
Specific Functional Features of the Cell Integrity MAP Kinase Pathway in the Dimorphic Fission Yeast Schizosaccharomyces japonicus

 , , , and
, , , and 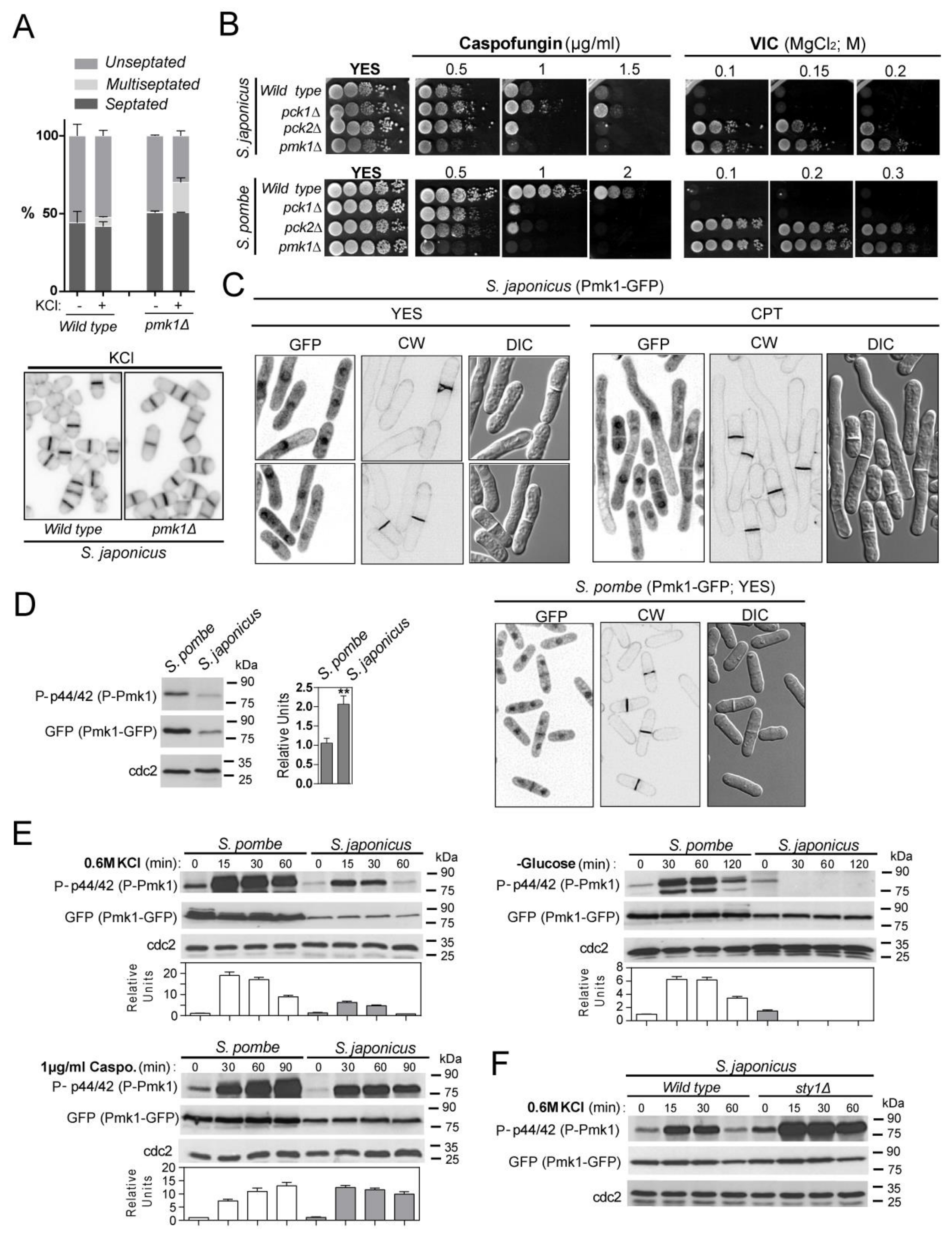{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Growth Conditions, and Reagents
2.2. Gene Disruption and Gene Fusion
2.3. Construction of S. pombe and S. japonicus Strains Expressing Wild-Type and Chimeric Pmk1–HA Fusions
2.4. Detection and Quantification of Activated and Total Pmk1 Levels
2.5. Co-immunoprecipitation
2.6. Plate Assay of Stress Sensitivity for Growth
2.7. Quantification of Mycelial Growth during Nutritional Stress
2.8. Microscopy Observation
3. Results
3.1. Environmental Control of S. japonicus Cell Integrity MAPK Pathway by the PKC Orthologs Pck1 and Pck2
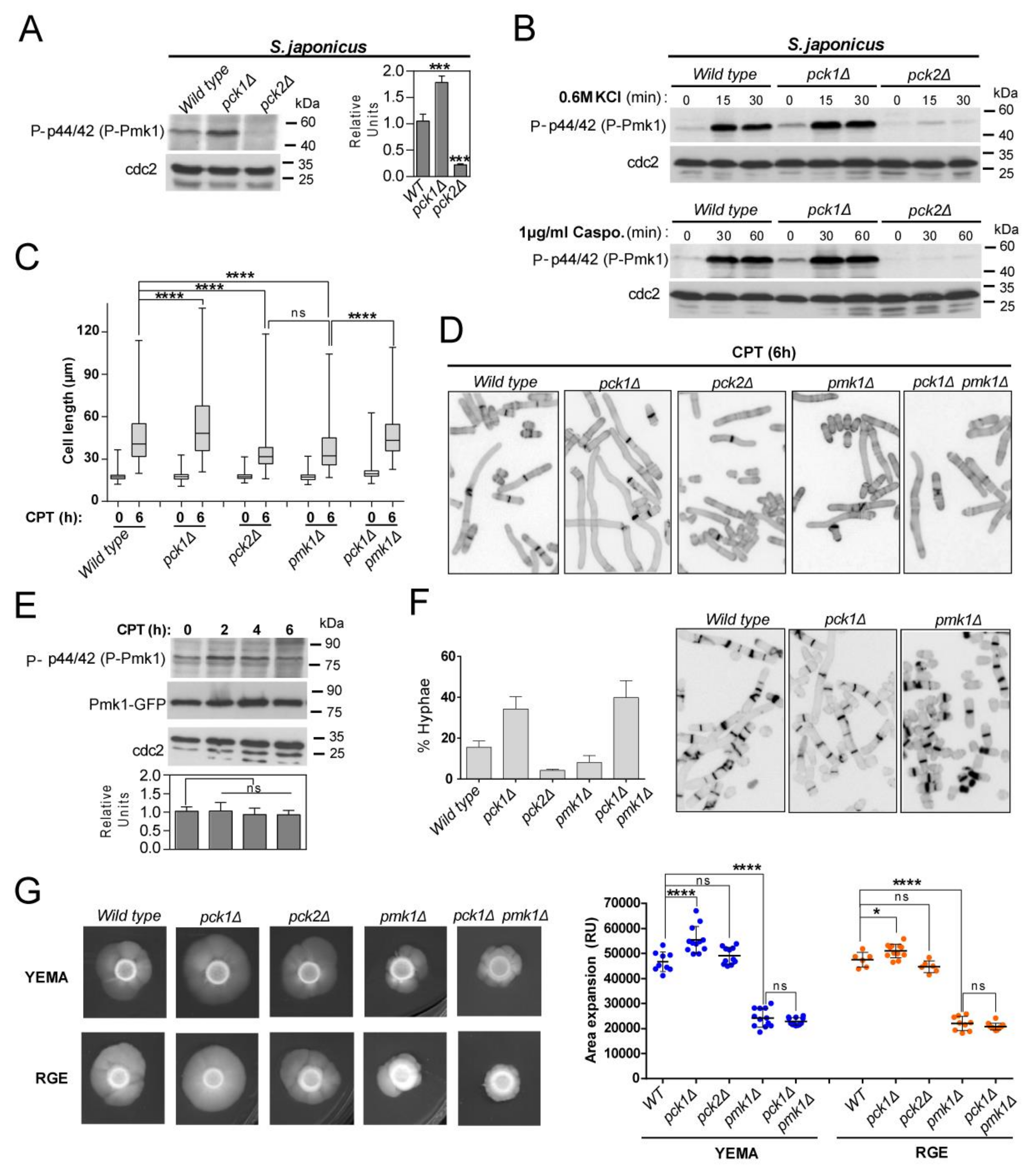
3.2. Fine-Tuning of Pmk1 Activity by Pck1 and Pck2 Modulates Antagonistically S. japonicus Hyphal Differentiation
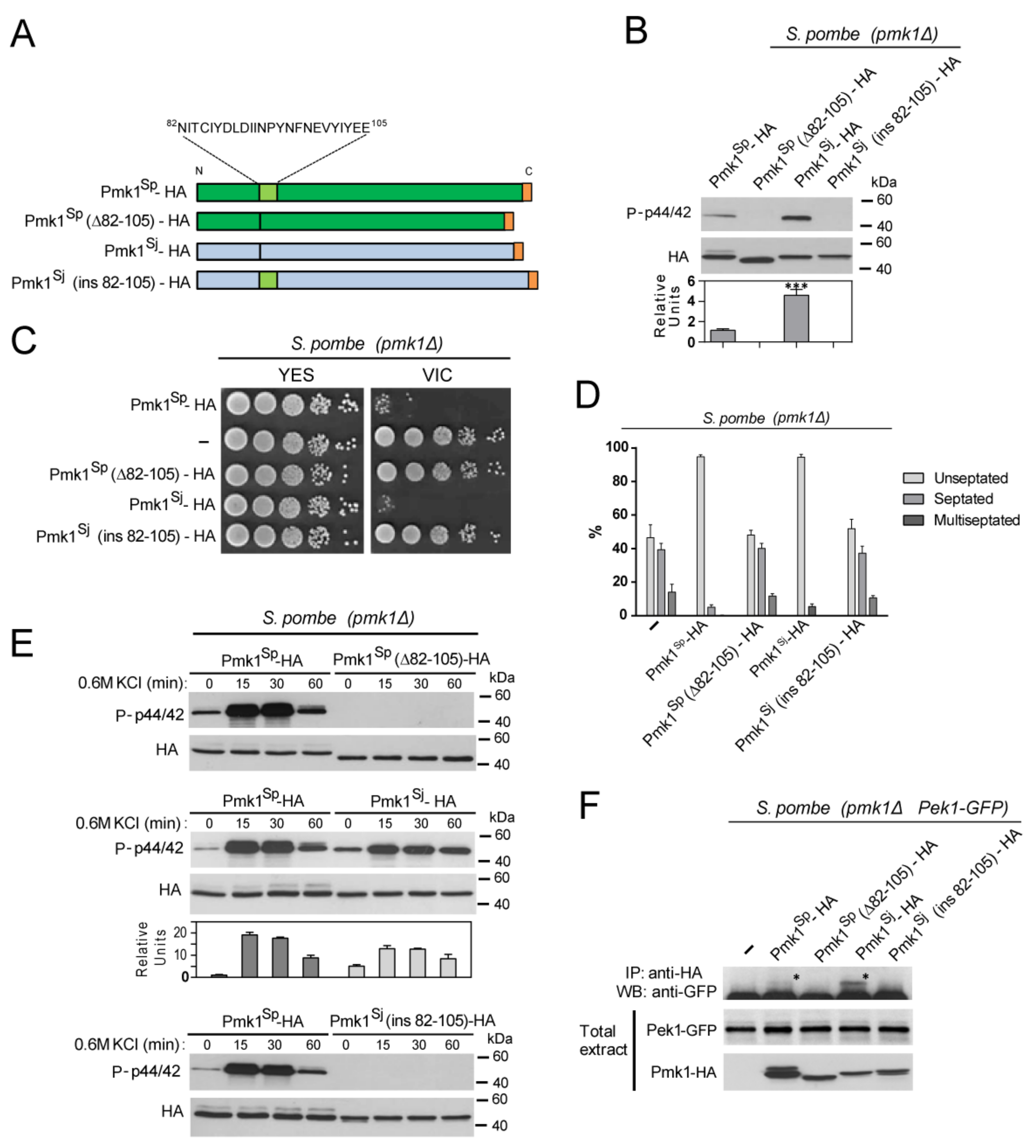
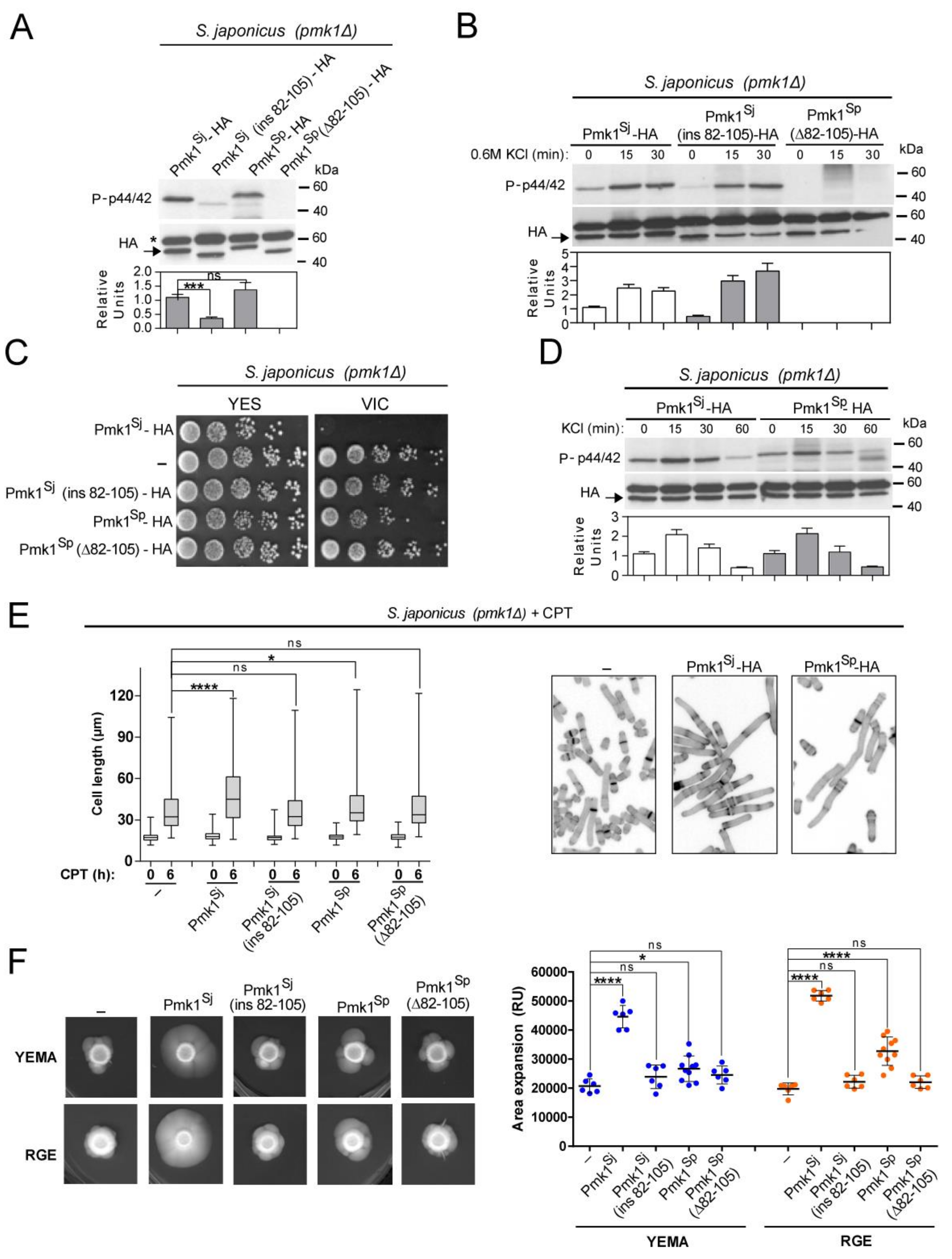
3.3. Species-Specific N-lobe Domain Architecture Accounts for Differential Control of Pmk1 Function in S. pombe and S. japonicus during Growth, Stress, and Hyphal Differentiation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perez, P.; Cansado, J. Cell Integrity Signaling and Response to Stress in Fission Yeast. Curr. Protein Pept. Sci. 2010, 11, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Kabeche, R.; Madrid, M.; Cansado, J.; Moseley, J.B. Eisosomes Regulate Phosphatidylinositol 4,5-Bisphosphate (PI(4,5)P2) Cortical Clusters and Mitogen-activated Protein (MAP) Kinase Signaling upon Osmotic Stress. J. Biol. Chem. 2015, 290, 25960–25973. [Google Scholar] [CrossRef] [PubMed]
