Abstract
Asthma is a chronic heterogeneous respiratory condition that is mainly associated with sensitivity to airborne agents such as pollen, dust mite products and fungi. Key pathological features include increased airway inflammation and airway wall remodelling. In particular, goblet cell hyperplasia, combined with excess mucus secretion, impairs clearance of the inhaled foreign material. Furthermore, structural changes such as subepithelial fibrosis and increased smooth muscle hypertrophy collectively contribute to deteriorating airway function and possibility of exacerbations. Current pharmacological therapies focused on airway wall remodelling are limited, and as such, are an area of unmet clinical need. Sensitisation to the fungus, Aspergillus fumigatus, is associated with enhanced asthma severity, bronchiectasis, and hospitalisation. How Aspergillus fumigatus may drive airway structural changes is unclear, although recent evidence points to a central role of the airway epithelium. This review provides an overview of the airway pathology in patients with asthma and fungal sensitisation, summarises proposed airway epithelial cell–fungal interactions and discusses the initiation of a tissue remodelling response. Related findings from in vivo animal models are included given the limited analysis of airway pathology in patients. Lastly, an important role for Aspergillus fumigatus-derived proteases in triggering a cascade of damage-repair events through upregulation of airway epithelial-derived factors is proposed.
1. Introduction
Asthma is a chronic respiratory condition that develops through a complex interplay of genetic and environmental influences, affecting at least 300 million people worldwide [1]. Symptoms include recurring episodes of persistent cough, wheeze and breathlessness, ultimately leading to poor quality of life [2]. Allergic asthma is characterised by sensitisation to environmental and biologic stimuli, such as pollen, dust mite products, animal dander, tobacco smoke and a wide range of fungi including Aspergillus fumigatus (A. fumigatus) [3,4,5]. Pathological features of asthma include airflow obstruction, due to mucus hypersecretion, and loss of cilia function, bronchial hyperresponsiveness, inflammation and airway remodelling [6]. Airway remodelling comprises of bronchial epithelial denudation, goblet cell hyperplasia and subepithelial fibrosis due to increased extracellular matrix (ECM) deposition [7]. Furthermore, remodelling involves increased smooth muscle mass, vascular remodelling and angiogenesis that correlates with deteriorating airway function [8,9,10]. Goblet cell hyperplasia causes excess mucus production which contributes to trapping foreign airborne particles but can also result in airway narrowing, whilst smooth muscle hypertrophy enhances the intensity of airway reactivity to such irritants. Much of our detailed knowledge with regards to airway remodelling comes from investigating the histopathology of patient bronchial biopsy samples, but these can be difficult to obtain and only provide a snapshot of the disease. High Resolution Computer Topography (HRCT), a useful non-invasive alternative for assessing airway remodelling, has demonstrated bronchial wall thickening and narrowing of the airway lumen in asthmatic lungs that correlates with airflow limitation [11,12,13,14]. Furthermore, HRCT scans have revealed the presence of bronchial obstruction caused by mucus-rich plugs, dilation of bronchi and co-existence of bronchiectasis, a widening of the airways associated with the thickening of the bronchial wall and presence of excess thick mucus [14,15]. In addition to contributing to asthma symptoms, airway remodelling may precede, or occur in parallel with inflammatory changes, making the development of targeted therapeutics to reduce these structural changes an important priority [9,16,17]. Despite this, pharmacological therapies mainly focus on inflammatory targets in asthma patients. Bronchial thermoplasty, an endoscopic procedure that delivers heat treatment to the airway wall to reduce smooth muscle mass, remains one of the few procedures that is proposed to modulate airway remodelling in uncontrolled asthma [18].
Inhalation of airborne components, including airborne conidia, hyphae and fungal fragments, is typically inconsequential for most but for some individuals, dependent on their individual immune status, it may cause a spectrum of disease ranging from sensitisation to severe invasive infection [19]. Intriguingly, the causative fungal species involved in disease may be geographically determined [20]. Furthermore, assessment of respiratory mycobiota in a UK population cohort showed that one of the dominant species present in the airway was A. fumigatus [21]. Another UK-based study found that the respiratory fungal load was much higher in severe asthmatics receiving corticosteroids compared with healthy controls, with A. fumigatus being one of the most prominent species [22]. A. fumigatus can grow both indoors and outdoors with an average adult inhaling several hundred conidia per day and, owing to their small size, they are able to reach the distal alveoli [23]. Following germination and subsequent growth, A. fumigatus produces a multitude of factors, some of which can act as allergens that may either mediate or aggravate asthma symptoms. Moreover, its ability to colonise the respiratory tract, if not readily cleared by inflammatory cells, means it may drive a sustained release of allergens and other products over a prolonged period of time [23]. However, despite strong evidence for an association between A. fumigatus and severe asthma, pathophysiological mechanisms driving an altered airway structure are still being elucidated. Our current understanding of A. fumigatus-associated asthma, fungal–airway epithelial cell interactions and proposed mechanisms leading to airway remodelling are discussed.
2. Aspergillus fumigatus Sensitised Asthma
In healthy individuals, inhaled conidia are removed by the innate immune system through phagocytosis, [24] whereas in immunocompromised patients, or those with reduced lung function, conidia clearance is less effective, allowing germination, prolonged host-allergen exposure and even fungal colonisation with possible invasion [19]. Global estimates indicate that approximately 28% of people with asthma are hypersensitive to A. fumigatus [25]. Furthermore, several studies have demonstrated that fungal sensitisation is associated with increased asthma severity, hospitalisation and even mortality [26,27,28]. A subtype of asthma, identified as ‘Severe Asthma with Fungal Sensitization (SAFS)’, collectively describes a diagnosis of severe uncontrolled asthma, serological or skin prick test detection of fungal sensitisation, and exclusion of Allergic Bronchopulmonary Aspergillosis (ABPA) [29]. In clinical practice, fungal sensitisation is detected by immediate skin hyperreactivity to fungal antigens and/or an increase in IgE antibodies (<1000 IU/mL) [30]. Interestingly, in a cohort of severe asthmatics, 19% showed bronchiectasis associated with poor lung function and frequent exacerbations which was commonly associated with A. fumigatus sensitisation specifically and positive sputum cultures [31]. Findings from another study indicated that 35% of severe asthma patients had central or peripheral bronchiectasis that was again highly associated with A. fumigatus sensitisation and/or sputum- positive cultures [32,33]. Moreover, bronchiectasis patients with persistent Aspergillus cultures were more likely to be hospitalised, have secondary infections, and a more severe lung function decline [34]. Whilst clearly a major health concern, there is limited information regarding histopathological changes of the airways and features of bronchiectasis in the lungs of SAFS patients. Another allergic hypersensitivity disease of the airways that complicates conditions such as asthma and cystic fibrosis and primarily associated with A. fumigatus is ABPA [19,29]. Compared with SAFS, ABPA presents with a more intense inflammatory response [29]. Clinical characteristics of ABPA include lung function deterioration, elevated A. fumigatus specific antibodies, higher than that observed in SAFS (IgE antibodies >1000 IU/mL) and airway eosinophilia [19]. The presence of highly thickened mucus is a hallmark of ABPA and is postulated to facilitate an uncontrolled vicious cycle of fungal growth, antigen production, inflammation and repair and worsening lung pathology [35]. Lung damage frequently localises to the proximal airway region, combined with granulomatous bronchiolitis as well as pulmonary fibrosis [29,36]. Similar to SAFS, bronchiectasis is also a common feature of ABPA, [37] although it remains unclear whether fungi cause or complicate established bronchiectasis, ultimately placing the patient on a pathway to ABPA development [38]. One study using HRCT reported comparable bronchial wall thickening, but a greater prevalence of bronchiectasis in ABPA patients compared with non-ABPA patients [39].
3. Interaction of Airway Epithelium with A. fumigatus
Airway epithelium, lying at the interface of the respiratory system and external environment, acts as one of the first lines of defence against inhaled conidia and hyphal fragments, and plays a central role in innate and adaptive immune responses (reviewed in [40]). Inhaled A. fumigatus conidia adhere to the airways in a number of ways including via exposed ECM components [41] and directly to airway epithelial cells [42], possibly by interaction with integrin receptors [43] and junctional complex protein, E-cadherin [44]. Conidia internalisation can lead to clearance by epithelial cells, but also inflammation and/or epithelial cell death (for review see [45]). It is conceivable that the exposed, damaged, mucus-rich environment of an asthmatic airway is an ideal platform for conidia entrapment, attachment, germination and growth. Interestingly, in cultures of airway epithelial cells, A. fumigatus conidia showed weak adherence to ciliated cells due to the action of cilia beat, but were able to adhere to non-ciliated cells [46]. In an organ culture model with human bronchial mucosal tissue, A. fumigatus conidia mainly adhered to mucus, microscopic indentations of damaged epithelial cells and, to a lesser extent, directly to epithelial cells [47]. FleA, a lectin expressed on the surface of A. fumigatus conidia, binds to purified mucin glycoproteins and so may facilitate this event [48]. Another favoured theory is that damage to and denuding of airway epithelium exposes basal lamina proteins supporting the adhesion of inhaled conidia [49]. Further support for this proposal comes from the fact that A. fumigatus conidia adhere to purified laminin and fibronectin as well as fibrinogen, types I and III collagen in culture [49,50,51,52,53]. Moreover, conidia bind to purified fibrinogen in a dose- and time-dependent fashion in vitro [54]. A. fumigatus also showed superior binding compared with less pathogenic species of Aspergillus, suggesting that adhesion to ECM may be important in disease pathogenesis [55]. Of note, A. fumigatus culture filtrate enhanced conidia–ECM adherence [49], suggesting that secreted fungal factors somehow facilitate binding to the damaged airway.
In addition to acting as a physical barrier, airway epithelium recognises A. fumigatus conidia though expression of pattern recognition receptors (PRRs) such as C-type lectins and toll-like receptors (TLRs), that bind to pathogen associated molecular patterns (PAMPs) to elicit an immune reaction. The outer rodlet layer of inhaled A. fumigatus conidia renders them inert, but the shedding of this layer exposes immunogenic moieties [56]. The type of interaction between A. fumigatus and airway epithelium is dependent on the stage of germination. For instance, swollen or germinating conidia expose β-glucan residues on the surface that interact with the β-glucan receptor, Dectin-1. In addition, bronchial epithelial cells show increased expression of this receptor in response to A. fumigatus [57]. In vivo, loss of Dectin-1 receptor correlates with increased lung damage and cell death in response to A. fumigatus, suggesting that this receptor has a protective role against the effects of fungal exposure [58,59]. Gene silencing in human bronchial epithelial cells showed that Toll-like receptor 2 (TLR2) expression was essential for A. fumigatus induced Dectin-1 expression [57]. Based on observations made using immune cell cultures and animal models, it is likely that several other PAMP–PRR interactions, including those involving soluble receptors such as Pentadextrin 3, facilitate the epithelial response to A. fumigatus (for review see [60]). However, complete mechanistic studies are still needed to fully understand the detailed implications of these relationships.
