
Role of RIM101 for Sporulation at Alkaline pH in Ashbya gossypii


1
Carlsberg Research Laboratory, Carlsberg A/S, 1799 Copenhagen V, Denmark
2
Department of Microbiology and Biochemistry, Hochschule Geisenheim University, Von-Lade-Strasse 1, 65366 Geisenheim, Germany
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(7), 527; https://doi.org/10.3390/jof7070527
Submission received: 14 June 2021
/
Revised: 27 June 2021
/
Accepted: 28 June 2021
/
Published: 30 June 2021
(This article belongs to the Special Issue Formation and Function of Fungal Ascospores)
Abstract
:Microorganisms need to sense and adapt to fluctuations in the environmental pH. In fungal species, this response is mediated by the conserved pacC/RIM101 pathway. In Aspergillus nidulans, PacC activates alkaline-expressed genes and represses acid-controlled genes in response to alkaline pH and has important functions in regulating growth and conidia formation. In Saccharomyces cerevisiae, the PacC homolog Rim101 is required for adaptation to extracellular pH and to regulate transcription of IME1, the Initiator of MEiosis. S. cerevisiae rim101 mutants are defective in sporulation. In Ashbya gossypii, a filamentous fungus belonging to the family of Saccharomycetaceae, little is known about the role of pH in regulating growth and sporulation. Here, we deleted the AgRIM101 homolog (AFR190C). Our analyses show that Rim101 is important for growth and essential for sporulation at alkaline pH in A. gossypii. Acidic liquid sporulation media were alkalinized by sporulating strains, while the high pH of alkaline media (starting pH = 8.6) was reduced to a pH ~ 7.5 by these strains. However, Agrim101 mutants were unable to sporulate in alkaline media and failed to reduce the initial high pH, while they were capable of sporulation in acidic liquid media in which they increased the pH like the wild type.
1. Introduction
All microorganisms need to respond to environmental changes in order to survive. This often involves signal transduction pathways that transcriptionally induce genes to improve the species’ adaptation. In many fungal species, the response to changes in extracellular pH is mediated by the conserved pacC/RIM101 pathway [1].
Gene regulation by ambient pH has been most extensively studied in Aspergillus nidulans, where it is mediated by the C2H2 zinc finger transcription factor PacC. In response to alkaline pH, PacC acts both as an activator and a repressor by binding the consensus sequence GCCARG. Alkaline-expressed genes are activated while acid-controlled genes are repressed [2]. PacC plays an essential role in pulmonary pathogenesis in A. nidulans [3] and deletion results in poor growth and conidiation [4]. Orthologs of pacC are also important for virulence in other fungal species.
In the human pathogen Candida albicans, Rim101 is essential for dimorphism and virulence since it regulates alkaline pH-induced filamentation [5,6]. In the plant pathogen Fusarium oxysporum, PacC most likely acts as a negative regulator of virulence by inhibiting transcription of acid-expressed genes that are essential for pathogenicity [7]. In Magnaporthe oryzae deletion of PacC abolishes conidiation at pH 8 but only reduces virulence [8].
In Saccharomyces cerevisiae, the PacC homolog ScRim101 is a transcriptional repressor that is required for adaptation to extracellular alkalization by regulating expression of genes encoding ions pumps and transporters [9]. In contrast to PacC in A. nidulans, which acts directly as a transcriptional activator, Rim101 in S. cerevisiae activates genes indirectly via repressing transcriptional repressors, mainly NRG1 and SMP1. ScRim101-mediated repression of SMP1 transcription results in activation of IME1 expression [10]. A prerequisite for IME1 activation by ScRim101 is the absence of glucose and presence of non-fermentable sugars, which, when metabolized, leads to an alkalization of the media. This triggers the highly conserved alkaline-sensing pathway to activate Rim101 by proteolytic cleavage catalyzed by the Rim13 protease (Figure 1) [11,12,13]. In A. nidulans, this proteolytic cleavage is mediated by the signaling protease PalB [12]. Deletion of ScRIM101 results in a strain severely reduced in sporulation that fails to induce haploid-invasive growth in response to alkaline pH. A Scrim101 mutant grows poorly at alkaline pH and low temperature and is sensitive to Na+ and Li+-ions [9].
The filamentous ascomycete Ashbya gossypii is a pre-whole genome duplication fungus that belongs to the family of the yeasts Saccharomycetaceae and shares homologs of 95% of its genes with S. cerevisiae [15]. Previous studies have shown that the main regulators of sporulation in S. cerevisiae are also essential for sporulation in A. gossypii, including IME1, IME2, IME4, KAR4, NDT80, and also the cAMP/Protein Kinase A pathway [16,17,18]. In S. cerevisiae, IME1 expression requires ScRim101, and sporulation is therefore dependent on alkaline pH [19]. In this study, we investigated the role of pH on sporulation and vegetative growth in A. gossypii by deleting the AgRIM101 homolog. Our results show that AgRIM101 is important for both sporulation and growth only at alkaline pH in A. gossypii. However, RIM101-mediated signaling at alkaline pH is not a prerequisite for sporulation in A. gossypii, since both the wild-type and the rim101 mutant strains were able to sporulate abundantly when incubated at acidic starting pH (pH 5.3), which increased to ~ pH 7.2 due to metabolic activity.
2. Materials and Methods
2.1. Strains and Media
A. gossypii strains were grown in Ashbya Full Medium (AFM) (1% yeast extract, 1% peptone, 2% dextrose), and G418/geneticin (200 μg/mL) was used for selection of antibiotic-resistant transformants. For pH studies, full media plates were buffered with Tris-HCl to pH 6.5, 7.0, 7.5, 8.0, and 8.5. In sporulation assays, full media plates supplemented with 1 g/L myo-inositol were used. Liquid sporulation assays were carried out in a minimal medium (1.7 g/L yeast nitrogen base (YNB), w/o ammonium sulfate and w/o amino acids, 0.69 g/L CSM (Formedium, Hunstanton, UK), 20 g/L glucose, 2 g/L asparagine, and 1 g/L myo-inositol) buffered with Tris-HCl to pH 8.5 where indicated. Two independent transformants were generated for the Agrim101 deletion strain. The strains used in this study are listed in Table 1.