- Madrid, M.; Fernandez-Zapata, J.; Sanchez-Mir, L.; Soto, T.; Franco, A.; Vicente-Soler, J.; Gacto, M.; Cansado, J. Role of the fission yeast cell integrity MAPK pathway in response to glucose limitation. BMC Microbiol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Madrid, M.; Nunez, A.; Soto, T.; Vicente-Soler, J.; Gacto, M.; Cansado, J. Stress-activated protein kinase-mediated down-regulation of the cell integrity pathway mitogen-activated protein kinase Pmk1p by protein phosphatases. Mol. Biol. Cell 2007, 18, 4405–4419. [Google Scholar] [CrossRef]
- Madrid, M.; Soto, T.; Khong, H.; Franco, A.; Vicente, J.; Perez, P.; Gacto, M.; Cansado, J. Stress-induced response, localization, and regulation of the Pmk1 cell integrity pathway in Schizosaccharomyces pombe. J. Biol. Chem. 2006, 281, 2033–2043. [Google Scholar] [CrossRef]
- Barba, G.; Soto, T.; Madrid, M.; Nunez, A.; Vicente, J.; Gacto, M.; Cansado, J.; Grp, Y.P.; Grp, Y.P. Activation of the cell integrity pathway is channelled through diverse signalling elements in fission yeast. Cell. Signal. 2008, 20, 748–757. [Google Scholar] [CrossRef]
- Sanchez-Mir, L.; Soto, T.; Franco, A.; Madrid, M.; Viana, R.; Vicente, J.; Gacto, M.; Perez, P.; Cansado, J. Rho1 GTPase and PKC Ortholog Pck1 Are Upstream Activators of the Cell Integrity MAPK Pathway in Fission Yeast. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Nunez, A.; Franco, A.; Madrid, M.; Soto, T.; Vicente, J.; Gacto, M.; Cansado, J. Role for RACK1 Orthologue Cpc2 in the Modulation of Stress Response in Fission Yeast. Mol. Biol. Cell 2009, 20, 3996–4009. [Google Scholar] [CrossRef]