4. A. fumigatus-Induced Epithelial Cell Damage
The idea that the bronchial epithelium is dysfunctional in asthma, and that injury-induced repair events are activated is widely accepted (for review see [61]). Histopathological analysis revealed areas of denuded ciliated epithelium, as well as goblet cell hyperplasia, as common features of the asthmatic airway [10,62]. In addition, bronchiectasis biopsies showed extensive epithelial hyperplasia, with reduced lung progenitor cells, and an enhanced number of self-renewing cells capable of differentiation into ciliated and secretory cells [63]. Such observations may suggest that aberrant epithelial repair mechanisms contribute to the pathophysiology of asthma with associated bronchiectasis.
As well as interacting directly with epithelial cells through PRRs, A. fumigatus also releases a host of soluble factors, such as allergensand secondary metabolites that can activate epithelial cells but also cause damage and loss of epithelial integrity [64]. The use of A. fumigatus culture filtrates, which are devoid of live spores or hyphae but include soluble secreted factors, are often used in laboratory studies to represent fungal exposure. Indeed, addition of culture filtrate from A. fumigatus to human nasal epithelial cells causes decreased transepithelial resistance, slowed cilial beat frequency and disrupted monolayer integrity [65,66,67,68]. Furthermore, cilial beat frequency reduction and epithelial cell detachment was observed in a human bronchial mucosa organ culture model exposed to germinating A. fumigatus [47,68]. Secondary metabolites, such as gliotoxin and verruculogen, are proposed to contribite to these effects [65,69]. However, A. fumigatus also produces a plethora of allergens, many of which have been cloned and produced as recombinant proteins [70] and more recently assessed as diagnostic tools in SAFS and ABPA [71,72]. Of note, many A. fumigatus allergens show proteolytic activity and include Asp f 5, a fungal matrix metalloproteinase (MMP), Asp f 10 a fungal aspartic protease and Asp f 13 and Asp f 18, fungal serine proteases [73]. Culture filtrates of A. fumigatus cause airway epithelial cell detachment in a dose-dependant fashion [74,75,76,77], and the addition of protease inhibitors, or the use of filtrates with diminished total protease activity, signficantly inhibited this effect [74,75,76]. Furthermore, both germinating A. fumigatus conidia and culture filtrate decreased cytoplasmic actin stress fibres, disrupted focal adhesions and induced membrane blebbing of airway epithelial cells. After 8 h, aggregates of polymerised actin formed in the cytoplasm and by 24 h, the actin cytoskeleton was almost completely depolymerised, the actin mesh disrupted and cell shrinkage and nuclear condensation observed [76,77]. These findings were significantly diminished in response to an Asp f 13 mutant or in the presence of a serine protease inhibitor [76], suggesting that proteases have a major role in epithelial damage and the ensuing pathogical response to A. fumigatus. More recently, purified Asp f 13 was shown to damage the integrity of bronchial epithelial cells grown at an air–liquid interface and the extracellular domains of junctional protein, E-cadherin [78]. The resultant junctional damage and mechanical force was shown to activate the mechanosensor TRPV4 and ultimatly drive airway inflammation and allergic sensitisation [78]. Our previous findings showed that A. fumigatus grown in complex protein growth media composed of homogenised pig lung or mucin resulted in filtrates rich in fungal serine protease and MMP activity, whereas low activity was found when cultured in minimal Vogels media [79]. It is envisaged that the fungus produces proteases to degrade complex proteins, thereby releasing factors required for growth. Indeed, proteases secreted by A. fumigatus have been found to degrade a range of ECM components [80,81,82]. Furthermore, we and others have demonstrated an important role for these fungal proteases in pro-inflammatory cytokine induction, including that of IL-6 and IL-8 [76,83,84,85], [85]. Indeed, when airway epithelial cells were cultured in the presence of germinating conidia from A. fumigatus strains previously shown to be low (Af293) or high (A1160) protease producers [79,85], secreted soluble factors were found to induce the pro-inflammatory cytokine response to strain A1160, which was diminished in the presence of MMP and cysteine protease inhibitors [85]. Taken together, the evidence suggests that A. fumigatus adheres to a range of epithelial and ECM components, activates PRRs, modulates epithelial function and ultimately drives inflammation. How these initial interactions between A. fumigatus and airway epithelium translate to longer-term airway remodelling events are less well understood. This is in part due to the difficulty in obtaining temporal histopathological airway biopsy samples from patients, but also technical issues associated with longer-term in vitro culture of A.fumigatus with epithelial cell for mechanistic studies.
5. A. fumigatus Involvement in Driving Airway Structural Changes
Although histopathological assessment of airway structural changes in lung biopsies from SAFS and ABPA patients is limited, consensus, mainly from CT scans, indicates that their airways show features of an asthmatic airway with extensive remodelling, subepithelial fibrosis and smooth muscle hypertrophy (Figure 1). Findings from murine models of A. fumigatus exposure provide a growing body of evidence to demonstrate that this fungus drives airway inflammation, remodelling and a deterioration of lung function (for review see [86,87]). In vivo models use a range of methods to deliver A. fumigatus to the airways, including conidia, extracts, purified antigens and culture filtrate via intranasal and intratracheal routes or by aerosolisation (Table 1) [88,89,90]. Porter et al. exposed mice to A. fumigatus conidia (unspecified strain) via an intranasal route, three times per week over the course of 18 days, resulting in airway hyperreactivity, eosinophilia and elevated IL-4 and suggested that conidia persist and drive inflammatory responses [90]. Another study exposed mice by aerosol challenge to conidia (strain B-5233/ATCC 13073, initially taken from pulmonary lesions [91]), twice a week for 4 weeks, causing peribronchiolar inflammation and evidence of Th2 sensitisation, subepithelial fibrosis and goblet cell hyperplasia [88]. Intriguingly, these pathological features were not present in mice exposed to melanin-deficient conidia (∆alb1). Silver staining demonstrated that inhaled wild-type conidia germinated within the interstitium, whereas fewer ∆alb1 conidia remained and germinated [88]. Furthermore, Th2 and Th17 sensitisation was only apparent in mice receiving wild-type but not ∆alb1 conidia, demonstrating a crucial role for A. fumigatus melanin in germination and driving a host inflammatory process that contributes to airway pathology.
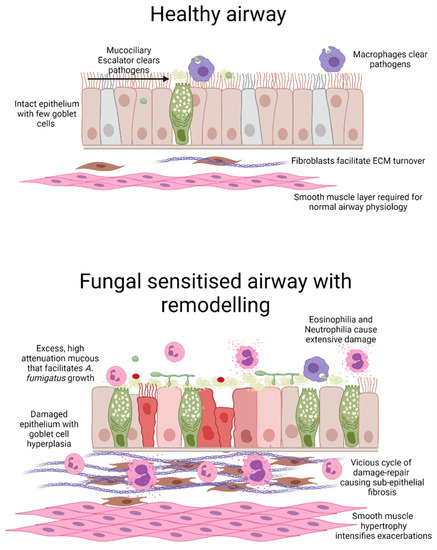
Figure 1.
Schematic representation of A. fumigatus-induced airway wall remodelling. Compared with the healthy airway, a fungal-sensitised asthmatic airway displays extensive inflammation and remodelling. Airway remodelling includes areas of damaged epithelium coupled with goblet cell hyperplasia and excess mucus secretion which likely facilitates fungal growth. Coupled with increased inflammation, activation of fibroblasts and expansion of smooth muscle cells results in airway narrowing with possible luminal obstruction. Image generated in BioRender.
Sustained colonisation of the airways with A. fumigatus conidia, similar to in patients with ABPA, is difficult to model in mice as the use of immunosuppressant agents in rodents can cause rapid invasive disease and mortality [87]. Urb et al. developed a novel solution by delivering A. fumigatus conidia (strain Af293, initially isolated from a neutropenic patient [91]) embedded in agar beads via an intratracheal route [89]. Exposure to conidia or agar beads alone caused no obvious signs of inflammation or airway pathology, whereas hyphal growth was visible in the airway lumen of mice exposed to agar beads with embedded conidia. Fungal growth was associated with significant inflammation, Th2 and Th17 sensitisation and evidence of airway remodelling, although structural changes were not quantified [87]. More recently, Jones et al. challenged mice intranasally with A. fumigatus conidia harvested from a patient with ABPA (strain W72310) seven times over the course of 2 weeks. Relative to mice being challenged with a more widely used strain (CEA10, derived from a patient with invasive aspergillosis [91]), a far greater number of W72310 conidia were found to persist in the lungs at the 14-day time point. This persistence was associated with relatively fewer infiltrating neutrophils and macrophages and a significant reduction in phagocytosis of conidia from the W72310 strain compared with the CEA10 strain [92]. In a separate experiment comparing W72310 strain exposed mice to vehicle control, the ABPA-derived isolate caused IgE induction associated with profound inflammation, goblet cell hyperplasia and increased lavage albumin [92]. Intriguingly, W72310 conidia that persisted in the lungs of mice and caused allergic sensitisation did so without evidence of extensive hyphal growth [92].
