
Trichoderma longibrachiatum and Trichoderma asperellum Confer Growth Promotion and Protection against Late Wilt Disease in the Field



Abstract

1. Introduction
2. Materials and Methods
2.1. Origin and Growth of Magnaporthiopsis maydis
2.2. Origin and Growth of the Trichoderma Species
2.3. Overall Description of the Field Experiments
2.3.1. Sowing and Irrigation Regime
2.3.2. Complementary Infection Method
2.3.3. Trichoderma-Based Biocontrol Treatments
2.4. Trichoderma spp. Late Wilt Control in the 2019 Growing Season
2.5. Trichoderma spp. Late Wilt Control in the 2020 Growing Season
2.5.1. Enhancing the Disease Using Wooden Toothpicks Inoculation
2.5.2. The 2020 Experiment
2.6. qPCR Diagnosis of M. maydis DNA in the Maize Plants
2.7. Statistical Analysis
3. Results
3.1. Trichoderma spp. Late Wilt Control in the 2019 Growing Season
3.2. Trichoderma spp. Late Wilt Control in the 2020 Growing Season
3.2.1. Enhancing the Disease Using Wooden Toothpicks Inoculation
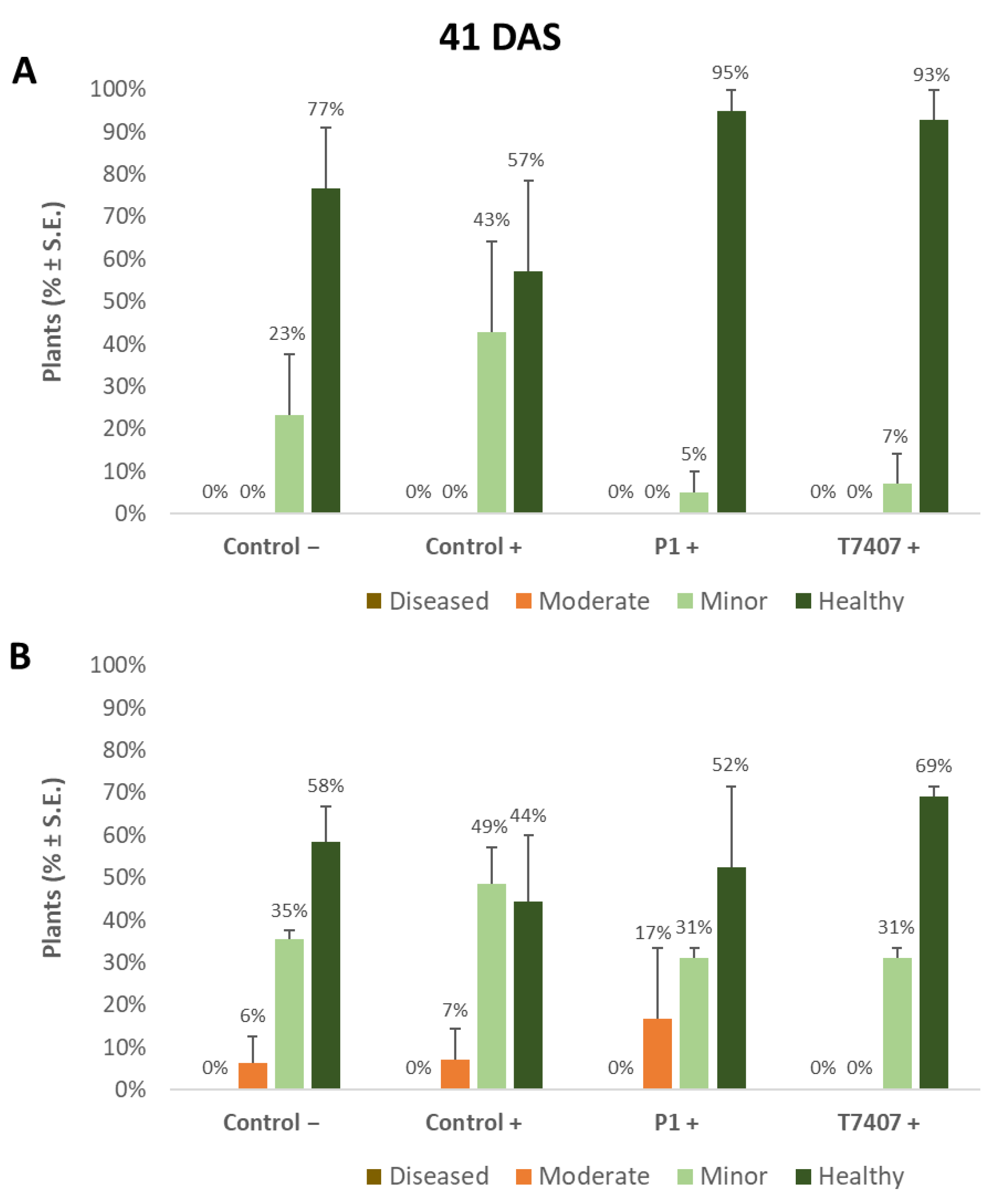
3.2.2. The 2020 Experiment Results at the Sprouting Growth Phase
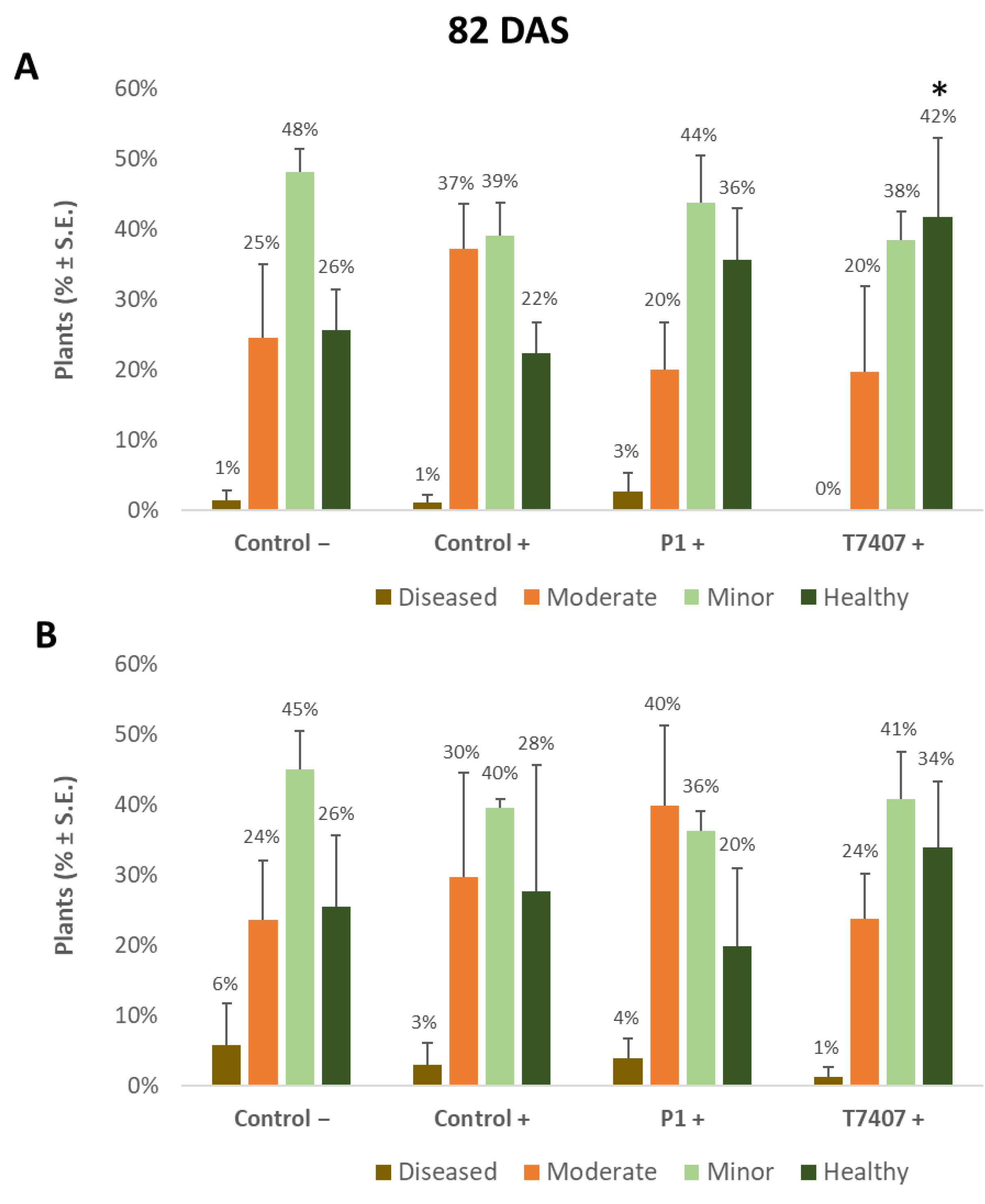
3.2.3. The 2020 Experiment Results at the Harvest
3.2.4. Lower Stem Symptoms Evaluation in the 2020 Growing Season
3.2.5. Cobs’ Spathes Symptoms Evaluation in the 2020 Growing Season
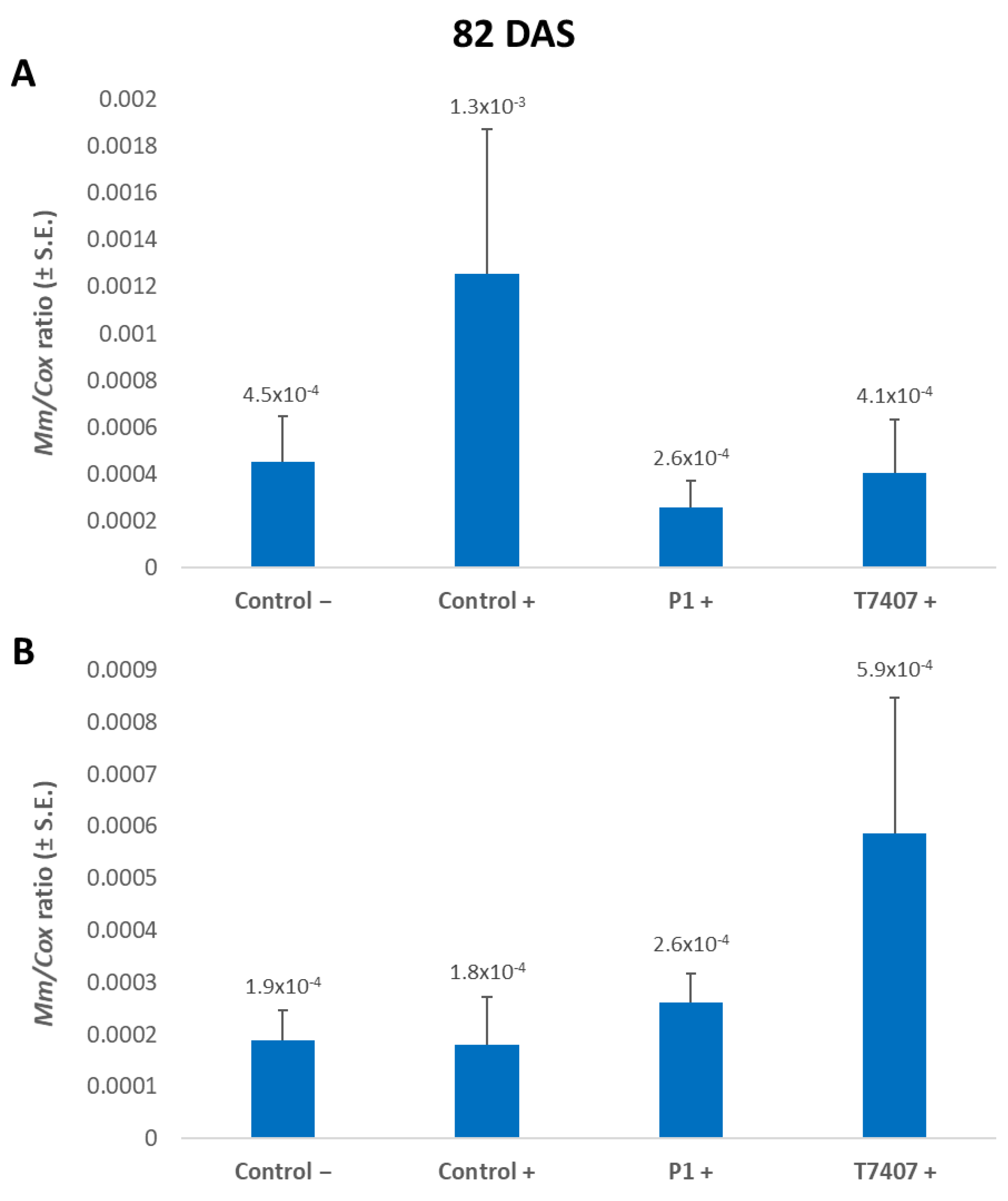
3.2.6. M. maydis DNA Evaluation at the 2020 Season’s End
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.Z.; Crous, P.W.; de Vries, R.P.; Lebrun, M.H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 2014, 79, 85–120. [Google Scholar] [CrossRef] [PubMed]
- Samra, A.S.; Sabet, K.A.; Hingorani, M.K. A new wilt disease of maize in Egypt. Plant Dis. Rep. 1962, 46, 481–483. [Google Scholar]