- Soto, T.; Villar-Tajadura, M.; Madrid, M.; Vicente, J.; Gacto, M.; Perez, P.; Cansado, J. Rga4 Modulates the Activity of the Fission Yeast Cell Integrity MAPK Pathway by Acting as a Rho2 GTPase-activating Protein. J. Biol. Chem. 2010, 285, 11516–11525. [Google Scholar] [CrossRef]
- Villar-Tajadura, M.; Coll, P.; Madrid, M.; Cansado, J.; Santos, B.; Perez, P. Rga2 is a Rho2 GAP that regulates morphogenesis and cell integrity in S-pombe. Mol. Microbiol. 2008, 70, 867–881. [Google Scholar] [CrossRef]
- Sanchez-Mir, L.; Franco, A.; Martin-Garcia, R.; Madrid, M.; Vicente-Soler, J.; Soto, T.; Gacto, M.; Perez, P.; Cansado, J. Rho2 palmitoylation is required for plasma membrane localization and proper signaling to the fission yeast cell integrity MAPK pathway. Mol. Cell. Biol. 2014. [Google Scholar] [CrossRef]
- Garcia, P.; Tajadura, V.; Sanchez, Y. The Rho1p exchange factor Rgf1p signals upstream from the Pmk1 mitogen-activated protein kinase pathway in fission yeast. Mol. Biol. Cell 2009, 20, 721–731. [Google Scholar] [CrossRef][Green Version]
- Madrid, M.; Vazquez-Marin, B.; Franco, A.; Soto, T.; Vicente-Soler, J.; Gacto, M.; Cansado, J. Multiple crosstalk between TOR and the cell integrity MAPK signaling pathway in fission yeast. Sci. Rep. 2016, 6, 37515. [Google Scholar] [CrossRef]
- Prieto-Ruiz, F.; Vicente-Soler, J.; Franco, A.; Gomez-Gil, E.; Sanchez-Marinas, M.; Vazquez-Marin, B.; Aligue, R.; Madrid, M.; Moreno, S.; Soto, T.; et al. RNA-Binding Protein Rnc1 Regulates Cell Length at Division and Acute Stress Response in Fission Yeast through Negative Feedback Modulation of the Stress-Activated Mitogen-Activated Protein Kinase Pathway. mBio 2020, 11. [Google Scholar] [CrossRef]