Several research groups have developed mouse models involving adjuvant A. fumigatus priming followed by intranasal conidia exposure, which may better model the impact of sustained allergen exposure. For instance, Hogaboam and colleagues gave mice commercially sourced intraperitoneal and subcutaneous injections of soluble A. fumigatus antigen, followed by a weekly intranasal antigen challenge for 3 weeks, and finally an intratracheal instillation of live conidia (strain 13073) [93]. This approach caused significant inflammation of the airways, Th2 sensitisation and airway hyperresponsiveness with many of the features apparent for up to 30 days post-exposure. Of relevance, histopathological analysis revealed significant subepithelial inflammation, goblet cell hyperplasia and subepithelial collagen deposition [93]. In a similar study, Hoselton and colleagues gave a single intraperitoneal and subcutaneous injection with commercially available Aspergillus antigen, followed by a series of intranasal challenges, completed with exposure to aerosolised conidia (unspecified ATCC strain) [94]. This regimen caused significant and even longer lasting Th2 sensitisation and airway hyperreactivity with, again, goblet cell metaplasia, subepithelial thickening with collagen deposition and smooth muscle hypertrophy evident at 35 days post-exposure [94]. To assess the role of B cells in airway remodelling, the same group compared inflammation and tissue pathology using two strains of mice—BALB/c mice that are particularly allergy prone and JH2/2 mice, which do not produce variable antibody heavy chain [95]. Comparable induction of subepithelial collagen deposition and airway hyperreactivity were observed, suggesting that B cells do not drive subepithelial fibrosis. However, mucus production was diminished, whilst eosinophilia, neutrophilia and some markers of Th2 and Th17 sensitisation were enhanced in JH2/2 mice compared with BALB/c mice [95]. These observations suggest disparate pathways may drive inflammation and the various features of airway remodelling in response to A. fumigatus. The role of Th2-inducing cytokines, IL-4 and IL-13, in driving airway pathology has also been assessed in murine models of A. fumigatus exposure. For instance, mice deficient in IL-4 repeatedly exposed to A. fumigatus antigen failed to develop an IgE response but showed comparable lung eosinophilia and lung pathology to that observed in wild-type mice [96]. These observations suggest that factors driving Th2 immunity and airway remodelling may in part be distinct. In another study of murine A. fumigatus antigen sensitisation, IL-13 neutralisation using specific antiserum caused a far greater reduction in airway hyperreactivity associated with a significant reduction in subepithelial fibrosis and goblet cell hyperplasia compared with controls or those receiving IL-4 antiserum [97]. These findings suggest that IL-13 plays an instrumental role in driving multiple features of airway pathology [97].
Taken together, animal models of A. fumigatus-induced airway inflammation demonstrate the ability of the fungus to persist in the airways of immunocompetent mice and drive pathological features, similar to that observed in SAFS and ABPA patients. Whilst these studies have been instrumental in developing a better understanding of disease evolution, the precise cellular mechanisms and the entire array of fungal components responsible remain to be fully described. As outlined in the subsequent section, evidence points to an important role for fungal-derived proteases in mediating A. fumigatus- induced airway wall remodelling.

Table 1.
Findings from selected mouse models of A. fumigatus airway sensitisation.
Table 1.
Findings from selected mouse models of A. fumigatus airway sensitisation.
| Mouse Strain | A. Fumigatus Delivery Method | Key Findings | Ref |
|---|---|---|---|
| A. fumigatus conidia-only models | |||
| C57BL6J | Intranasal delivery of 4 × 105 conidia (isolated from household dust samples), three times per week over 18 days. | Relative to fixed conidia, live conidia caused airway hyperreactivity, eosinophilia, elevated IL-4 and IL-17 levels. Both live and fixed conidia caused modest neutrophilia. | [90] |
| BALB/c | Nose-only aerosol challenge with approximately 1 × 105 conidia (strain B-5233/ATCC 13073) twice per week for 4 weeks. | Histology revealed persistence and germination of wild-type but not melanin-deficient conidia (∆alb1) 48 h post-final dose. Mice exposed to the wild-type but not ∆alb1 strain also displayed eosinophilia, neutrophilia Th2/Th17 sensitisation and evidence of subepithelial fibrosis and goblet cell hyperplasia by histology | [88] |
| C57BL6J | Intratracheal delivery of 2.5 × 106 conidia (strain Af293) embedded in agar beads once. | Non-invasive fungal growth in airway lumen coupled with galactomannan detection. Robust inflammation, including Th2, Th17 and neutrophilia. Severe airway remodelling by histology | [89] |
| C57BL6J | Intranasal delivery of 1 × 107 conidia (strain W72310 from ABPA patient or CEA10) seven times over 2 weeks. | W72310 but not CEA10 conidia persisted in the lung and could be detected as late as 28 days post-final exposure associated with eosinophilia, neutrophilia and Th2 sensitisation. Histology indicates subepithelial fibrosis and goblet cell hyperplasia in response to W72310. | [92] |
| C57BL6J | Intranasal delivery of 4 × 105 conidia (strain Af293), three times per week over 18 days | Evidence of Th2 sensitisation, increased subepithelial fibrosis and epithelial thickening coupled with increased Endothelin-1 levels. | [98] |
| Systemic sensitisation followed by A. fumigatus conidia and/or antigen | |||
| CBA/J | Systemic sensitisation by intraperitoneal and subcutaneous delivery of A. fumigatus antigen in Freund’s adjuvant, followed by a weekly challenge with A. fumigatus antigen for 3 weeks. In week 4, mice received an intratracheal dose of 5 × 106 conidia (strain 13073). | Relative to conidia alone, pre-sensitisation caused profound Th2 sensitisation, profound eosinophilia, neutrophilia and peribronchiolar inflammation. Analysis of histopathology showed that pre-sensitisation also caused goblet cell hyperplasia and subepithelial fibrosis. | [93] |
| BALB/c | Systemic sensitisation by intraperitoneal and subcutaneous delivery of A. fumigatus antigen in alum. After 2 weeks, mice received a weekly intranasal dose of A. fumigatus antigen for 3 weeks and 1 week later, an estimated 6.6 × 105 conidia (strain NIH 5233) were delivered by aerosol | Thickening of the epithelium, goblet cell hyperplasia and airway hyperreactivity persisted for at least 7 days post-final dose. Persistence of Th2 sensitisation and subepithelial fibrosis at the 35-day timepoint. | [94] |
| CBA/J | Systemic sensitisation by intraperitoneal and subcutaneous delivery of A. fumigatus antigen in Alum, followed by a weekly intranasal delivery of A. fumigatus antigen for 3 weeks. Finally, mice received a single intratracheal dose of 5 × 106 conidia (strain not specified). | Upregulation of IL-4 and IL-13. Neutralisation of IL-13, but not IL-4 significantly reduced airway hyperresponsiveness, collagen deposition and subepithelial fibrosis as shown by histology. | [97] |
| BALB/c | Systemic intraperitoneal sensitisation with alum and crude A. fumigatus extract (strain not specified) followed by intranasal delivery of crude extract on days 25–27. In a separate group, mice received crude extract eleven times over the course of 5 weeks. | Alp1/Asp f 13 immunoreactivity visible in the submucosa of A. fumigatus sensitised mice. | [99] |
| A. fumigatus extract or culture filtrate models | |||
| BALB/c and C57BL/6 derived genetically altered | Intranasal delivery of A. fumigatus extract (strain not specified), heat-inactivated extract or purified Alp1/Asp f 13, three times per week for 2 weeks. | Compared to mice receiving A. fumigatus extract or purified Alp1, those exposed to heat inactivated extract or Alp 1 showed diminished airway hyperreactivity, Th2 sensitisation, neutrophilia, peribronchiolar inflammation and goblet cell hyperplasia. Eosinophil-deficient and PAR2-deficient mice developed comparable inflammation, neutrophilia, Th2 sensitisation and goblet cell hyperplasia in response to Alp 1 to that found in wild-type mice. | [100] |
| BALB/c | Intranasal delivery of A. fumigatus sterilised and dialysed culture filtrate (CEA10 derived and protease allergen-deficient strains), twice per week for 4 weeks. | Neutrophilia, eosinophilia and Th2 sensitisation coupled with airway hyperreactivity and remodelling. Exposure to culture filtrates lacking protease allergens, Asp f 5 or Asp f 13, significantly reduced the extent of airway wall remodelling | [101] |
| C57BL6J | Intranasal delivery of A. fumigatus sterilised and dialysed culture filtrates (strain Af293), twice per week for 4 weeks. | Extensive inflammation and Th2 sensitisation in parallel with extensive subepithelial fibrosis. Endothelin-1 receptor antagonism prevented A. fumigatus-induced airway wall remodelling. | [98] |
6. Involvement of A. fumigatus-Derived Proteases in Airway Wall Remodelling
Airway wall remodelling is a key feature of the fungal-sensitised asthmatic lung and demonstrated in vivo following prolonged A. fumigatus exposure (Table 1). Cellular and molecular mechanisms driving these structural changes are not fully understood and require further investigation, however, fungal-derived proteases produced during germination and growth are highly likely to be involved. Through using protease inhibitors and protease gene-deficient condidia, A. fumigatus-derived proteases have been shown to induce airway epithelial cell disruption with release of pro-inflammatory cytokines in addition to driving mucus production. Mucus hypersecretion may support the growth of inhaled conidia in the asthmatic lung and allow extended secretion of proteolytically active allergens resulting in a cycle of injury-repair with subsequent loss of epithelial integrity and long-term alterations in airway structure. Indeed, mucin-enriched culture media supported the production of A. fumigatus proteases [79] and such proteases have been found to upregulate mucin gene expression, MUC5B and MUC5AC [102], suggesting that a continual feedback loop may be in operation. Others have found that Asp f 13 induced smooth muscle contraction in a murine precision-cut lung slice model [100] and upregulated production of host MMPs [99]. Interestingly, mice receiving intraperitoneal sensitisation followed by intranasal challenge with A. fumigatus antigen showed positive immunostaining for Asp f 13 within bronchiole smooth muscle cells [103]. Such an observation mirrors clinical findings whereby Asp f 13, immunoreactivity was localised in the epithelium, mucus layer and smooth muscle of airway biopsies from asthmatic patients, in particular those patients with A. fumigatus sensitisation [99,103]. Our previous work demonstrated that A. fumigatus (derived from strain CEA10) produced high protease activity even in Vogels minimal media [101]. When mice were exposed intranasally to these protease-rich culture filtrates, they developed airway hyperreactivity, Th2 sensitisation and airway remodelling characterised by subepithelial fibrosis and goblet cell hyperplasia [101]. Of interest, exposure to culture filtrates derived from mutant A. fumigatus strains lacking Asp f 5 or Asp f 13, only partially reduced airway inflammation, but significantly reduced features of airway remodelling [101]. Further evidence that fungal proteases drive structural changes comes from a related study, whereby mice were intranasally exposed to A. fumigatus extract or purified Asp f 13 repeatedly over a two week period [100]. Both heat inactivation or the use of a serine protease inhibitor successfully prevented the deterioration of airway function, reduced lung lavage Th2 cytokines and infiltrating immune cells as well as goblet cell hyperplasia [100]. Intriguingly, Asp f 13 directly elicited airway smooth muscle contraction, making targetted antagonism of fungal protease an attractive treatment option for A. fumigatus-sensitised asthma [100].