- Abdel-Hamid, M.S.; Abdel-Momeim, M.F.; El-Shafey, H.A.; El-Deeb, S.T. Biological control of late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. 1981, 59, 253–260. [Google Scholar]
- Payak, M.M.; Lal, S.; Lilaramani, J.; Renfro, B.L. Cephalosporium maydis—A new threat to maize in India. Indian Phytopathol. 1970, 23, 562–569. [Google Scholar]
- Molinero-Ruiz, M.L.; Melero-Vara, J.M.; Mateos, A. Cephalosporium maydis, the cause of late wilt in maize, a pathogen new to Portugal and Spain. Plant Dis. 2011, 94, 379. [Google Scholar] [CrossRef] [PubMed]
- Drori, R.; Sharon, A.; Goldberg, D.; Rabinovitz, O.; Levy, M.; Degani, O. Molecular diagnosis for Harpophora maydis, the cause of maize late wilt in Israel. Phytopathol. Mediterr. 2013, 52, 16–29. [Google Scholar]
- Sabet, K.A.; Samra, A.S.; Mansour, I.M. Saprophytic behaviour of Cephalosporium maydis and C. acremonium. Ann. Appl. Biol. 1970, 66, 265–271. [Google Scholar] [CrossRef]
- Michail, S.H.; Abou-Elseoud, M.S.; Nour Eldin, M.S. Seed health testing of corn for Cephalosporium maydis. Acta Phytopathol. Entomol. Hung. 1999, 34, 35–42. [Google Scholar]
- Sahab, A.F.; Osman, A.R.; Soleman, N.K.; Mikhail, M.S. Studies on root-rot of lupin in Egypt and its control. Egypt. J. Phytopathol. 1985, 17, 23–35. [Google Scholar]
- Degani, O.; Dor, S.; Abraham, D.; Cohen, R. Interactions between Magnaporthiopsis maydis and Macrophomina phaseolina, the Causes of Wilt Diseases in Maize and Cotton. Microorganisms 2020, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Dor, S.; Degani, O. Uncovering the Host Range for Maize Pathogen Magnaporthiopsis maydis. Plants 2019, 8, 259. [Google Scholar] [CrossRef] [PubMed]