- Makarova, M.; Peter, M.; Balogh, G.; Glatz, A.; MacRae, J.I.; Lopez Mora, N.; Booth, P.; Makeyev, E.; Vigh, L.; Oliferenko, S. Delineating the Rules for Structural Adaptation of Membrane-Associated Proteins to Evolutionary Changes in Membrane Lipidome. Curr. Biol. 2020, 30, 367–380.e368. [Google Scholar] [CrossRef]
- Gu, Y.; Yam, C.; Oliferenko, S. Divergence of mitotic strategies in fission yeasts. Nucleus 2012, 3, 220–225. [Google Scholar] [CrossRef]
- Kaino, T.; Tonoko, K.; Mochizuki, S.; Takashima, Y.; Kawamukai, M. Schizosaccharomyces japonicus has low levels of CoQ10 synthesis, respiration deficiency, and efficient ethanol production. Biosci. Biotechnol. Biochem. 2018, 82, 1031–1042. [Google Scholar] [CrossRef]
- Sipiczki, M.; Takeo, K.; Yamaguchi, M.; Yoshida, S.; Miklos, I. Environmentally controlled dimorphic cycle in a fission yeast. Microbiology 1998, 144 Pt 5, 1319–1330. [Google Scholar] [CrossRef]
- Papp, L.; Sipiczki, M.; Holb, I.J.; Miklos, I. Optimal conditions for mycelial growth of Schizosaccharomyces japonicus cells in liquid medium: It enables the molecular investigation of dimorphism. Yeast 2014, 31, 475–482. [Google Scholar] [CrossRef]
- Aoki, K.; Furuya, K.; Niki, H. Schizosaccharomyces japonicus: A Distinct Dimorphic Yeast among the Fission Yeasts. Cold Spring Harb. Protoc. 2017, 2017, 082651. [Google Scholar] [CrossRef]
- Niki, H. Induction of Hyphal Growth in Schizosaccharomyces japonicus. Cold Spring Harb. Protoc. 2017, 2017, 091868. [Google Scholar] [CrossRef]
- Nozaki, S.; Furuya, K.; Niki, H. The Ras1-Cdc42 pathway is involved in hyphal development of Schizosaccharomyces japonicus. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Gomez-Gil, E.; Franco, A.; Madrid, M.; Vazquez-Marin, B.; Gacto, M.; Fernandez-Breis, J.; Vicente-Soler, J.; Soto, T.; Cansado, J. Quorum sensing and stress-activated MAPK signaling repress yeast to hypha transition in the fission yeast Schizosaccharomyces japonicus. PLoS Genet. 2019, 15, e1008192. [Google Scholar] [CrossRef]