Endogenous host serine proteases, such as tryptase and thrombin, are known to activate lung epithelial cells through protease-activated receptors (PARs) via cleavage of the N terminus of the receptor [104,105]. PAR activation causes cytokine release as well as vasodilation, platelet aggregation, cellular proliferation and smooth muscle contraction [106,107,108]. PAR-2 is the most prominent PAR in allergic airway disease with increased expression on the surface of the epithelial cells of the bronchi in people with asthma [109]. Findings suggest that A. fumigatus proteases can also activate PAR-2 to drive inflammation and induce an increase in IgE as well as IL-6, IL-8, exotaxin, GM-CSF and MMP9 [110]. However, a role for A. fumigatus-derived proteases in activating PARs to mediate airway remodelling remains unclear. Suprisingly, compared with wild-type mice, PAR-2 knockout mice exposed to Asp f 13 showed comparable airway hyperreactivty and goblet cell hyperplasia, and demonstrated a modest decrease in lung inflammatory cells and inflammatory cytokines [100], which may suggest that fungal proteases drive pathology through multiple mechanisms. In vitro, human airway epithelial cells showed an upregulation of mucin genes, MUC5AC and MUC5B, in response to A. fumigatus extract which was blocked in the presence of a serine protease inhibitor but not a PAR-2 antibody [111]. Rather, A. fumigatus proteolytic activity was shown to drive the phosphorylation/activation of Ras/Raf1/ERK to induce mucin production suggesting involvement of fungal proteases but independent of PAR activation [111]. Taken together, these observations indicate an important role for A. fumigatus-derived proteases in epithelial damage and aspects of airway remodelling which ultimatly leads to poor lung function.
7. A. fumigatus-Induced Airway Epithelial-Derived Profibrogenic Factor Production
It is speculated that cross-communication between airway epithelium and underlying mesenchymal cells, coined the ‘Epithelial-mesenchymal trophic unit (EMTU)’ shapes the way that the airway architecture changes in response to inhaled airborne agents [112]. Accordingly, the airway epithelium of asthmatic patients may respond to such exposure by secreting growth factors, which then drive an upregulation of ECM components [113]. In addition, evidence from genetically altered mice and inhibitor blocking studies suggest that these growth factors induce subepithelial fibrosis, which is linked to asthma severity [114,115,116,117,118,119]. In this regard, little is known about the ability of A. fumigatus to induce the production of fibrogenic growth factors by airway epithelial cells. Findings suggest that cultured murine lung epithelial cells exposed to both live and heat-inactivated A. fumigatus conidia induce an upregulation of epithelial-derived cytokines, such as thymic stromal lymphopoietin (TSLP), IL-25 and IL-33, that promote a Th2-type response, [120]. In a murine model of chronic multiple allergen exposure (house dust mite, ragweed, and A. fumigatus extract), treatment with anti-TSLP antibody reduced perivascular inflammation, reduced goblet cell hyperplasia and improved lung function [121] although remodelling of the airways was not analysed. Further studies investigating the role of TSLP, IL-25 and IL-33 in airway remodelling elicited by A. fumigatus are therefore required. Findings from another study showed that exposure to commercially sourced A. fumigatus antigens caused significant upregulation of peribronchiolar periostin, a matricellular protein, and features of airway remodelling, but mice deficient in periostin did not show any reduction of these features suggesting that periostin may not be a key mediator of airway remodelling [122]. We assessed the induction of key profibrogenic growth factors, TGFβ1, TGFβ2, periostin and Endothelin-1 by primary airway epithelial cells exposed to A. fumigatus conidia (strain Af293) or culture filtrate and found only a highly selective upregulation of Endothelin-1 [98]. Mice intranasally exposed to conidia or repeatedly with culture filtrate also displayed a highly selective upregulation of Endothelin-1. Furthermore, culture filtrate-exposed mice showed significant inflammation with Th2 sensitisation coupled with subepithelial fibrosis, whereas interestingly, antagonism of the Endothelin-1 receptor diminished this response [98]. How Endothelin-1 drives A. fumigatus-induced airway structural changes and the importance of Endothelin-1 in patients with A. fumigatus sensitised asthma still needs to be addressed (Figure 2).
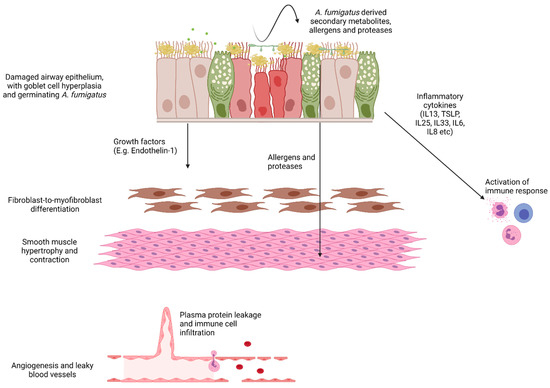
Figure 2.
Schematic demonstrating possible pathways to A. fumigatus-induced airway remodelling. The asthmatic epithelium is characterised by goblet cell hyperplasia and mucus hypersecretion, loss of ciliated cells and subepithelial fibrosis. This fibrosis is likely driven by injury signals from the epithelium activating underlying fibroblasts. Loss of cilia function coupled with increased mucus and exposure of basement membrane components may enhance A. fumigatus adhesion and allergen production. A. fumigatus-derived factors drive the upregulation of pro-inflammatory cytokines to shape the immune response. In addition, epithelial-derived growth factors such as Endothelin-1 are upregulated in response to A. fumigatus and may directly activate underlying fibroblasts. Angiogenesis and vascular permeability support the arrival of infiltrating immune cells and circulating mediators likely contribute to airway wall remodelling. Image generated in BioRender.
Vascular leakiness is a key pathophysiological feature of asthma that correlates with induction of Vascular Endothelial Growth Factor (VEGF) and occurs in advance of inflammation in patients with deteriorating lung function [123,124]. Of importance, Endothelin-1 also promotes vascular permeability [125] and is upregulated in human asthmatic airways [126] as well as in an A. fumigatus-exposed airway mouse model [98]. Taken together, upregulation of VEGF and Endothelin-1 in the asthmatic airway may mediate vascular leak and facilitate the deposition of plasma proteins such as fibrin (Figure 2). Dysregulation of the fibrinolytic pathway in severe asthma patients treated with corticosteroid also favours fibrin deposition [127]. Indeed, the airway lumen of fatal asthmatics shows positive immunostaining for fibrin [128] and mucus plugs from airways of ABPA patients are rich in fibrin [129]. However, whether A. fumigatus interacts with fibrin to drive airway pathology remains to be elucidated. Of significance, co-administration of fibrinogen and thrombin were sufficient in driving airway disease in mice. The fibrinolytic enzyme, tissue plasminogen activator (tPA), also reduced airway reactivity [128], implying that coagulation and fibrinolytic pathways may contribute to airway remodelling and hyperreactivity. Of note, fibrin and Endothelin-1 are both known to mediate fibroblast activation and collagen deposition [130,131] supporting this concept.
In vitro, fibrinogen cleavage products are produced by incubation with proteases derived from another Aspergillus species, A. oryzae or thrombin (serine protease that converts fibrinogen to fibrin). These cleavage products can activate bone marrow-derived macrophages via TLR4 and support subsequent fungicidal activity to conidia, but may also contribute to the initiation or perpetuation of airway pathology [132]. It is plausible that A. fumigatus proteases may also cleave fibrinogen, producing by-products that activate PRRs. In another experiment, mice exposed to Aspergillus oryzae protease were compared to those receiving fibrinogen cleavage products. Results showed that cleavage products alone are not sufficient to drive airway disease to the extent observed with fungal protease. However, inhibition of thrombin in fungal protease exposed mice, attenuated all features of airway disease in this murine model, suggesting that the fibrinolytic pathway or degradation products play an important role in the mechanism of disease progression [132].
Taken together, fungal proteases may generate fibrin cleavage products in a similar manner to host proteases and these products may play a central role in driving both airway inflammation and airway remodelling. Linked to the concept that reduced fibrinolysis is involved in A.fumgatus airway effects, Asp f 2, a major A. fumigatus allergen and emerging biomarker for ABPA and SAFS [71,72], was recently shown to bind to plasminogen [133]. Plasmin, the active component of plasminogen, is a major fibrinolytic protease involved in fibrin matrix removal. In vitro, airway epithelial cells exposed to plasminogen-coated conidia in the presence of a plasminogen activator showed far greater epithelial damage than cells treated with conidia alone or conidia that lacked Asp f 2 expression [133]. The actions of plasminogen-coated conidia were blocked using a serine protease inhibitor suggesting that an interplay between the major allergen Asp f 2 and fibrinolytic protease activity is involved in inducing airway epithelial damage [133].
8. Conclusions
Taken together, the findings strongly suggest that the initial interaction between A. fumigatus and the airway epithelium is key to subsequent inflammation and damage-repair responses. In the asthmatic milieu, an environment rich in mucin, exposed matrix components and plasma proteins likely provides the ideal growth conditions for inhaled A. fumigatus. Through the course of germination, a wide range of allergens, some with protease activity, and metabolic by-products are produced which directly disrupt the integrity of the epithelium and elicit the productionof proinflammatory cytokines and fibrogenic growth factors. The ensuing recruitment of immune cells and further leakage of plasma proteins would support the development of a vicious cycle of inflammation, fibrin deposition and structural changes. A. fumigatus allergens with protease activity seem to be particularly important in disrupting the integrity of the epithelium, driving mucin production, subepithelial fibrosis, and hyperactivity of smooth muscle cells. Understanding the relative contribution of individual A. fumigatus-derived proteases in mediating these pathological events resulting in airway remodelling is now required. Future studies that quantify features of airway wall remodelling and the development of bronchiectasis in animal models would improve our knowledge of disease pathogenesis. Moreover, detailed analysis of the epithelium, subepithelial fibrosis and bronchiectasis in A. fumigatus-sensitised patients is also an important area for future investigation. As highlighted, a range of in vitro and in vivo models have been developed to investigate A. fumigatus- induced airway changes which have included using a wide variety of cell types (cell lines, primary cells, nasal, bronchial, alveolar), culture methods (submerged, air-liquid interface), different mouse strains, dosing regimens and fungal components (conidia/commercial extract/culture filtrate) from many A. fumigatus isolates. This variability in study design is likely to introduce heterogeneity in findings which should be considered, and a possible standardisation approach explored [91]. Lastly, although the current review has focused on the contributions of A. fumigatus to airway epithelial damage and subsequent pathology, it is recognised that a multitude of foreign agents are inhaled simultaneously, and the consequence of combined exposure is an area for further research. In particular, the important contribution of other fungi found to be associated with asthmashould not be overlooked. For instance, one study detected raised specific IgE levels against Aspergillus, Candida and Trichophyton in asthma patients, although some others such as Cladosporium, Penicillium and Schizophyllum commune were also observed [4]. In other patient cohorts, Alternaria and Cladosporium appeared to be especially frequent in patients with asthma and fungal sensitisation [5,134]. Detailed mechanistic studies investigating whether these other fungi and the factors they produce, can activate and damage the airway epithelium are limited [135,136]. Furthermore, emerging evidence points to an important role for sensation to a range of fungal allergens in the skin of atopic dermatitis patients, in particular those derived from Alternaria and Malassezia, Aspergillus, Saccharomyces and Cladosporium [135,137]. Such allergens may contribute to the process of ‘atopic march’, where sensitisation in the skin during infancy leads to subsequent development of allergic disease at other sites, including the airways. Indeed, higher frequencies of severe asthma and rhinitis are seen in atopic dermatitis patients with allergy to Alternaria, Saccharomyces and Cladosporium [137]. Taken together, fungal sensitisation is unlikely to be limited to one organism at a single body site and further large multi-centre global studies comparing the lung mycoflora and allergies, coupled with detailed in vitro and in vivo mechanistic studies, are required.