- Gazala, P.; Gangappa, E.; Ramesh, S.; Swamy, D. Comparative Breeding potential of two crosses for response to late wilt disease (LWD) in maize (Zea mays L.). Genet. Resour. Crop Evol. 2021, 68, 1751–1757. [Google Scholar] [CrossRef]
- García-Carneros, A.; Girón, I.; Molinero-Ruiz, L. Aggressiveness of Cephalosporium maydis causing late wilt of maize in Spain. Commun. Agric. Appl. Biol. Sci. 2011, 77, 173–179. [Google Scholar]
- Zeller, K.A.; Ismael, A.M.; El-Assiuty, E.M.; Fahmy, Z.M.; Bekheet, F.M.; Leslie, J.F. Relative Competitiveness and Virulence of Four Clonal Lineages of Cephalosporium maydis from Egypt Toward Greenhouse-Grown Maize. Plant Dis. 2002, 86, 373–378. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S.; Chen, A.; Orlov-Levin, V.; Stolov-Yosef, A.; Regev, D.; Rabinovitz, O. Molecular Tracking and Remote Sensing to Evaluate New Chemical Treatments Against the Maize Late Wilt Disease Causal Agent, Magnaporthiopsis maydis. J. Fungi 2020, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Dor, S.; Movshowitz, D.; Fraidman, E.; Rabinovitz, O.; Graph, S. Effective chemical protection against the maize late wilt causal agent, Harpophora maydis, in the field. PLoS ONE 2018, 13, e0208353. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Movshowitz, D.; Dor, S.; Meerson, A.; Goldblat, Y.; Rabinovitz, O. Evaluating Azoxystrobin Seed Coating Against Maize Late Wilt Disease Using a Sensitive qPCR-Based Method. Plant Dis. 2019, 103, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Weinberg, T.; Graph, S. Chemical control of maize late wilt in the field. Phytoparasitica 2014, 42, 559–570. [Google Scholar] [CrossRef]