- Moreno, S.; Klar, A.; Nurse, P. Molecular Genetic-Analysis Of Fission Yeast Schizosaccharomyces pombe. Methods Enzymol. 1991, 194, 795–823. [Google Scholar]
- Petersen, J.; Russell, P. Growth and the Environment of Schizosaccharomyces pombe. Cold Spring Harb. Protoc. 2016, 2016, 079764. [Google Scholar] [CrossRef]
- Kinnaer, C.; Dudin, O.; Martin, S.G. Yeast-to-hypha transition of Schizosaccharomyces japonicus in response to environmental stimuli. Mol. Biol. Cell 2019, 30, 975–991. [Google Scholar] [CrossRef]
- Aoki, K.; Niki, H. Transformation of Schizosaccharomyces japonicus. Cold Spring Harb. Protoc. 2017, 2017, 091850. [Google Scholar] [CrossRef]
- Furuya, K.; Niki, H. Isolation of heterothallic haploid and auxotrophic mutants of Schizosaccharomyces japonicus. Yeast 2009, 26, 221–233. [Google Scholar] [CrossRef]
- Hentges, P.; Van Driessche, B.; Tafforeau, L.; Vandenhaute, J.; Carr, A.M. Three novel antibiotic marker cassettes for gene disruption and marker switching in Schizosaccharomyces pombe. Yeast 2005, 22, 1013–1019. [Google Scholar] [CrossRef]
- Krawchuk, M.D.; Wahls, W.P. High-efficiency gene targeting in Schizosaccharomyces pombe using a modular, PCR-based approach with long tracts of flanking homology. Yeast 1999, 15, 1419–1427. [Google Scholar] [CrossRef]