2.2. Transformation of A. gossypii
A. gossypii was transformed by electroporation as described previously [20]. G418/geneticin (200 μg/mL) was used for selection of antibiotic-resistant transformants. The A. gossypii rim101 deletion strains were generated by amplifying PCR cassettes from pFA-GEN3 using gene-specific S1- and S2-primers. These PCR cassettes were then used as transforming DNA [21]. All oligonucleotide primers were purchased from LGC genomics GmbH (Germany) and are listed in Table 2. To verify the correct integration of the cassettes and absence of the target gene in two independent homokaryotic deletion strains, diagnostic PCR was used.
2.3. Sporulation Assay
The sporulation ability at different pH values was determined in the wild-type strain and the Agrim101 mutant by isolating spores from the central part of colonies grown for 10 days at 30 °C on full media plates buffered with Tris-HCl to pH 6.5, 7.0, 7.5, 8.0, and 8.5. A circle of the central mycelia (15 mm in diameter) was cut out from the plate and suspended in a 5 mL TE buffer containing 200 µL zymolyase (10 mg/mL) to degrade vegetative hyphal cells and release the spores. After 3 hours of incubation at 37 °C (on a tilting rotor) the spores were collected by centrifugation and washed twice in spore buffer (0.03 % Triton-X-100). Serial dilutions were performed in spore buffer and 100 µL of the appropriate dilutions were plated on full media plates and incubated at 30 °C until colonies appeared. It should be noted that single-spore suspensions are difficult to obtain with Ashbya due to the hydrophobicity of the spores and a terminal filament at the proximal end of each spore, which leads to the agglomeration of spores. The CFUs therefore derive mostly from clumps of spores. The data are, thus, presented as the number of colony forming units/mL.
Sporulation in liquid culture was monitored by transferring mycelia pre-cultured in AFM to a minimal medium. These cultures were then incubated for one week at 28 °C. Thereafter, sporulation was observed by microscopy and end-point pH was measured. Experiments were done three times and representative data are shown. Clumps of spores were easily identified by microscopy. Under these conditions Ashbya mycelia sporulate quantitatively. Alkaline pH was obtained by buffering minimal media (pH 5.3) with Tris-HCl to pH 8.6.
2.4. Microscopy
Microscopic images of sporulating cultures were observed using a fully motorized Zeiss Axiovert 200M microscope equipped with a Plan-Neofluar 100×/1.30 objective (Carl Zeiss AG, Feldbach, CH). Imaging was controlled by AxioVision 4 software and single-plane brightfield images were acquired using an AxioCam MRm Rev.3 and compiled using Fiji/ImageJ 2.1.0/1.53c.
3. Results
3.1. Rim101 Has No Role in Vegetative Growth in A. gossypii
Ashbya RIM101 has been identified in the Ashbya Genome Database and annotated as AFR190C being a syntenic homolog of the S. cerevisiae YHL027W [22]. The conserved C2H2 zinc finger domains are shown in Figure 2. Similarly, RIM13/YMR154C and RIM20/YOR275C are also highly conserved in Ashbya as AgRIM13/ADR274C and AgRIM20/AER342C.
Using the well-established and highly efficient PCR-based gene-targeting approach, we generated independent rim101 deletion strains in A. gossypii that were verified by standard diagnostic PCR [20,21]. Gene deletions generate heterokaryotic mutants that harbor both wild-type and transformed nuclei in their hyphal compartments. Usually, and also in our study, heterokaryotic transformants harbor wild-type phenotypes. Sporulation of these heterokaryotic strains allows for the selection of uninucleate mutant spores based on G418 antibiotic selection, which germinate and generate homokaryotic mycelia harboring only nuclei with the linked mutation. Two independent mutant strains were generated that showed similar phenotypes. Both mutants were analyzed in parallel and representative images of one strain (AWL63) are shown unless otherwise indicated (see Table 1).
The effects on vegetative growth in the Agrim101 mutant were studied comparatively by plating strains on full medium plates and monitoring radial growth for 7 days at various temperatures (22 °C, 30 °C, and 37 °C). Under these conditions, deletion of RIM101 in A. gossypii did not reveal a negative effect on vegetative growth compared to the wild type (Figure 3).
3.2. Sporulation at High pH in A. gossypii Requires Rim101
Next, we went on to investigate the role of AgRIM101 for growth under alkaline pH conditions. We monitored radial growth of the wild-type and the Agrim101 mutant on full medium plates set to pH 6.5, 7.0, 7.5, 8.0, and 8.5. This indicated reduced vegetative growth of the Agrim101 mutant at the alkaline pH 8.0–8.5 (Figure 4).
Sporulation in S. cerevisiae is only initiated at alkaline pH when Rim101 is functional and promotes IME1 expression via repression of SMP1. Deletion of RIM101 in yeast, therefore, results in a generally sporulation-deficient strain [23]. Thus, we studied the effect of pH on sporulation in the A. gossypii wild-type and Agrim101 mutant strains. Sporulation commences from central parts of Ashbya mycelia and leads to sporangium formation of cell compartments and mycelial fragmentation. We isolated spores from these central parts of mycelia of strains grown for 10 days on full media plates buffered to different pH levels (as shown in Figure 4A). Dilutions of the spore suspensions were plated and the numbers of colony-forming units were determined (Figure 4B). This showed that AgRIM101 is required for sporulation at alkaline pH since sporulation efficiency dropped several orders of magnitude in the Agrim101 mutant at pH 8.0 and 8.5. However, both the Ashbya wild-type strain and the Agrim101 strain sporulated equally well at pH 6.5. This indicates a differential response to pH in A. gossypii resulting in the inability to sporulate when challenged by high pH. Both the Ashbya wild-type strain and the Agrim101 mutant formed similar-sized sporulation zones at pH 6.5. However, while the sporulation zone in the wild-type strain increased at pH 7.0 and 7.5, this zone got smaller and smaller in the rim101 mutant challenged by higher initial pH values.