- Fernández-Ortuño, D.; Torés, J.A.; De Vicente, A.; Pérez-García, A. Mechanisms of resistance to QoI fungicides in phytopathogenic fungi. Int. Microbiol. 2010, 11, 1–9. [Google Scholar]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Ghazy, N.; El-Nahrawy, S. Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 2020, 203, 1195–1209. [Google Scholar] [CrossRef] [PubMed]
- Elshahawy, I.E.; El-Sayed, A.E.-K.B. Maximizing the efficacy of Trichoderma to control Cephalosporium maydis, causing maize late wilt disease, using freshwater microalgae extracts. Egypt. J. Biol. Pest Control 2018, 28, 48. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Gal-Hemed, I.; Atanasova, L.; Komon-Zelazowska, M.; Druzhinina, I.S.; Viterbo, A.; Yarden, O. Marine isolates of Trichoderma spp. as potential halotolerant agents of biological control for arid-zone agriculture. Appl. Environ. Microbiol. 2011, 77, 5100–5109. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Danielle, R.; Dor, S. The microflora of maize grains as a biological barrier against the late wilt causal agent, Magnaporthiopsis maydis. Agronomy 2021, 11, 965. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S. Trichoderma Biological Control to Protect Sensitive Maize Hybrids against Late Wilt Disease in the Field. J. Fungi 2021, 7, 315. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S.; Movshovitz, D.; Rabinovitz, O. Methods for Studying Magnaporthiopsis maydis, the Maize Late Wilt Causal Agent. Agronomy 2019, 9, 181. [Google Scholar] [CrossRef]
- Degani, O.; Goldblat, Y. Ambient Stresses Regulate the Development of the Maize Late Wilt Causing Agent, Harpophora maydis. Agric. Sci. 2014, 5, 571–582. [Google Scholar] [CrossRef]