- Madrid, M.; Jimenez, R.; Sanchez-Mir, L.; Soto, T.; Franco, A.; Vicente-Soler, J.; Gacto, M.; Perez, P.; Cansado, J. Multiple layers of regulation influence cell integrity control by the PKC ortholog Pck2 in fission yeast. J. Cell. Sci. 2015, 128, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Hagan, I.M. Immunofluorescence Microscopy of Schizosaccharomyces pombe Using Chemical Fixation. Cold Spring Harb. Protoc. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Mir, L.; Franco, A.; Madrid, M.; Vicente-Soler, J.; Antonia Villar-Tajadura, M.; Soto, T.; Perez, P.; Gacto, M.; Cansado, J. Biological Significance of Nuclear Localization of Mitogen-activated Protein Kinase Pmk1 in Fission Yeast. J. Biol. Chem. 2012, 287, 26038–26051. [Google Scholar] [CrossRef]
- Edreira, T.; Celador, R.; Manjón, E.; Sánchez, Y. A novel checkpoint pathway controls actomyosin ring constriction trigger in fission yeast. eLife 2020, 9. [Google Scholar] [CrossRef]
- Zaitsevskaya-Carter, T.; Cooper, J.A. Spm1, a stress-activated MAP kinase that regulates morphogenesis in S.pombe. EMBO J. 1997, 16, 1318–1331. [Google Scholar] [CrossRef]
- Sugiura, R.; Toda, T.; Dhut, S.; Shuntoh, H.; Kuno, T. The MAPK kinase Pek1 acts as a phosphorylation-dependent molecular switch. Nature 1999, 399, 479–483. [Google Scholar] [CrossRef]
- Toda, T.; Dhut, S.; SupertiFurga, G.; Gotoh, Y.; Nishida, E.; Sugiura, R.; Kuno, T. The fission yeast pmk1(+) gene encodes a novel mitogen-activated protein kinase homolog which regulates cell integrity and functions coordinately with the protein kinase C pathway. Mol. Cell. Biol. 1996, 16, 6752–6764. [Google Scholar] [CrossRef]
- Madrid, M.; Vazquez-Marin, B.; Soto, T.; Franco, A.; Gomez-Gil, E.; Vicente-Soler, J.; Gacto, M.; Perez, P.; Cansado, J. Differential functional regulation of PKC orthologs in fission yeast. J. Biol. Chem. 2017. [Google Scholar] [CrossRef]