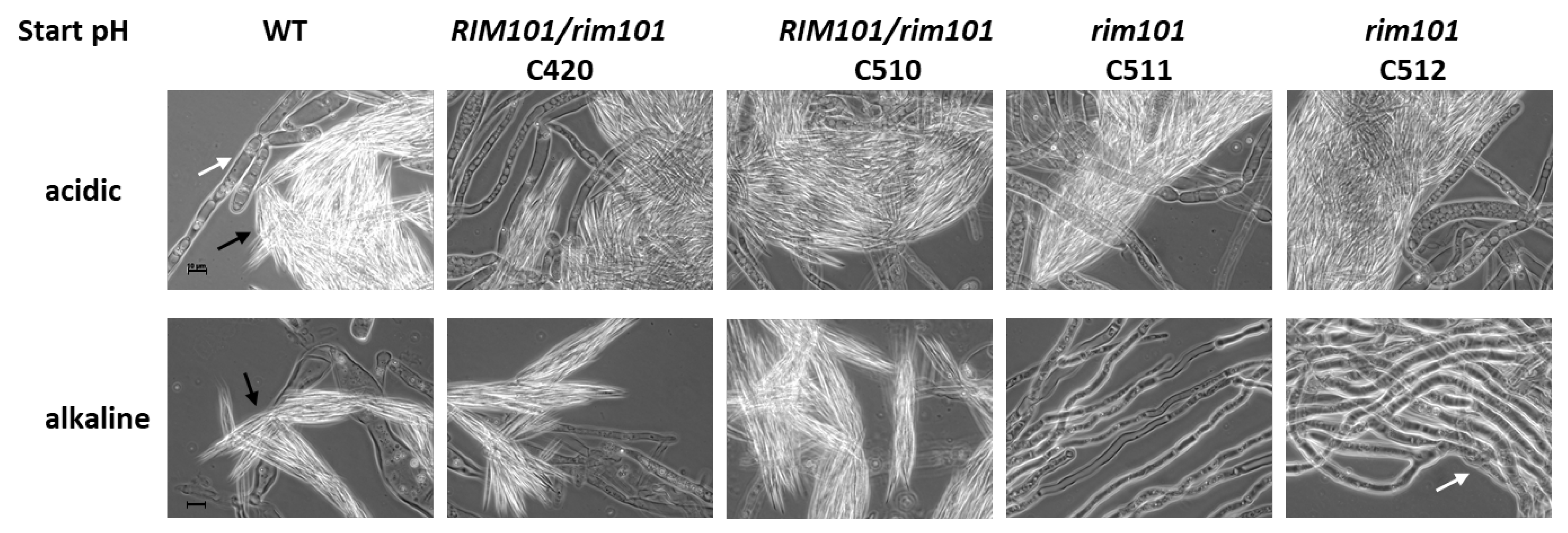
Radial colony growth at alkaline pH was reduced in the rim101 mutant compared to the wild type. Accordingly, the sporulation zone may have been decreased promoting the finding of lack of sporulation under these conditions. To analyze this in more detail we investigated sporulation potential at different pH values in liquid media. A. gossypii is able to sporulate abundantly in liquid media. Thus, the wild-type, heterokaryotic, and homokaryotic rim101 strains where pre-grown overnight in AFM and equal amounts of mycelia were transferred to the minimal media of acidic (pH 5.3) and Tris-buffered alkaline (pH 8.6) starting pH values. After one week of incubation, the cultures were analyzed. The wild-type and heterokaryotic mutants sporulated profusely under both conditions (Figure 5).
Sporulation of rim101 mutants, however, was specifically blocked in the alkaline medium, while they sporulated well in the acidic medium. Sporulation of rim101 mycelia was as abundant as in the wild-type strain under acidic starting conditions, while it was completely abolished in liquid cultures with an alkaline starting pH, where no spores were found at all. This indicates that an alkaline response via RIM101 is required to enable sporulation in Ashbya at elevated pH.
Cellular metabolism and adaptation to growth media results in alterations in media pH. Therefore, we determined the endpoint pH of these cultures. The pH of the acidic minimal media was raised to between pH 6.7 and pH 7.6 (WT pH 7.6; heterokaroytic strains C420 pH 6.7 and C510 pH 7.1; homokaryotic strains C511 pH 7.2 and C512 pH 7.0). The pH of the alkaline-buffered media was lowered in the sporulating strains of the wild type to pH 7.8 and in the heterokaryotic strains to pH 7.0 (C420) and pH 7.2 (C510). The homokaryotic rim101 mutant strains, however, failed to substantially reduce this alkaline pH, which remained high at pH 8.4 for both strains. Both homokaryotic rim101 strains failed to sporulate under these alkaline pH conditions (Figure 5).
4. Discussion
Fungi are able to grow over a wide range of extracellular pH and must therefore be able to sense and respond to changes in environmental pH. In fungi, this is mediated by the conserved Rim101/PacC pathway. In both S. cerevisiae and A. nidulans Rim101 and PacC are important developmental regulators as they promote sporulation and conidiation in these fungi, respectively. In alkaline media, A. nidulans PacC activates expression of alkaline genes and represses acidic-expressed genes, while this does not occur in acidic conditions [1 and references therein]. This is different in S. cerevisiae, where Rim101 acts indirectly by repressing the regulators NRG1 and SMP1 [10].
In our study, we showed that in Ashbya, the Rim101 homolog has a role in in adaptation to alkaline pH and sporulation under elevated pH conditions.