- Samra, A.S.; Sabet, K.A.; Hingorani, M.K. Late wilt disease of maize caused by Cephalosporium maydis. Phytopathology 1963, 53, 402–406. [Google Scholar]
- Degani, O.; Cernica, G. Diagnosis and Control of Harpophora maydis, the Cause of Late Wilt in Maize. Adv. Microbiol. 2014, 4, 94–105. [Google Scholar] [CrossRef]
- Samuels, G.J.; Ismaiel, A.; Bon, M.-C.; De Respinis, S.; Petrini, O. Trichoderma asperellum sensu lato consists of two cryptic species. Mycologia 2010, 102, 944–966. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Bustos, C.M.; Testi, L.; García-Carneros, A.B.; Molinero-Ruiz, L. Geographic distribution and aggressiveness of Harpophora maydis in the Iberian Peninsula, and thermal detection of maize late wilt. Eur. J. Plant Pathol. 2015, 144, 383–397. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.; Leslie, J.F. Cephalosporium maydis is a distinct species in the Gaeumannomyces-Harpophora species complex. Mycologia 2004, 96, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Zeller, K.A.; Jurgenson, J.E.; El-Assiuty, E.M.; Leslie, J.F. Isozyme and amplified fragment length polymorphisms from Cephalosporium maydis in Egypt. Phytoparasitica 2000, 28, 121–130. [Google Scholar] [CrossRef]
- Weller, S.; Elphinstone, J.; Smith, N.; Boonham, N.; Stead, D. Detection of Ralstonia solanacearumstrains with a quantitative, multiplex, Real-Time, fluorogenic PCR (TaqMan) assay. Appl. Environ. Microbiol. 2000, 66, 2853–2858. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N., Jr. Statistical analysis of Real-Time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hartung, J.S.; Levy, L. Quantitative Real-Time PCR for detection and identification of Candidatus liberibacter species associated with citrus huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef]