Author Contributions
Conceptualization, S.N. and S.H.; writing S.N., B.L., J.R. and S.H. Review and editing, S.N., B.L. and S.H.; funding acquisition, S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Medical Research Council grant number [MR/K017497/1].
Conflicts of Interest
The authors declare no conflict of interest.
References
- Loftus, P.A.; Wise, S.K. Epidemiology and economic burden of asthma. Int. Forum Allergy Rhinol. 2015, 5 (Suppl. S1), S7–S10. [Google Scholar] [CrossRef] [PubMed]
- Mims, J.W. Asthma: Definitions and pathophysiology. Int. Forum Allergy Rhinol. 2015, 5 (Suppl. S1), S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Schatz, M.; Rosenwasser, L. The allergic asthma phenotype. J. Allergy Clin. Immunol. Pract. 2014, 2, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Masaki, K.; Fukunaga, K.; Matsusaka, M.; Kabata, H.; Tanosaki, T.; Mochimaru, T.; Kamatani, T.; Ohtsuka, K.; Baba, R.; Ueda, S.; et al. Characteristics of severe asthma with fungal sensitization. Ann. Allergy Asthma Immunol. 2017, 119, 253–257. [Google Scholar] [CrossRef]
- Zureik, M.; Neukirch, C.; Leynaert, B.; Liard, R.; Bousquet, J.; Neukirch, F. Sensitisation to airborne moulds and severity of asthma: Cross sectional study from European Community respiratory health survey. BMJ 2002, 325, 411–414. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. The airway epithelium in asthma. Nat. Med. 2012, 18, 684–692. [Google Scholar] [CrossRef]
- Manuyakorn, W.; Howarth, P.H.; Holgate, S.T. Airway remodelling in asthma and novel therapy. Asian Pac. J. Allergy Immunol. 2013, 31, 3–10. [Google Scholar]
- Fehrenbach, H.; Wagner, C.; Wegmann, M. Airway remodeling in asthma: What really matters. Cell Tissue Res. 2017, 367, 551–569. [Google Scholar] [CrossRef]
- Lezmi, G.; Deschildre, A.; Taam, R.A.; Fayon, M.; Blanchon, S.; Troussier, F.; Mallinger, P.; Mahut, B.; Gosset, P.; De Blic, J. Remodelling and inflammation in preschoolers with severe recurrent wheeze and asthma outcome at school age. Clin. Exp. Allergy 2018, 48, 806–813. [Google Scholar] [CrossRef]
- Kaminska, M.; Foley, S.; Maghni, K.; Storness-Bliss, C.; Coxson, H.; Ghezzo, H.; Lemière, C.; Olivenstein, R.; Ernst, P.; Hamid, Q.; et al. Airway remodeling in subjects with severe asthma with or without chronic persistent airflow obstruction. J. Allergy Clin. Immunol. 2009, 124, 45–51.e4. [Google Scholar] [CrossRef]
- Grenier, P.A.; Fetita, C.I.; Brillet, P.Y. Quantitative computed tomography imaging of airway remodeling in severe asthma. Quant. Imaging Med. Surg. 2016, 6, 76–83. [Google Scholar] [PubMed]
- Gupta, S.; Siddiqui, S.; Haldar, P.; Entwisle, J.J.; Mawby, D.; Wardlaw, A.; Bradding, P.; Pavord, I.; Green, R.H.; Brightling, C. Quantitative analysis of high-resolution computed tomography scans in severe asthma subphenotypes. Thorax 2010, 65, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Aysola, R.S.; Hoffman, E.A.; Gierada, D.; Wenzel, S.; Cook-Granroth, J.; Tarsi, J.; Zheng, J.; Schechtman, K.B.; Ramkumar, T.P.; Cochran, R.; et al. Airway remodeling measured by multidetector CT is increased in severe asthma and correlates with pathology. Chest 2008, 134, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.I.; Colby, T.V.; Muller, N.L. Asthma and associated conditions: High-resolution CT and pathologic findings. AJR Am. J. Roentgenol. 2004, 183, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Coman, I.; Pola-Bibián, B.; Barranco, P.; Vila-Nadal, G.; Dominguez-Ortega, J.; Romero, D.; Villasante, C.; Quirce, S. Bronchiectasis in severe asthma: Clinical features and outcomes. Ann. Allergy Asthma Immunol. 2018, 120, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Castro-Rodriguez, J.A.; Saglani, S.; Rodriguez-Martinez, C.E.; Oyarzun, M.A.; Fleming, L.; Bush, A. The relationship between inflammation and remodeling in childhood asthma: A systematic review. Pediatr. Pulmonol. 2018, 53, 824–835. [Google Scholar] [CrossRef]
- Saglani, S.; Lloyd, C.M. Novel concepts in airway inflammation and remodelling in asthma. Eur. Respir. J. 2015, 46, 1796–1804. [Google Scholar] [CrossRef]
- Chaudhuri, R.; Rubin, A.; Sumino, K.; e Silva, J.R.L.; Niven, R.; Siddiqui, S.; Klooster, K.; McEvoy, C.; Shah, P.L.; Simoff, M.; et al. Safety and effectiveness of bronchial thermoplasty after 10 years in patients with persistent asthma (BT10+): A follow-up of three randomised controlled trials. Lancet Respir Med 2021, 9, 457–466. [Google Scholar] [CrossRef]
- Kosmidis, C.; Denning, D.W. The clinical spectrum of pulmonary aspergillosis. Thorax 2015, 70, 270–277. [Google Scholar] [CrossRef]
- Anees-Hill, S.; Douglas, P.; Pashley, C.H.; Hansell, A.; Marczylo, E.L. A systematic review of outdoor airborne fungal spore seasonality across Europe and the implications for health. Sci. Total Environ. 2021, 151716. [Google Scholar] [CrossRef]
- Rick, E.; Woolnough, K.F.; Seear, P.J.; Fairs, A.; Satchwell, J.; Richardson, M.; Monteiro, W.R.; Craner, M.; Bourne, M.; Wardlaw, A.J.; et al. The airway fungal microbiome in asthma. Clin. Exp. Allergy 2020, 50, 1325–1341. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.G.; Chishimba, L.; Niven, R.M.; Bromley, M.; Simpson, A.; Smyth, L.; Denning, D.; Bowyer, P. Corticosteroid treatment is associated with increased filamentous fungal burden in allergic fungal disease. J. Allergy Clin. Immunol. 2018, 142, 407–414. [Google Scholar] [CrossRef]
- Wardlaw, A.J.; Rick, E.-M.; Ozyigit, L.P.; Scadding, A.; Gaillard, E.A.; Pashley, C.H. New Perspectives in the Diagnosis and Management of Allergic Fungal Airway Disease. J. Asthma Allergy 2021, 14, 557–573. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef]
- Agarwal, R.; Aggarwal, A.N.; Gupta, D.; Jindal, S.K. Aspergillus hypersensitivity and allergic bronchopulmonary aspergillosis in patients with bronchial asthma: Systematic review and meta-analysis. Int. J. Tuberc. Lung Dis. 2009, 13, 936–944. [Google Scholar]
- O’Driscoll, B.R.; Hopkinson, L.C.; Denning, D.W. Mold sensitization is common amongst patients with severe asthma requiring multiple hospital admissions. BMC Pulm. Med. 2005, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Medrek, S.K.; Kao, C.C.; Yang, D.H.; Hanania, N.A.; Parulekar, A.D. Fungal Sensitization Is Associated with Increased Risk of Life-Threatening Asthma. J. Allergy Clin. Immunol. Pract. 2017, 5, 1025–1031.e2. [Google Scholar] [CrossRef] [PubMed]
- Black, P.N.; Udy, A.A.; Brodie, S.M. Sensitivity to fungal allergens is a risk factor for life-threatening asthma. Allergy 2000, 55, 501–504. [Google Scholar] [CrossRef]
- Denning, D.W.; Pashley, C.; Hartl, D.; Wardlaw, A.; Godet, C.; Del Giacco, S.; Delhaes, L.; Sergejeva, S. Fungal allergy in asthma-state of the art and research needs. Clin. Transl. Allergy 2014, 4, 14. [Google Scholar] [CrossRef]
- Agarwal, R.; Gupta, D. Severe asthma and fungi: Current evidence. Med. Mycol. 2011, 49 (Suppl. S1), S150–S157. [Google Scholar] [CrossRef]
- Bendien, S.A.; van Loon-Kooij, S.; Kramer, G.; Huijgen, W.; Altenburg, J.; Ten Brinke, A.; Maitland-van der Zee, A.H. Bronchiectasis in Severe Asthma: Does It Make a Difference? Respiration 2020, 99, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Menzies, D.; Holmes, L.; McCumesky, G.; Prys-Picard, C.; Niven, R. Aspergillus sensitization is associated with airflow limitation and bronchiectasis in severe asthma. Allergy 2011, 66, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Fairs, A.; Agbetile, J.; Hargadon, B.; Bourne, M.; Monteiro, W.R.; Brightling, C.E.; Bradding, P.; Green, R.H.; Mutalithas, K.; Desai, D.; et al. IgE sensitization to Aspergillus fumigatus is associated with reduced lung function in asthma. Am. J. Respir. Crit. Care Med. 2010, 182, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Máiz, L.; Vendrell, M.; Olveira, C.; Girón, R.; Nieto, R.; Martínez-García, M.Á. Prevalence and factors associated with isolation of Aspergillus and Candida from sputum in patients with non-cystic fibrosis bronchiectasis. Respiration 2015, 89, 396–403. [Google Scholar] [CrossRef]
- Sisodia, J.; Bajaj, T. Allergic Bronchopulmonary Aspergillosis. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Chetty, A. Pathology of allergic bronchopulmonary aspergillosis. Front. Biosci. 2003, 8, e110–e114. [Google Scholar] [CrossRef]
- Mitchell, T.A.M.; Hamilos, D.L.; Lynch, D.A.; Newell, J.D. Distribution and Severity of Bronchiectasis in Allergic Bronchopulmonary Aspergillosis (ABPA). J. Asthma 2000, 37, 65–72. [Google Scholar] [CrossRef]