Our functional analysis of AgRIM101 shows that there are several differences to ScRIM101. AgRIM101 is not required for vegetative growth at 22 °C, 30 °C, and 37 °C, which is in contrast to S. cerevisiae, as Scrim101 mutants grow poorly at low temperatures [24]. Previously, it was shown in S. cerevisiae that Rim101 is required for a proper response to salt stress [25]. When the A. gossypii rim101 mutant was subjected to NaCl salt stress only a slightly increased sensitivity to 0.4 M NaCl was observed, resulting in uneven radial growth compared to the wild type.
Loss of AgRim101, however, was found to reduce radial growth at pH 7.5–8.5, and we also observed increased colony lysis of older mycelia. This phenotype is somewhat similar to that observed in the Agmkk1 and Agbck1 cell wall integrity mutants [26]. This suggests a link in Ashbya between the RIM and PKC pathways in cell wall maintenance as was shown for S. cerevisiae, where RIM101 was found to be synthetic lethal with the cell wall integrity MAP Kinase SLT2 [27].
AgRim101 is not required for sporulation at an acidic starting pH since the Agrim101 mutant sporulates at approximately wild-type levels under standard laboratory sporulation-inducing conditions. Under these conditions, the metabolism of both wild-type and mutant strains achieves alkalization of the medium to slightly above pH 7.0. However, Agrim101 mutants fail to sporulate in alkaline media. Under these conditions, RIM101 mycelia are able to reduce the pH from alkaline towards neutral pH. In these aspects, Ashbya differs from the sporulation-deficient phenotypes seen in both S. cerevisiae and A. nidulans rim101 and pacC mutants [28].
Despite its redundancy at acidic/neutral pH, Rim101 clearly also regulates the adaptation at alkaline pH in A. gossypii, since both vegetative growth and sporulation are affected at pH 7.5–8.5 in the Agrim101 mutant.
In Ashbya, Rim101 may act as a repressor of sporulation-specific genes. However, in that case, more abundant sporulation would be expected in the Agrim101 mutant at least under acidic conditions. Such an oversporulation phenotype was observed, e.g., in Agste12 mutants but not in Agrim101 [15]. It is not known if AgRim101 shares the same DNA-binding site as ScRim101. If so, there is a potential Rim101-binding site in the IME1 promoter with the motif GCCAAG present 517 bp upstream of the start codon. This needs to be studied in more detail. In a promoter deletion analysis of AgIME1 we showed, however, that 491 bp of the intergenic region upstream of IME1 are sufficient for the wild-type-like transcriptional activation of IME1 [29]. With this, we can conclude that the presumptive Rim101-binding site at position -517 is not required for IME1 regulation since (i) the core promoter did not result in oversporulation and (ii) sporulation at acidic/neutral pH is not affected in Agrim101. RNAseq transcriptomics of non-sporulating vs. sporulating mycelia showed no differential regulation of RIM101 expression in Ashbya [29]. This suggests that activation of Rim101 requires pH-mediated processing similar to what has been described in S. cerevisiae.
5. Conclusions
This study shows that in A. gossypii the RIM pathway does not exert a dominant role in sporulation as described in S. cerevisiae. This and our previous findings indicate that sporulation in A. gossypii is mainly controlled by nutrients such as glucose and nitrogen. Extracellular signals including pH or pheromone signaling are not required for Ashbya to produce spores. A. gossypii is able to complete its life cycle, from a haploid spore that matures into a sporulating mycelium, without the need for a mating partner. This specialized lifestyle of A. gossypii with a close association to insect vectors may have put strong selection pressure on evolving a developmental program that allows for rapid sporulation based on limited environmental inputs.
Author Contributions
Conceptualization of the research, L.W. and J.W.; methodology, L.W. and J.W.; validation, L.W. and J.W.; formal analysis, L.W. and J.W.; investigation, L.W. and J.W.; resources, J.W.; data curation, L.W. and J.W.; writing—original draft preparation, L.W.; writing—review and editing, L.W. and J.W.; visualization, L.W. and J.W.; supervision, J.W.; project administration, J.W.; funding acquisition, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the European Union Marie Curie Initial Training Network Ariadne (237936). We acknowledge support by the German Research Foundation (Deutsche Forschungsgemeinschaft DFG)—project number 432888308—and the Open Access Publishing Fund of Geisenheim University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We wish to thank Klaus Lengeler and Andrea Walther for their support of this project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Penalva, M.A.; Arst, H.N., Jr. Recent advances in the characterization of ambient pH regulation of gene expression in filamentous fungi and yeasts. Annu. Rev. Microbiol. 2004, 58, 425–451. [Google Scholar] [CrossRef]
- Penalva, M.A.; Tilburn, J.; Bignell, E.; Arst, H.N., Jr. Ambient pH gene regulation in fungi: Making connections. Trends Microbiol. 2008, 16, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bignell, E.; Negrete-Urtasun, S.; Calcagno, A.M.; Haynes, K.; Arst, H.N., Jr.; Rogers, T. The Aspergillus ph-responsive transcription factor PacC regulates virulence. Mol. Microbiol. 2005, 55, 1072–1084. [Google Scholar] [CrossRef]
- Tilburn, J.; Sarkar, S.; Widdick, D.A.; Espeso, E.A.; Orejas, M.; Mungroo, J.; Penalva, M.A.; Arst, H.N., Jr. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid- and alkaline-expressed genes by ambient pH. EMBO J. 1995, 14, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.; Edwards, J.E., Jr.; Mitchell, A.P.; Ibrahim, A.S. Candida albicans Rim101 pH response pathway is required for host-pathogen interactions. Infect. Immun. 2000, 68, 5953–5959. [Google Scholar] [CrossRef] [Green Version]