- Ortiz-Bustos, C.; López-Bernal, A.; Testi, L.; Molinero-Ruiz, L.J.P.P. Environmental and irrigation conditions can mask the effect of Magnaporthiopsis maydis on growth and productivity of maize. Plant Pathol. 2019, 68, 1555–1564. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Pratap, A.; Kumar, J. Alien Gene Transfer in Crop. Plants, Volume 2: Achievements and Impacts; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2014; Volume 2. [Google Scholar] [CrossRef]
- El-Shafey, H.A.; El-Shorbagy, F.A.; Khalil, I.I.; El-Assiuty, E.M. Additional sources of resistance to the late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. Egypt 1988, 66, 221–230. [Google Scholar]
- Singh, S.D.; Siradhana, B.S. Date of sowing in relation to late wilt disease of maize. Indian Phytopathol. 1988, 41, 489–491. [Google Scholar]
- Johal, L.; Huber, D.M.; Martyn, R. Late wilt of corn (maize) pathway analysis: Intentional introduction of Cephalosporium maydis. In Pathways Analysis for the Introduction to the U.S. of Plant Pathogens of Economic Importance; United States Department of Agriculture, Animal and Plant Health Inspection Service: Washington, DC, USA, 2004. [Google Scholar]
- Abd El-Rahim, M.F.; Fahmy, G.M.; Fahmy, Z.M. Alterations in transpiration and stem vascular tissues of two maize cultivars under conditions of water stress and late wilt disease. Plant Pathol. 2007, 47, 216–223. [Google Scholar] [CrossRef]
- Samra, A.S.; Sabet, K.A.; Abdel-Rahim, M.F. Effect of Soil Conditions and Cultural Practices on Infection with Stalk Rots; U.A.R. Ministry of Agriculture, Government Printing Offices: Cairo, Egypt, 1966; pp. 117–164.