- De Soyza, A.; Aliberti, S. Bronchiectasis and Aspergillus: How are they linked? Med. Mycol. 2017, 55, 69–81. [Google Scholar] [CrossRef]
- Angus, R.M.; Davies, M.L.; Cowan, M.D.; McSharry, C.; Thomson, N. Computed tomographic scanning of the lung in patients with allergic bronchopulmonary aspergillosis and in asthmatic patients with a positive skin test to Aspergillus fumigatus. Thorax 1994, 49, 586–589. [Google Scholar] [CrossRef][Green Version]
- Holgate, S.T.; Roberts, G.; Arshad, H.S.; Howarth, P.H.; Davies, D.E. The role of the airway epithelium and its interaction with environmental factors in asthma pathogenesis. Proc. Am. Thorac. Soc. 2009, 6, 655–659. [Google Scholar] [CrossRef]
- Sheppard, D.C. Molecular mechanism of Aspergillus fumigatus adherence to host constituents. Curr. Opin. Microbiol. 2011, 14, 375–379. [Google Scholar] [CrossRef]
- Takahashi-Nakaguchi, A.; Sakai, K.; Takahashi, H.; Hagiwara, D.; Toyotome, T.; Chibana, H.; Watanabe, A.; Yaguchi, T.; Yamaguchi, M.; Kamei, K.; et al. Aspergillus fumigatus adhesion factors in dormant conidia revealed through comparative phenotypic and transcriptomic analyses. Cell Microbiol. 2018, 20, e12802. [Google Scholar] [CrossRef]
- Liu, H.; Lee, M.J.; Solis, N.V.; Phan, Q.T.; Swidergall, M.; Ralph, B.; Ibrahim, A.S.; Sheppard, D.C.; Filler, S.G. Aspergillus fumigatus CalA binds to integrin α(5)β(1) and mediates host cell invasion. Nat. Microbiol. 2016, 2, 16211. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-Y.; Chen, F.; Sun, H.; Chen, C.; Zhao, B.-L. Important factors mediates the adhesion of Aspergillus fumigatus to alveolar epithelial cells with E-cadherin. Am. J. Transl. Res. 2016, 8, 2419–2425. [Google Scholar] [PubMed]
- Bertuzzi, M.; Hayes, G.E.; Icheoku, U.J.; Van Rhijn, N.; Denning, D.W.; Osherov, N.; Bignell, E.M. Anti-Aspergillus Activities of the Respiratory Epithelium in Health and Disease. J. Fungi 2018, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Paris, S.; Boisvieux-Ulrich, E.; Crestani, B.; Houcine, O.; Taramelli, D.; Lombardi, L.; Latgé, J.P. Internalization of Aspergillus fumigatus conidia by epithelial and endothelial cells. Infect. Immun. 1997, 65, 1510–1514. [Google Scholar] [CrossRef] [PubMed]
- Amitani, R.; Kawanami, R. Interaction of Aspergillus with human respiratory mucosa: A study with organ culture model. Med. Mycol. 2009, 47 (Suppl. S1), S127–S131. [Google Scholar] [CrossRef] [PubMed]
- Kerr, S.C.; Fischer, G.J.; Sinha, M.; McCabe, O.; Palmer, J.M.; Choera, T.; Lim, F.Y.; Wimmerova, M.; Carrington, S.D.; Yuan, S.; et al. FleA Expression in Aspergillus fumigatus Is Recognized by Fucosylated Structures on Mucins and Macrophages to Prevent Lung Infection. PLoS Pathog. 2016, 12, e1005555. [Google Scholar] [CrossRef]
- Yang, Z.; Jaeckisch, S.M.; Mitchell, C.G. Enhanced binding of Aspergillus fumigatus spores to A549 epithelial cells and extracellular matrix proteins by a component from the spore surface and inhibition by rat lung lavage fluid. Thorax 2000, 55, 579–584. [Google Scholar] [CrossRef]
- Bromley, I.M.; Donaldson, K. Binding of Aspergillus fumigatus spores to lung epithelial cells and basement membrane proteins: Relevance to the asthmatic lung. Thorax 1996, 51, 1203–1209. [Google Scholar] [CrossRef]
- Bouchara, J.P.; Sanchez, M.; Chevailler, A.; Marot-Leblond, A.; Lissitzky, J.C.; Tronchin, G.; Chabasse, D. Sialic acid-dependent recognition of laminin and fibrinogen by Aspergillus fumigatus conidia. Infect. Immun. 1997, 65, 2717–2724. [Google Scholar] [CrossRef]
- Peñalver, M.C.; O’Connor, J.E.; Martinez, J.P.; Gil, M.L. Binding of human fibronectin to Aspergillus fumigatus conidia. Infect. Immun. 1996, 64, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Coulot, P.; Bouchara, J.P.; Renier, G.; Annaix, V.; Planchenault, C.; Tronchin, G.; Chabasse, D. Specific interaction of Aspergillus fumigatus with fibrinogen and its role in cell adhesion. Infect. Immun. 1994, 62, 2169–2177. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Gautam, P.; Pandit, H.; Singh, Y.; Basir, S.F.; Madan, T. Identification of fibrinogen-binding proteins of Aspergillus fumigatus using proteomic approach. Mycopathologia 2012, 173, 73–82. [Google Scholar] [CrossRef]
- Wasylnka, J.A.; Moore, M.M. Adhesion of Aspergillus species to extracellular matrix proteins: Evidence for involvement of negatively charged carbohydrates on the conidial surface. Infect. Immun. 2000, 68, 3377–3384. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Bayry, J.; Bozza, S.; Kniemeyer, O.; Perruccio, K.; Elluru, S.R.; Clavaud, C.; Paris, S.; Brakhage, A.A.; Kaveri, S.V.; et al. Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature 2009, 460, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-K.; Lu, X.; Li, X.; Sun, Q.-Y.; Su, X.; Song, Y.; Sun, H.-M.; Shi, Y. Dectin-1 is inducible and plays a crucial role in Aspergillus-induced innate immune responses in human bronchial epithelial cells. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2755–2764. [Google Scholar] [CrossRef]
- Werner, J.L.; Metz, A.E.; Horn, D.; Schoeb, T.R.; Hewitt, M.M.; Schwiebert, L.M. Requisite role for the dectin-1 beta-glucan receptor in pulmonary defense against Aspergillus fumigatus. J. Immunol. 2009, 182, 4938–4946. [Google Scholar] [CrossRef]
- Carrion Sde, J.; Leal, S.M., Jr.; Ghannoum, M.A.; Aimanianda, V.; Latgé, J.P.; Pearlman, E. The RodA hydrophobin on Aspergillus fumigatus spores masks dectin-1- and dectin-2-dependent responses and enhances fungal survival in vivo. J. Immunol. 2013, 191, 2581–2588. [Google Scholar] [CrossRef]
- Bigot, J.; Guillot, L.; Guitard, J.; Ruffin, M.; Corvol, H.; Balloy, V.; Hennequin, C. Bronchial Epithelial Cells on the Front Line to Fight Lung Infection-Causing Aspergillus fumigatus. Fron. Immunol. 2020, 11, 1041. [Google Scholar] [CrossRef]
- Heijink, I.H.; Kuchibhotla, V.N.S.; Roffel, M.P.; Maes, T.; Knight, D.A.; Sayers, I.; Nawijn, M.C. Epithelial cell dysfunction, a major driver of asthma development. Allergy 2020, 75, 1902–1917. [Google Scholar] [CrossRef]
- Fedorov, I.A.; Wilson, S.J.; Davies, D.; Holgate, S.T. Epithelial stress and structural remodelling in childhood asthma. Thorax 2005, 60, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Xu, A.R.; Chen, S.Y.; Huang, Y.; Han, X.R.; Guan, W.J.; Wang, D.Y.; Zhong, N.S. Aberrant Epithelial Cell Proliferation in Peripheral Airways in Bronchiectasis. Front. Cell Dev. Biol. 2020, 8, 88. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, P.; Denning, D.W. Genomic analysis of allergen genes in Aspergillus spp.: The relevance of genomics to everyday research. Med. Mycol. 2007, 45, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Amitani, R.; Taylor, G.; Elezis, E.N.; Llewellyn-Jones, C.; Mitchell, J.; Kuze, F.; Cole, P.J.; Wilson, R. Purification and characterization of factors produced by Aspergillus fumigatus which affect human ciliated respiratory epithelium. Infect. Immun. 1995, 63, 3266–3271. [Google Scholar] [CrossRef] [PubMed]
- Amitani, R.; Murayama, T.; Nawada, R.; Lee, W.; Niimi, A.; Suzuki, K.; Tanaka, E.; Kuze, F. Aspergillus culture filtrates and sputum sols from patients with pulmonary aspergillosis cause damage to human respiratory ciliated epithelium in vitro. Eur. Respir. J. 1995, 8, 1681–1687. [Google Scholar] [CrossRef]
- Cody, D.T., II.; McCaffrey, T.V.; Roberts, G.; Kern, E.B. Effects of Aspergillus fumigatus and Alternaria alternata on human ciliated epithelium in vitro. Laryngoscope 1997, 107 Pt 1, 1511–1514. [Google Scholar] [CrossRef]
- Botterel, F.; Cordonnier, C.; Barbier, V.; Wingerstmann, L.; Liance, M.; Coste, A.; Escudier, E.; Bretagne, S. Aspergillus fumigatus causes in vitro electrophysiological and morphological modifications in human nasal epithelial cells. Histol. Histopathol. 2002, 17, 1095–1101. [Google Scholar]
- Khoufache, K.; Puel, O.; Loiseau, N.; Delaforge, M.; Rivollet, D.; Coste, A.; Cordonnier, C.; Escudier, E.; Botterel, F.; Bretagne, S. Verruculogen associated with Aspergillus fumigatus hyphae and conidia modifies the electrophysiological properties of human nasal epithelial cells. BMC Microbiol. 2007, 7, 5. [Google Scholar] [CrossRef]
- Crameri, R.; Limacher, A.; Weichel, M.; Glaser, A.G.; Zeller, S.; Rhyner, C. Structural aspects and clinical relevance of Aspergillus fumigatus antigens/allergens. Med. Mycol. 2006, 44 Suppl. S1, S261–S267. [Google Scholar] [CrossRef][Green Version]
- Muthu, V.; Singh, P.; Choudhary, H.; Dhooria, S.; Sehgal, I.S.; Prasad, K.T.; Aggarwal, A.N.; Garg, M.; Chakrabarti, A.; Agarwal, R. Role of recombinant Aspergillus fumigatus antigens in diagnosing Aspergillus sensitisation among asthmatics. Mycoses 2020, 63, 928–936. [Google Scholar] [CrossRef]