- Ramon, A.M.; Porta, A.; Fonzi, W.A. Effect of environmental pH on morphological development of Candida albicans is mediated via the PacC-related transcription factor encoded by PRR2. J. Bacteriol. 1999, 181, 7524–7530. [Google Scholar] [CrossRef] [Green Version]
- Caracuel, Z.; Roncero, M.I.; Espeso, E.A.; Gonzalez-Verdejo, C.I.; Garcia-Maceira, F.I.; Di Pietro, A. The pH signalling transcription factor PacC controls virulence in the plant pathogen Fusarium oxysporum. Mol. Microbiol. 2003, 48, 765–779. [Google Scholar] [CrossRef]
- Landraud, P.; Chuzeville, S.; Billon-Grande, G.; Poussereau, N.; Bruel, C. Adaptation to pH and role of PacC in the rice blast fungus Magnaporthe oryzae. PLoS ONE 2013, 8, e69236. [Google Scholar] [CrossRef] [Green Version]
- Lamb, T.M.; Xu, W.; Diamond, A.; Mitchell, A.P. Alkaline response genes of Saccharomyces cerevisiae and their relationship to the RIM101 pathway. J. Biol. Chem. 2001, 276, 1850–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, T.M.; Mitchell, A.P. The transcription factor Rim101p governs ion tolerance and cell differentiation by direct repression of the regulatory genes NRG1 and SMP1 in Saccharomyces cerevisiae. Mol. Cell Biol. 2003, 23, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Zaman, S.; Lippman, S.I.; Zhao, X.; Broach, J.R. How Saccharomyces responds to nutrients. Annu. Rev. Genet. 2008, 42, 27–81. [Google Scholar] [CrossRef]
- Xu, W.; Mitchell, A.P. Yeast PalA/AIP1/Alix homolog Rim20p associates with a pest-like region and is required for its proteolytic cleavage. J. Bacteriol. 2001, 183, 6917–6923. [Google Scholar] [CrossRef] [Green Version]
- Diez, E.; Alvaro, J.; Espeso, E.A.; Rainbow, L.; Suarez, T.; Tilburn, J.; Arst, H.N., Jr.; Penalva, M.A. Activation of the Aspergillus PacC zinc finger transcription factor requires two proteolytic steps. EMBO J. 2002, 21, 1350–1359. [Google Scholar] [CrossRef] [Green Version]
- Broach, J.R. Nutritional control of growth and development in yeast. Genetics 2012, 192, 73–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, F.S.; Voegeli, S.; Brachat, S.; Lerch, A.; Gates, K.; Steiner, S.; Mohr, C.; Pohlmann, R.; Luedi, P.; Choi, S.; et al. The Ashbya gossypii genome as a tool for mapping the ancient Saccharomyces cerevisiae genome. Science 2004, 304, 304–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasserstrom, L.; Lengeler, K.B.; Walther, A.; Wendland, J. Molecular determinants of sporulation in Ashbya gossypii. Genetics 2013, 195, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Wasserstrom, L.; Lengeler, K.; Walther, A.; Wendland, J. Developmental growth control exerted via the protein a kinase Tpk2 in Ashbya gossypii. Eukaryot. Cell 2015, 14, 593–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendland, J. Sporulation in Ashbya gossypii. J. Fungi 2020, 6, 157. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Q.; Liu, C.; Shang, Y.; Wen, F.; Wang, F.; Liu, W.; Xiao, W.; Li, W. A role for the respiratory chain in regulating meiosis initiation in Saccharomyces cerevisiae. Genetics 2018, 208, 1181–1194. [Google Scholar] [CrossRef] [Green Version]
- Wendland, J.; Ayad-Durieux, Y.; Knechtle, P.; Rebischung, C.; Philippsen, P. PCR-based gene targeting in the filamentous fungus Ashbya gossypii. Gene 2000, 242, 381–391. [Google Scholar] [CrossRef]
- Walther, A.; Wendland, J. PCR-based gene targeting in Candida albicans. Nat. Protoc. 2008, 3, 1414–1421. [Google Scholar] [CrossRef]
- Gattiker, A.; Rischatsch, R.; Demougin, P.; Voegeli, S.; Dietrich, F.S.; Philippsen, P.; Primig, M. Ashbya genome database 3.0: A cross-species genome and transcriptome browser for yeast biologists. BMC Genom. 2007, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Mitchell, A.P. Proteolytic activation of Rim1p, a positive regulator of yeast sporulation and invasive growth. Genetics 1997, 145, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Su, S.S.; Mitchell, A.P. Molecular characterization of the yeast meiotic regulatory gene RIM1. Nucleic Acids Res. 1993, 21, 3789–3797. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.C.; Zamarbide-Fores, S.; Pedelini, L.; Llopis-Torregrosa, V.; Yenush, L. A functional Rim101 complex is required for proper accumulation of the Ena1 na+-atpase protein in response to salt stress in Saccharomyces cerevisiae. FEMS Yeast Res. 2015, 15, fov017. [Google Scholar] [CrossRef] [Green Version]
- Lengeler, K.B.; Wasserstrom, L.; Walther, A.; Wendland, J. Analysis of the cell wall integrity pathway of Ashbya gossypii. Microbiol Res. 2013, 168, 607–614. [Google Scholar] [CrossRef]
- Castrejon, F.; Gomez, A.; Sanz, M.; Duran, A.; Roncero, C. The Rim101 pathway contributes to yeast cell wall assembly and its function becomes essential in the absence of mitogen-activated protein kinase Slt2p. Eukaryot. Cell 2006, 5, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Piccirillo, S.; White, M.G.; Murphy, J.C.; Law, D.J.; Honigberg, S.M. The Rim101p/PacC pathway and alkaline pH regulate pattern formation in yeast colonies. Genetics 2010, 184, 707–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasserstrom, L.; Dunkler, A.; Walther, A.; Wendland, J. The APSES protein Sok2 is a positive regulator of sporulation in Ashbya gossypii. Mol. Microbiol. 2017, 106, 949–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Induction of sporulation in response to alkaline pH in S. cerevisiae. When a non-fermentable carbon source is metabolized by respiration the external pH increases. The alkaline pH induces proteolytic activation of Rim101 by Rim13/Rim20. Rim101 then activates IME1 transcription by repressing transcription of the repressor Smp1 [14].