- Arellano, M.; Valdivieso, M.; Calonge, T.; Coll, P.; Duran, A.; Perez, P. Schizosaccharomyces pombe protein kinase C homologues, pck1p and pck2p, are targets of rho1p and rho2p and differentially regulate cell integrity. J. Cell Sci. 1999, 112, 3569–3578. [Google Scholar] [CrossRef]
- Calonge, T.; Nakano, K.; Arellano, M.; Arai, R.; Katayama, S.; Toda, T.; Mabuchi, I.; Perez, P. Schizosaccharomyces pombe Rho2p GTPase regulates cell wall alpha-glucan biosynthesis through the protein kinase Pck2p. Mol. Biol. Cell 2000, 11, 4393–4401. [Google Scholar] [CrossRef]
- Gu, Y.; Oliferenko, S. Comparative biology of cell division in the fission yeast clade. Curr. Opin. Microbiol. 2015, 28, 18–25. [Google Scholar] [CrossRef]
- Kumamoto, C.A. A contact-activated kinase signals Candida albicans invasive growth and biofilm development. Proc. Natl. Acad. Sci. USA 2005, 102, 5576–5581. [Google Scholar] [CrossRef]
- Monge, R.A.; Román, E.; Nombela, C.; Pla, J. The MAP kinase signal transduction network in Candida albicans. Microbiology 2006, 152, 905–912. [Google Scholar] [CrossRef]
- Kraus, P.R.; Fox, D.S.; Cox, G.M.; Heitman, J. The Cryptococcus neoformans MAP kinase Mpk1 regulates cell integrity in response to antifungal drugs and loss of calcineurin function. Mol. Microbiol. 2003, 48, 1377–1387. [Google Scholar] [CrossRef]
- Birkaya, B.; Maddi, A.; Joshi, J.; Free, S.J.; Cullen, P.J. Role of the cell wall integrity and filamentous growth mitogen-activated protein kinase pathways in cell wall remodeling during filamentous growth. Eukaryot. Cell 2009, 8, 1118–1133. [Google Scholar] [CrossRef]
- Kumar, G.S.; Clarkson, M.W.; Kunze, M.B.A.; Granata, D.; Wand, A.J.; Lindorff-Larsen, K.; Page, R.; Peti, W. Dynamic activation and regulation of the mitogen-activated protein kinase p38. Proc. Natl. Acad. Sci. USA 2018, 115, 4655–4660. [Google Scholar] [CrossRef]