- Muthu, V.; Singh, P.; Choudhary, H.; Sehgal, I.S.; Dhooria, S.; Prasad, K.T.; Aggarwal, A.N.; Garg, M.; Chakrabarti, A.; Agarwal, R. Diagnostic Cutoffs and Clinical Utility of Recombinant Aspergillus fumigatus Antigens in the Diagnosis of Allergic Bronchopulmonary Aspergillosis. J. Allergy Clin. Immunol. Pract. 2020, 8, 579–587. [Google Scholar] [CrossRef]
- Kespohl, S.; Raulf, M. Mould allergens: Where do we stand with molecular allergy diagnostics?: Part 13 of the series Molecular Allergology. Allergo J. Int. 2014, 23, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.W.; Venaille, T.J.; Mendis, A.H.; McAleer, R. Allergens as proteases: An Aspergillus fumigatus proteinase directly induces human epithelial cell detachment. J. Allergy Clin. Immunol. 1990, 86, 726–731. [Google Scholar] [CrossRef]
- Sharon, H.; Amar, D.; Levdansky, E.; Mircus, G.; Shadkchan, Y.; Shamir, R.; Osherov, N. PrtT-regulated proteins secreted by Aspergillus fumigatus activate MAPK signaling in exposed A549 lung cells leading to necrotic cell death. PLoS ONE 2011, 6, e17509. [Google Scholar] [CrossRef]
- Kogan, T.V.; Jadoun, J.; Mittelman, L.; Hirschberg, K.; Osherov, N. Involvement of secreted Aspergillus fumigatus proteases in disruption of the actin fiber cytoskeleton and loss of focal adhesion sites in infected A549 lung pneumocytes. J. Infect. Dis. 2004, 189, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Daly, P.; Verhaegen, S.; Clynes, M.; Kavanagh, K. Culture filtrates of Aspergillus fumigatus induce different modes of cell death in human cancer cell lines. Mycopathologia 1999, 146, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.L.; Merkhofer, R.; Ober, C.; Kujoth, G.C.; Niu, M.; Keller, N.P.; Gern, J.E.; Brockman-Schneider, R.A.; Evans, M.; Jackson, D.J.; et al. Cell TRPV4 Serves as a Damage Sensor Driving Lung Allergic Inflammation. Cell Host Microbe 2020, 27, 614–628.e6. [Google Scholar] [CrossRef]
- Farnell, E.; Rousseau, K.; Thornton, D.J.; Bowyer, P.; Herrick, S.E. Expression and secretion of Aspergillus fumigatus proteases are regulated in response to different protein substrates. Fungal Biol. 2012, 116, 1003–1012. [Google Scholar] [CrossRef]
- Iadarola, P.; Lungarella, G.; Martorana, P.A.; Viglio, S.; Guglielminetti, M.; Korzus, E.; Gorrini, M.; Cavarra, E.; Rossi, A.; Travis, J.; et al. Lung injury and degradation of extracellular matrix components by Aspergillus fumigatus serine proteinase. Exp. Lung Res. 1998, 24, 233–251. [Google Scholar] [CrossRef]
- Larcher, G.; Bouchara, J.P.; Annaix, V.; Symoens, F.; Chabasse, D.; Tronchin, G. Purification and characterization of a fibrinogenolytic serine proteinase from Aspergillus fumigatus culture filtrate. FEBS Lett. 1992, 308, 65–69. [Google Scholar] [CrossRef]
- Gifford, A.H.; Klippenstein, J.R.; Moore, M.M. Serum stimulates growth of and proteinase secretion by Aspergillus fumigatus. Infect. Immun. 2002, 70, 19–26. [Google Scholar] [CrossRef]
- Kauffman, H.F.; Tomee, J.; van de Riet, M.A.; Timmerman, A.J.; Borger, P. Protease-dependent activation of epithelial cells by fungal allergens leads to morphologic changes and cytokine production. J. Allergy Clin. Immunol. 2000, 105 Pt 1, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Tomee, J.F.C.; Wierenga, A.T.J.; Hiemstra, P.; Kauffman, H.F. Proteases from Aspergillus fumigatus induce release of proinflammatory cytokines and cell detachment in airway epithelial cell lines. J. Infect. Dis. 1997, 176, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.; Namvar, S.; Gago, S.; Labram, B.; Bowyer, P.; Richardson, M.; Herrick, S. Differential Proinflammatory Responses to Aspergillus fumigatus by Airway Epithelial Cells In Vitro Are Protease Dependent. J. Fungi 2021, 7, 468. [Google Scholar] [CrossRef] [PubMed]
- Kurup, V.P.; Grunig, G. Animal models of allergic bronchopulmonary aspergillosis. Mycopathologia 2002, 153, 165–177. [Google Scholar] [CrossRef]
- Desoubeaux, G.; Cray, C. Animal Models of Aspergillosis. Comp. Med. 2018, 68, 109–123. [Google Scholar]
- Buskirk, A.D.; Green, B.J.; Lemons, A.; Nayak, A.P.; Goldsmith, W.T.; Kashon, M.L.; Anderson, S.E.; Hettick, J.M.; Templeton, S.P.; Germolec, D.R.; et al. A murine inhalation model to characterize pulmonary exposure to dry Aspergillus fumigatus conidia. PLoS ONE 2014, 9, e109855. [Google Scholar] [CrossRef]
- Urb, M.; Snarr, B.D.; Wojewodka, G.; Lehoux, M.; Lee, M.J.; Ralph, B.; Divangahi, M.; King, I.L.; McGovern, T.K.; Martin, J.G.; et al. Evolution of the Immune Response to Chronic Airway Colonization with Aspergillus fumigatus Hyphae. Infect. Immun. 2015, 83, 3590–3600. [Google Scholar] [CrossRef]
- Porter, P.C.; Roberts, L.; Fields, A.; Knight, M.; Qian, Y.; Delclos, G.L.; Han, S.; Kheradmand, F.; Corry, D.B. Necessary and sufficient role for T helper cells to prevent fungal dissemination in allergic lung disease. Infect. Immun. 2011, 79, 4459–4471. [Google Scholar] [CrossRef]
- Bertuzzi, M.; Van Rhijn, N.; Krappmann, S.; Bowyer, P.; Bromley, M.; Bignell, E. On the lineage of Aspergillus fumigatus isolates in common laboratory use. Med. Mycol. 2020, 59, 7–13. [Google Scholar] [CrossRef]
- Jones, J.T.; Liu, K.W.; Wang, X.; Kowalski, C.H.; Ross, B.S.; Mills, K.A.M.; Kerkaert, J.D.; Hohl, T.M.; Lofgren, L.A.; Stajich, J.E. Aspergillus fumigatus Strain-Specific Conidia Lung Persistence Causes an Allergic Broncho-Pulmonary Aspergillosis-Like Disease Phenotype. mSphere 2021, 6, e01250-20. [Google Scholar] [CrossRef] [PubMed]
- Hogaboam, C.M.; Blease, K.; Mehrad, B.; Steinhauser, M.L.; Standiford, T.J.; Kunkel, S.L.; Lukacs, N.W. Chronic airway hyperreactivity, goblet cell hyperplasia, and peribronchial fibrosis during allergic airway disease induced by Aspergillus fumigatus. Am. J. Pathol. 2000, 156, 723–732. [Google Scholar] [CrossRef]
- Hoselton, S.A.; Samarasinghe, A.E.; Seydel, J.M.; Schuh, J.M. An inhalation model of airway allergic response to inhalation of environmental Aspergillus fumigatus conidia in sensitized BALB/c mice. Med. Mycol. 2010, 48, 1056–1065. [Google Scholar] [CrossRef]
- Ghosh, S.; Hoselton, S.A.; Asbach, S.V.; Steffan, B.N.; Wanjara, S.B.; Dorsam, G.P.; Schuh, J.M. B lymphocytes regulate airway granulocytic inflammation and cytokine production in a murine model of fungal allergic asthma. Cell Mol. Immunol. 2015, 12, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Kurup, V.P.; Guo, J.; Murali, P.S.; Choi, H.; Fink, J.N. Immunopathologic responses to Aspergillus antigen in interleukin-4 knockout mice. J. Lab Clin. Med. 1997, 130, 567–575. [Google Scholar] [CrossRef]
- Blease, K.; Jakubzick, C.; Westwick, J.; Lukacs, N.; Kunkel, S.L.; Hogaboam, C.M. Therapeutic effect of IL-13 immunoneutralization during chronic experimental fungal asthma. J. Immunol. 2001, 166, 5219–5224. [Google Scholar] [CrossRef]
- Labram, B.; Namvar, S.; Hussell, T.; Herrick, S.E. Endothelin-1 mediates Aspergillus fumigatus-induced airway inflammation and remodelling. Clin. Exp. Allergy 2019, 49, 861–873. [Google Scholar] [CrossRef]
- Balenga, N.A.; Klichinsky, M.; Xie, Z.; Chan, E.C.; Zhao, M.; Jude, J.; Laviolette, M.; Panettieri, R.A., Jr.; Druey, K.M. A fungal protease allergen provokes airway hyper-responsiveness in asthma. Nat. Commun. 2015, 6, 6763. [Google Scholar] [CrossRef]
- Redes, J.L.; Basu, T.; Ram-Mohan, S.; Ghosh, C.C.; Chan, E.C.; Sek, A.; Zhao, M.; Krishnan, R.; Rosenberg, H.F.; Druey, K.M. Aspergillus fumigatus-Secreted Alkaline Protease 1 Mediates Airways Hyperresponsiveness in Severe Asthma. Immunohorizons 2019, 3, 368–377. [Google Scholar]
- Namvar, S.; Warn, P.; Farnell, E.; Bromley, M.; Fraczek, M.; Bowyer, P.; Herrick, S. Aspergillus fumigatus proteases, Asp f 5 and Asp f 13, are essential for airway inflammation and remodelling in a murine inhalation model. Clin. Exp. Allergy 2015, 45, 982–993. [Google Scholar] [CrossRef]
- Oguma, T.; Asano, K.; Tomomatsu, K.; Kodama, M.; Fukunaga, K.; Shiomi, T.; Ohmori, N.; Ueda, S.; Takihara, T.; Shiraishi, Y. Induction of mucin and MUC5AC expression by the protease activity of Aspergillus fumigatus in airway epithelial cells. J. Immunol. 2011, 187, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Basu, T.; Seyedmousavi, S.; Sugui, J.A.; Balenga, N.; Zhao, M.; Kwon Chung, K.J.; Biardel, S.; Laviolette, M.; Druey, K.M. Aspergillus fumigatus alkaline protease 1 (Alp1/Asp f13) in the airways correlates with asthma severity. J. Allergy Clin. Immunol. 2018, 141, 423–425.e7. [Google Scholar] [CrossRef] [PubMed]