Figure 1.
Induction of sporulation in response to alkaline pH in S. cerevisiae. When a non-fermentable carbon source is metabolized by respiration the external pH increases. The alkaline pH induces proteolytic activation of Rim101 by Rim13/Rim20. Rim101 then activates IME1 transcription by repressing transcription of the repressor Smp1 [14].
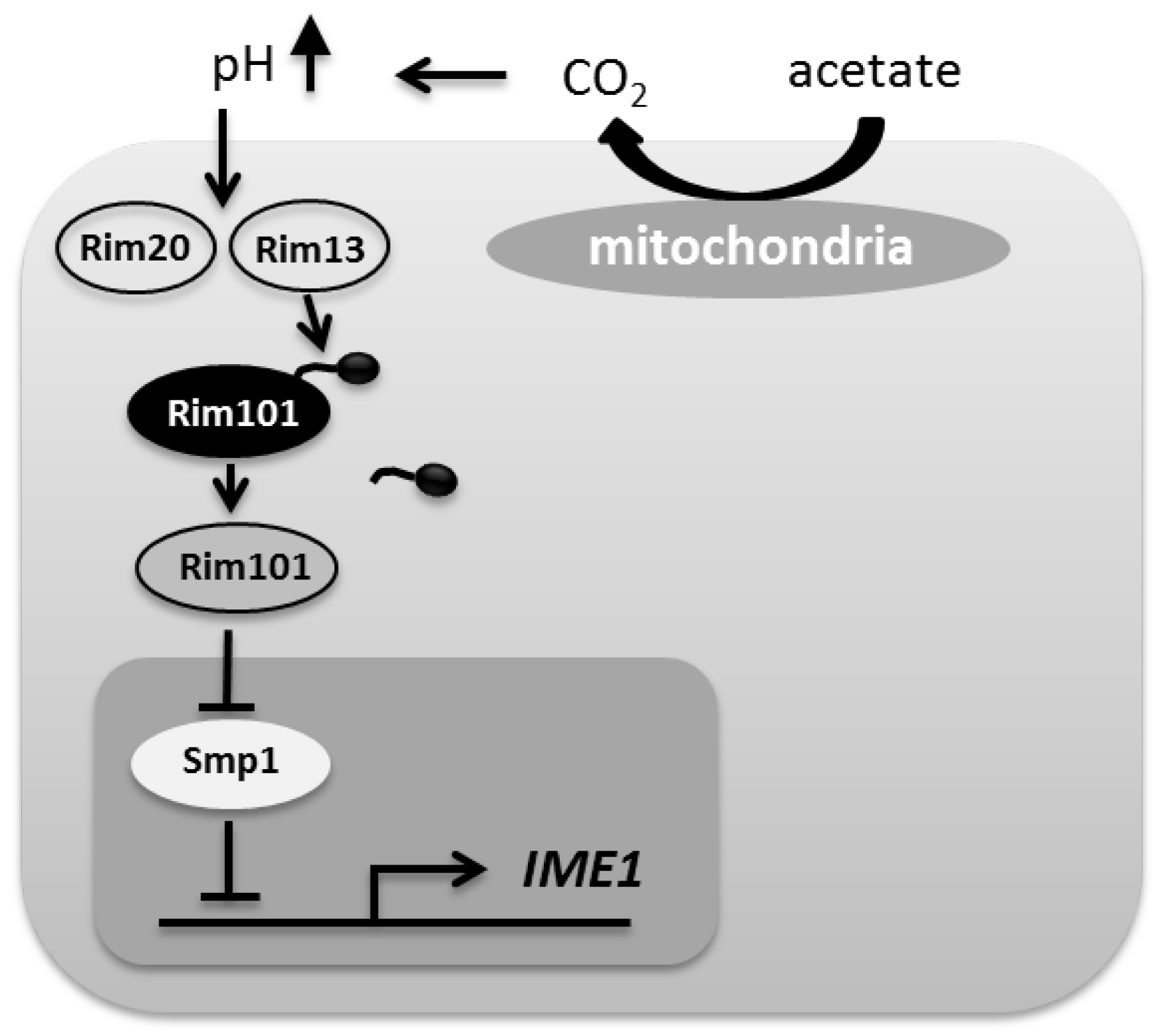
Figure 2.
Alignment of the zinc finger regions Rim101/PacC proteins of S. cerevisiae, A. gossypii, and Aspergillus nidulans. The conserved cysteine and histidine residues of the three C2H2 zinc-chelating residues are marked with asterisks [4].
Figure 2.
Alignment of the zinc finger regions Rim101/PacC proteins of S. cerevisiae, A. gossypii, and Aspergillus nidulans. The conserved cysteine and histidine residues of the three C2H2 zinc-chelating residues are marked with asterisks [4].

Figure 3.
Radial growth of the Ashbya parental (WT) strain and the Agrim101 mutant (AWL63) on full medium plates after one week of growth at the indicated temperatures.
Figure 3.
Radial growth of the Ashbya parental (WT) strain and the Agrim101 mutant (AWL63) on full medium plates after one week of growth at the indicated temperatures.