- Nguyen, T.; Ruan, Z.; Oruganty, K.; Kannan, N. Co-conserved MAPK features couple D-domain docking groove to distal allosteric sites via the C-terminal flanking tail. PLoS ONE 2015, 10, e0119636. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Gil, E.; Franco, A.; Vázquez-Marín, B.; Prieto-Ruiz, F.; Pérez-Díaz, A.; Vicente-Soler, J.; Madrid, M.; Soto, T.; Cansado, J. Specific Functional Features of the Cell Integrity MAP Kinase Pathway in the Dimorphic Fission Yeast Schizosaccharomyces japonicus. J. Fungi 2021, 7, 482. https://doi.org/10.3390/jof7060482
Gómez-Gil E, Franco A, Vázquez-Marín B, Prieto-Ruiz F, Pérez-Díaz A, Vicente-Soler J, Madrid M, Soto T, Cansado J. Specific Functional Features of the Cell Integrity MAP Kinase Pathway in the Dimorphic Fission Yeast Schizosaccharomyces japonicus. Journal of Fungi. 2021; 7(6):482. https://doi.org/10.3390/jof7060482
Chicago/Turabian StyleGómez-Gil, Elisa, Alejandro Franco, Beatriz Vázquez-Marín, Francisco Prieto-Ruiz, Armando Pérez-Díaz, Jero Vicente-Soler, Marisa Madrid, Teresa Soto, and José Cansado. 2021. "Specific Functional Features of the Cell Integrity MAP Kinase Pathway in the Dimorphic Fission Yeast Schizosaccharomyces japonicus" Journal of Fungi 7, no. 6: 482. https://doi.org/10.3390/jof7060482
APA StyleGómez-Gil, E., Franco, A., Vázquez-Marín, B., Prieto-Ruiz, F., Pérez-Díaz, A., Vicente-Soler, J., Madrid, M., Soto, T., & Cansado, J. (2021). Specific Functional Features of the Cell Integrity MAP Kinase Pathway in the Dimorphic Fission Yeast Schizosaccharomyces japonicus. Journal of Fungi, 7(6), 482. https://doi.org/10.3390/jof7060482