.; Hare, R.K.; Gertsen, J.B.; Kristensen, L.; Johansen, H.K.; Helweg-Larsen, J.; Abou-Chakra, N.; Pressler, T.; Skov, M.; Jensen-Fangel, S.; et al. Azole-Resistant Aspergillus fumigatus Among Danish Cystic Fibrosis Patients: Increasing Prevalence and Dominance of TR34/L98H. Front. Microbiol. 2020, 11, 1850. [Google Scholar] [CrossRef]
- Engel, T.G.P.; Erren, E.; Vanden Driessche, K.S.J.; Melchers, W.J.G.; Reijers, M.H.; Merkus, P.; Verweij, P.E. Aerosol Transmission of Aspergillus fumigatus in Cystic Fibrosis Patients in the Netherlands. Emerg. Infect. Dis. 2019, 25, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, B.; Normand, A.C.; Forel, J.M.; Cassir, N.; Piarroux, R.; Ranque, S. Hospitalized Patient as Source of Aspergillus fumigatus, 2015. Emerg. Infect. Dis. 2018, 24, 1524–1527. [Google Scholar] [CrossRef]
- Pegues, D.A.; Lasker, B.A.; McNeil, M.M.; Hamm, P.M.; Lundal, J.L.; Kubak, B.M. Cluster of cases of invasive aspergillosis in a transplant intensive care unit: Evidence of person-to-person airborne transmission. Clin. Infect. Dis. 2002, 34, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, 383–392. [Google Scholar] [CrossRef]
- Franquet, T.; Muller, N.L.; Gimenez, A.; Guembe, P.; de La Torre, J.; Bague, S. Spectrum of pulmonary aspergillosis: Histologic, clinical and radiologic findings. Radiographics 2001, 21, 825–837. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, S. Biofilm formation by Aspergillus fumigatus. Med. Mycol. 2014, 52, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.A.; Moss, R.B.; Hernandez, C.; Clemons, K.V.; Martinez, M. Effect of Media Modified To Mimic Cystic Fibrosis Sputum on the Susceptibility of Aspergillus fumigatus and the Frequency of Resistance at One Center. Antimicrob. Agents Chemother. 2016, 60, 2180–2184. [Google Scholar] [CrossRef]
- Ashu, E.E.; Hagen, F.; Chowdhary, A.; Meis, J.F.; Xu, J. Global Population Genetic Analysis of Aspergillus fumigatus. mSphere 2017, 2. [Google Scholar] [CrossRef]
- Duarte-Escalante, E.; Zuniga, G.; Ramirez, O.N.; Cordoba, S.; Refojo, N.; Arenas, R.; Delhaes, L.; Reyes-Montes Mdel, R. Population structure and diversity of the pathogenic fungus Aspergillus fumigatus isolated from different sources and geographic origins. Mem. Inst. Oswaldo Cruz 2009, 104, 427–433. [Google Scholar] [CrossRef][Green Version]
- Sewell, T.R.; Zhu, J.; Rhodes, J.; Hagen, F.; Meis, J.F.; Fisher, M.C.; Jombart, T. Nonrandom Distribution of Azole Resistance across the Global Population of Aspergillus fumigatus. mBio 2019, 10. [Google Scholar] [CrossRef]
- Zhou, D.; Korfanty, G.A.; Mo, M.; Wang, R.; Li, X.; Li, H.; Li, S.; Wu, J.Y.; Zhang, K.Q.; Zhang, Y.; et al. Extensive Genetic Diversity and Widespread Azole Resistance in Greenhouse Populations of Aspergillus fumigatus in Yunnan, China. mSphere 2021, 6. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Asexual | Parasexual | Sexual | |
|---|---|---|---|
| Growth type | -Unicellular conidia | Multicellular mycelium | Unicellular conidia Multicellular fruiting body Unicellular ascospores |
| -Multicellular mycelium | |||
| Genetic process | Mitotic mutations and replication slippage or unequal sister chromatid recombination (low frequency 10−5) | Mitotic recombination by crossing over and chromosome reassortment | Meiotic recombination by crossing over and chromosome reassortment |
| Assumed niche | -Lung cavity | Long-lasting mycelia in, for example, lung biofilm | Proposed: composting waste material |
| -Environmental plant waste substrates | |||
| Occurrence | A single spore can form a colony of ~5 cm diameter with up to 109 spores within a week | Diploids form at a rate of ~10−5 and haploidize at a rate of ~10−3 | Sexual reproduction requires mating between isolates of an opposite mating type, ~105–106 ascospores may be formed after ~6 weeks |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Debets, A.J.M.; Verweij, P.E.; Snelders, E. Azole-Resistance Development; How the Aspergillus fumigatus Lifecycle Defines the Potential for Adaptation. J. Fungi 2021, 7, 599. https://doi.org/10.3390/jof7080599
Zhang J, Debets AJM, Verweij PE, Snelders E. Azole-Resistance Development; How the Aspergillus fumigatus Lifecycle Defines the Potential for Adaptation. Journal of Fungi. 2021; 7(8):599. https://doi.org/10.3390/jof7080599
Chicago/Turabian StyleZhang, Jianhua, Alfons J. M. Debets, Paul E. Verweij, and Eveline Snelders. 2021. "Azole-Resistance Development; How the Aspergillus fumigatus Lifecycle Defines the Potential for Adaptation" Journal of Fungi 7, no. 8: 599. https://doi.org/10.3390/jof7080599
APA StyleZhang, J., Debets, A. J. M., Verweij, P. E., & Snelders, E. (2021). Azole-Resistance Development; How the Aspergillus fumigatus Lifecycle Defines the Potential for Adaptation. Journal of Fungi, 7(8), 599. https://doi.org/10.3390/jof7080599