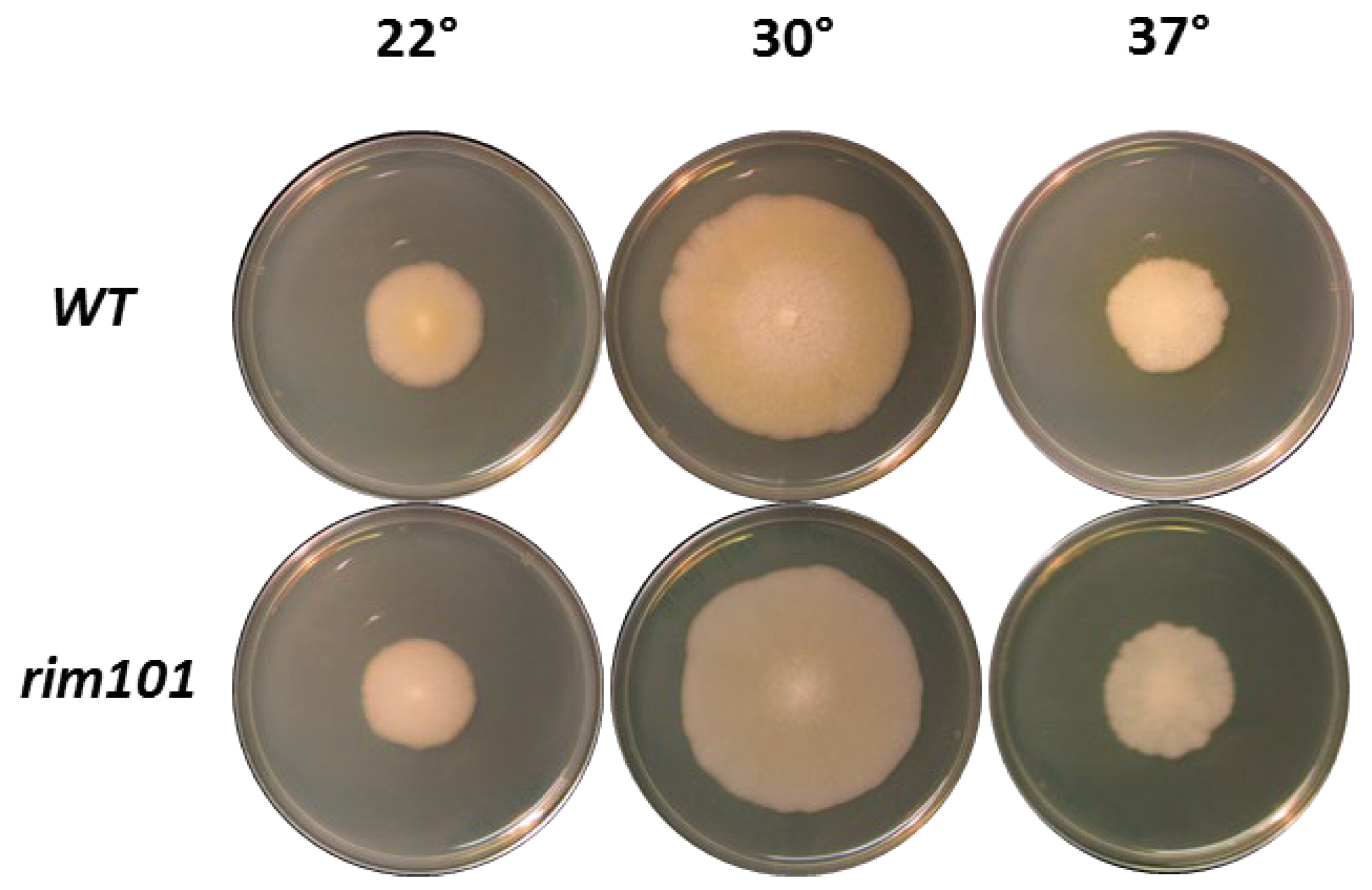
Figure 4.
Sporulation ability of the A. gossypii wild-type strain and rim101 mutant at different pH. (A) Spores were isolated from the central part of colonies grown for 10 days on a full medium plate. (B,C) Sporulation of mycelia on solid media buffered with Tris-HCl to different pH values. (B) Spore-forming units were determined by dilution plating of spore suspensions derived from the sporulation zones. (C) Sporulation zones of mycelia were marked with white-dotted circles.
Figure 4.
Sporulation ability of the A. gossypii wild-type strain and rim101 mutant at different pH. (A) Spores were isolated from the central part of colonies grown for 10 days on a full medium plate. (B,C) Sporulation of mycelia on solid media buffered with Tris-HCl to different pH values. (B) Spore-forming units were determined by dilution plating of spore suspensions derived from the sporulation zones. (C) Sporulation zones of mycelia were marked with white-dotted circles.