- Ossovskaya, V.S.; Bunnett, N.W. Protease-activated receptors: Contribution to physiology and disease. Physiol. Rev. 2004, 84, 579–621. [Google Scholar] [CrossRef] [PubMed]
- Yike, I. Fungal proteases and their pathophysiological effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.E.; Kita, H. The role of protease activation of inflammation in allergic respiratory diseases. J. Allergy Clin. Immunol. 2004, 114, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Skorkowska, K.; Adamiec, R. The biological role of protease-activated receptors in angiology. The present view. Int. Angiol. 2005, 24, 215–220. [Google Scholar]
- Ebeling, C.; Forsythe, P.; Ng, J.; Gordon, J.R.; Hollenberg, M.; Vliagoftis, H. Proteinase-activated receptor 2 activation in the airways enhances antigen-mediated airway inflammation and airway hyperresponsiveness through different pathways. J. Allergy Clin. Immunol. 2005, 115, 623–630. [Google Scholar] [CrossRef]
- Knight, D.A.; Lim, S.; Scaffidi, A.K.; Roche, N.; Chung, K.F.; Stewart, G.A.; Thompson, P.J. Protease-activated receptors in human airways: Upregulation of PAR-2 in respiratory epithelium from patients with asthma. J. Allergy Clin. Immunol. 2001, 108, 797–803. [Google Scholar] [CrossRef]
- Moretti, S.; Bellocchio, S.; Bonifazi, P.; Bozza, S.; Zelante, T.; Bistoni, F.; Romani, L. The contribution of PARs to inflammation and immunity to fungi. Mucosal Immunol. 2008, 1, 156–168. [Google Scholar] [CrossRef]
- Wu, X.; Lee, B.; Zhu, L.; Ding, Z.; Chen, Y. Exposure to mold proteases stimulates mucin production in airway epithelial cells through Ras/Raf1/ERK signal pathway. PLoS ONE 2020, 15, e0231990. [Google Scholar] [CrossRef]
- Holgate, S.T.; Holloway, J.; Wilson, S.; Bucchieri, F.; Puddicombe, S.; Davies, D.E. Epithelial-mesenchymal communication in the pathogenesis of chronic asthma. Proc. Am. Thorac. Soc. 2004, 1, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Smartt, H.; Holgate, S.T.; Roche, W.R. Growth factors secreted by bronchial epithelial cells control myofibroblast proliferation: An in vitro co-culture model of airway remodeling in asthma. Lab Investig. 1999, 79, 395–405. [Google Scholar] [PubMed]
- Ojiaku, C.A.; Yoo, E.J.; Panettieri, R.A., Jr. Transforming Growth Factor β1 Function in Airway Remodeling and Hyperresponsiveness. The Missing Link? Am. J. Respir. Cell Mol. Biol. 2017, 56, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Skrgat, S.; Malovrh, M.M.; Sarc, I.; Šilar, M.; Dimitric, V.; Korosec, P. TSLP as biomarker in asthma patients. Eur. Respir. J. 2015, 46 (Suppl. S59), PA3868. [Google Scholar]
- Burgess, J.K.; Jonker, M.R.; Berg, M.; Ten Hacken, N.T.H.; Meyer, K.B.; van den Berge, M.; Nawijn, M.C.; Heijink, I.H. Periostin: Contributor to abnormal airway epithelial function in asthma? Eur. Respir. J. 2021, 57, 2001286. [Google Scholar] [CrossRef]
- Cheng, D.; Xue, Z.; Yi, L.; Shi, H.; Zhang, K.; Huo, X.; Bonser, L.R.; Zhao, J.; Xu, Y.; Erle, D.; et al. Epithelial interleukin-25 is a key mediator in Th2-high, corticosteroid-responsive asthma. Am. J. Respir. Crit. Care Med. 2014, 190, 639–648. [Google Scholar] [CrossRef]
- Drake, L.Y.; Kita, H. IL-33: Biological properties, functions, and roles in airway disease. Immunol. Rev. 2017, 278, 173–184. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Lv, Z.; Li, Y.; Chen, Y.; Huang, K.; Corrigan, C.J.; Ying, S. Elevated Expression of IL-33 and TSLP in the Airways of Human Asthmatics In Vivo: A Potential Biomarker of Severe Refractory Disease. J. Immunol. 2018, 200, 2253. [Google Scholar] [CrossRef]
- Khosravi, A.R.; Shokri, H.; Hassan Al-Heidary, S.; Ghafarifar, F. Evaluation of murine lung epithelial cells (TC-1 JHU-1) line to develop Th2-promoting cytokines IL-25/IL-33/TSLP and genes Tlr2/Tlr4 in response to Aspergillus fumigatus. J. Mycol. Med. 2018, 28, 349–354. [Google Scholar] [CrossRef]
- Verma, M.; Liu, S.; Michalec, L.; Sripada, A.; Gorska, M.M.; Alam, R. Experimental asthma persists in IL-33 receptor knockout mice because of the emergence of thymic stromal lymphopoietin-driven IL-9(+) and IL-13(+) type 2 innate lymphoid cell subpopulations. J. Allergy Clin. Immunol. 2018, 142, 793–803.e8. [Google Scholar] [CrossRef]
- Gordon, E.D.; Sidhu, S.S.; Wang, Z.E.; Woodruff, P.G.; Yuan, S.; Solon, M.C.; Conway, S.J.; Huang, X.; Locksley, R.M.; Fahy, J.V. A protective role for periostin and TGF-beta in IgE-mediated allergy and airway hyperresponsiveness. Clin. Exp. Allergy 2012, 42, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Khor, Y.H.; Teoh, A.K.Y.; Lam, S.M.; Mo, D.C.Q.; Weston, S.; Reid, D.W.; Walters, E.H. Increased vascular permeability precedes cellular inflammation as asthma control deteriorates. Clin. Exp. Allergy 2009, 39, 1659–1667. [Google Scholar] [CrossRef]
- Hoshino, M.; Nakamura, Y.; Hamid, Q.A. Gene expression of vascular endothelial growth factor and its receptors and angiogenesis in bronchial asthma. J. Allergy Clin. Immunol. 2001, 107, 1034–1038. [Google Scholar] [CrossRef]
- Filep, J.G.; Sirois, M.G.; Rousseau, A.; Fournier, A.; Sirois, P. Effects of endothelin-1 on vascular permeability in the conscious rat: Interactions with platelet-activating factor. Br. J. Pharmacol. 1991, 104, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Redington, A.E.; Springall, D.R.; Ghatei, M.A.; Lau, L.C.; Bloom, S.R.; Holgate, S.T.; Polak, J.M.; Howarth, P.H. Endothelin in bronchoalveolar lavage fluid and its relation to airflow obstruction in asthma. Am. J. Respir. Crit. Care Med. 1995, 151, 1034–1039. [Google Scholar] [PubMed]
- Brims, F.J.H.; Chauhan, A.J.; Higgins, B.; Shute, J.K. Coagulation factors in the airways in moderate and severe asthma and the effect of inhaled steroids. Thorax 2009, 64, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Wagers, S.S.; Norton, R.J.; Rinaldi, L.M.; Bates, J.H.; Sobel, B.E.; Irvin, C.G. Extravascular fibrin, plasminogen activator, plasminogen activator inhibitors, and airway hyperresponsiveness. J. Clin. Investig. 2004, 114, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Panchabhai, T.S.; Mukhopadhyay, S.; Sehgal, S.; Bandyopadhyay, D.; Erzurum, S.C.; Mehta, A.C. Plugs of the Air Passages: A Clinicopathologic Review. Chest 2016, 150, 1141–1157. [Google Scholar] [CrossRef]
- de Giorgio-Miller, A.; Bottoms, S.; Laurent, G.; Carmeliet, P.; Herrick, S. Fibrin-induced skin fibrosis in mice deficient in tissue plasminogen activator. Am. J. Pathol. 2005, 167, 721–732. [Google Scholar] [CrossRef]
- Shi-Wen, X.; Chen, Y.; Denton, C.P.; Eastwood, M.; Renzoni, E.; Bou-Gharios, G.; Pearson, J.D.; Dashwood, M.; Du Bois, R.M.; Black, C.M.; et al. Endothelin-1 promotes myofibroblast induction through the ETA receptor via a rac/phosphoinositide 3-kinase/Akt-dependent pathway and is essential for the enhanced contractile phenotype of fibrotic fibroblasts. Mol. Biol. Cell 2004, 15, 2707–2719. [Google Scholar] [CrossRef]
- Millien, V.O.; Lu, W.; Shaw, J.; Yuan, X.; Mak, G.; Roberts, L.; Song, L.Z.; Knight, J.M.; Creighton, C.J.; Luong, A.; et al. Cleavage of fibrinogen by proteinases elicits allergic responses through Toll-like receptor 4. Science 2013, 341, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Dasari, P.; Shopova, I.; Stroe, M.; Wartenberg, D.; Martin-Dahse, H.; Beyersdorf, N.; Hortschansky, P.; Dietrich, S.; Cseresnyés, Z.; Figge, M.T.; et al. Aspf2 From Aspergillus fumigatus Recruits Human Immune Regulators for Immune Evasion and Cell Damage. Front. Immunol. 2018, 9, 1635. [Google Scholar] [CrossRef]
- Salo, P.M.; Arbes, S.J., Jr.; Sever, M.; Jaramillo, R.; Cohn, R.D.; London, S.J.; Zeldin, D.C. Exposure to Alternaria alternata in US homes is associated with asthma symptoms. J. Allergy Clin. Immunol. 2006, 118, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ramirez, G.; Barber, D.; Tome-Amat, J.; Garrido-Arandia, M.; Diaz-Perales, A. Alternaria as an Inducer of Allergic Sensitization. J. Fungi 2021, 7, 838. [Google Scholar] [CrossRef] [PubMed]
- Pashley, C.H.; Wardlaw, A.J. Allergic fungal airways disease (AFAD): An under-recognised asthma endotype. Mycopathologia 2021, 186, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Celakovska, J.; Vankova, R.; Bukac, J.; Cermakova, E.; Andrys, C.; Krejsek, J. Atopic Dermatitis and Sensitisation to Molecular Components of Alternaria, Cladosporium, Penicillium, Aspergillus, and Malassezia-Results of Allergy Explorer ALEX 2. J. Fungi 2021, 7, 183. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).