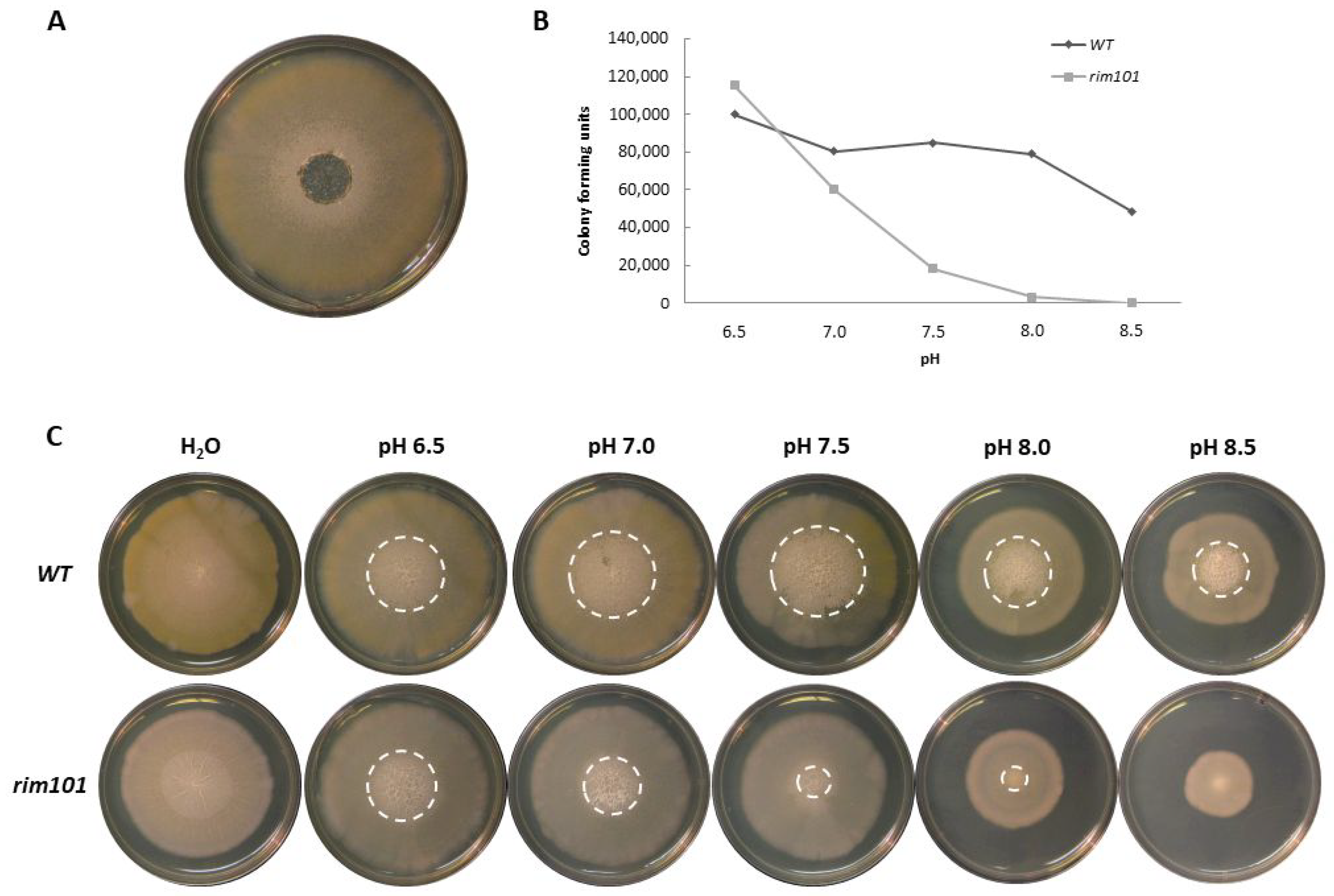
Figure 5.
Liquid sporulation assays at acidic and alkaline pH. The indicated strains were pre-grown in AFM and then transferred to CSM-sporulation media (acidic = unbuffered with a starting pH of 5.3; alkaline = Tris-buffered to a pH of 8.6 at room temperature). After one week of incubation at 28 °C, samples were analyzed by microscopy. Representative images are shown. Clumps of spores (indicated by black arrows in the WT) were abundant in sporulating strains but completely absent in homokaryotic rim101 mutants showing only hyphal filaments (indicated by white arrows). Scale bars are 10 µm.
Figure 5.
Liquid sporulation assays at acidic and alkaline pH. The indicated strains were pre-grown in AFM and then transferred to CSM-sporulation media (acidic = unbuffered with a starting pH of 5.3; alkaline = Tris-buffered to a pH of 8.6 at room temperature). After one week of incubation at 28 °C, samples were analyzed by microscopy. Representative images are shown. Clumps of spores (indicated by black arrows in the WT) were abundant in sporulating strains but completely absent in homokaryotic rim101 mutants showing only hyphal filaments (indicated by white arrows). Scale bars are 10 µm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains used in this study. A. gossypii strains used in this study. All strains are derivatives of Agleu2.
Table 1.
Strains used in this study. A. gossypii strains used in this study. All strains are derivatives of Agleu2.
| No | Strain | Genotype | Source |
|---|---|---|---|
| 71 | ATCC10895 | Agleu2 | Lab collection |
| C420 | AWL13 | leu2, rim101:: GEN3/RIM101 heterokaryon | This study |
| C510 | AWL62 | leu2, rim101:: GEN3/RIM101 heterokaryon | This study |
| C511 | AWL63 | leu2, rim101:: GEN3 homokaryon derived from AWL62 | This study |
| C512 | AWL64 | leu2, rim101:: GEN3 homokaryon derived from C420 | This study |
Table 2.
Oligonucleotide primers used in this study.
| Oligonucleotide | Sequence 5′-to-3′ * |
|---|---|
| #1214-G2 | GGGTAATTTGTCGCGGTCTGGG |
| #1215-G3 | GCCCATCAGATTGATGTCCTCC |
| #4603-G1-RIM101 | GCTGCTATCGGACGCAGC |
| #4604-G4-RIM101 | GTGGAGTGCGACTTCTCC |
| #4605-S1-RIM101 | ACGATTATACGGCCAGCCAAATAGCAAGCGCGTTACTGCATGAACgaagcttcgtacgctgcaggtc |
| #4606-S2-RIM101 | ATTATGTTCACAGTCTGGAGTTCTGCACGGAGGGCATCAGTGTCGctgatatcatcgatgaattcgag |
| #4607-I1-RIM101 | CACTTATGCCAGGATCACG |
| #4608-I2-RIM101 | GCTCCCGGAGAGGTCACC |
* Upper-case sequences correspond to A. gossypii DNA sequences and lower-case sequences correspond to annealing regions on pFA cassettes. All sequences are written from 5′ to 3′.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wasserstrom, L.; Wendland, J. Role of RIM101 for Sporulation at Alkaline pH in Ashbya gossypii. J. Fungi 2021, 7, 527. https://doi.org/10.3390/jof7070527
AMA Style
Wasserstrom L, Wendland J. Role of RIM101 for Sporulation at Alkaline pH in Ashbya gossypii. Journal of Fungi. 2021; 7(7):527. https://doi.org/10.3390/jof7070527
Chicago/Turabian StyleWasserstrom, Lisa, and Jürgen Wendland. 2021. "Role of RIM101 for Sporulation at Alkaline pH in Ashbya gossypii" Journal of Fungi 7, no. 7: 527. https://doi.org/10.3390/jof7070527
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.