- Chen, J.; Vallikkannu, M.; Karuppiah, V. Systemically Induced Resistance against Maize Diseases by Trichoderma spp. In Trichoderma; Springer: Singapore, 2020; pp. 111–123. [Google Scholar] [CrossRef]
- Veenstra, A.; Rafudeen, M.S.; Murray, S.L. Trichoderma asperellum isolated from African maize seed directly inhibits Fusarium verticillioides growth in vitro. Eur. J. Plant Pathol. 2019, 153, 279–283. [Google Scholar] [CrossRef]
- He, A.-L.; Liu, J.; Wang, X.-H.; Zhang, Q.-G.; Song, W.; Chen, J. Soil application of Trichoderma asperellum GDFS1009 granules promotes growth and resistance to Fusarium graminearum in maize. J. Integr. Agric. 2019, 18, 599–606. [Google Scholar] [CrossRef]
- Prasetyo, J.; Ginting, C.; Akin, H.M.; Suharjo, R.; Niswati, A.; Afandi, A.; Adiwijaya, R.; Sudiono, S.; Nurdin, M. The effect of biological agent and botanical fungicides on maize downy mildew. Biodivers. J. Biol. Divers. 2021, 22. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial biopesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74–81. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of biopesticides and their mode of action against insect pests. Environ. Sustain. 2015, 49–63. [Google Scholar] [CrossRef]
- Chanthini, K.M.-P.; Senthil-Nathan, S.; Soranam, R.; Thanigaivel, A.; Karthi, S.; Sreenath Kumar, C.; Kingsley, S.J.; Kanagaraj Murali-Baskaran, R. Bacterial compounds, as biocontrol agent against early blight (Alternaria solani) and tobacco cut worm (Spodoptera litura Fab.) of tomato (Lycopersicon esculentum Mill.). Arch. Phytopathol. Plant Prot. 2018, 51, 729–753. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolate Designation | Origin | Isolation Location in Northern Israel | Reference | Tested Here in the Field |
|---|---|---|---|---|---|
| Magnaporthiopsis maydis | Hm5 | Zea mays, 618 cv. | Sasa field 2 | [28] | 2019 |
| Magnaporthiopsis maydis | Hm21 | Zea mays, Prelude cv. | Amir field 2 | This study | 2019 |
| Magnaporthiopsis maydis | Hm7 | Zea mays, Colossus cv. | Dovrat field 3 | [28] | 2019, 2020 |
| Magnaporthiopsis maydis | Hm2 | Zea mays, Jubilee cv., CBS 133165 | Sede Nehemia field 2 | [6,28] | 2020 |
| Magnaporthiopsis maydis | Hm29 | Zea mays, 32D99 cv. | Malcia field 2 | This study | 2020 |
| Magnaporthiopsis maydis | Hm30 | Zea mays, 32D99 cv. | Malcia field 2 | This study | 2020 |
| Trichoderma asperelloides | T203 | ATCC 36042, CBS 396.92 | [27,32] | 2019 | |
| Trichoderma longibrachiatum | T7407 | Psammocinia sp. 1 | [25,27] | 2019, 2020 | |
| Trichoderma asperellum | P1 | Zea mays (Prelude cv.) | [26] | 2020 |
| Parameters | 2019 | 2020 |
|---|---|---|
| Dates | 6 August–30 October | 10 September–1 December |
| Temperature (°C) | 25.8 ± 5.7 | 23.0 ± 7.1 |
| Humidity (%) | 63.2 ± 19.2 | 64.0 ± 23.4 |
| Soil temp. top 5 cm (°C) | 30.7 ± 11.2 | 25.6 ± 7.6 |
| Radiation (W/m2) | 235.3 | 172.2 |
| Precipitation (mm) | 53.3 | 140.6 |
| Evaporation (mm) | 556.58 | 299.2 |
| Pairs | Primer | Sequence | Uses | Amplification | References |
|---|---|---|---|---|---|
| Pair 2 | A200a-for A200a-rev | 5′-CCGACGCCTAAAATACAGGA-3′ 5′-GGGCTTTTTAGGGCCTTTTT-3′ | qPCR target gene | 200 bp M. maydis species-specific fragment | [6] |
| Pair 3 | COX-F COX-R | 5′-GTATGCCACGTCGCATTCCAGA-3′ 5′-CAACTACGGATATATAAGRRCCRR AACTG-3′ | qPCR control | Cytochrome c oxidase (COX) gene product | [38,40] |
| Growth Parameter | Control − | Control + | T203 + | T7407 + | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | S.E. | Mean | S.E. | Mean | S.E. | Mean | S.E. | |
| Emergence (plants/m2)) 7 DAS | 16.1 A | 0.19 | 16.4 A | 0.40 | 15.7 B | 0.65 | 16.0 A,B | 0.26 |
| Root weight (g) | 43.5 | 4.5 | 38.7 | 3.9 | 34.4 | 3.7 | 42.4 | 3.9 |
| Shoot weight (g) | 240.9 | 12.2 | 217.5 | 16.2 | 212.8 | 9.5 | 242.7 | 16.8 |
| Shoot height (cm) | 169.0 | 2.9 | 168.2 | 4.7 | 168.8 | 4.2 | 177.6 | 3.1 |
| Cob wet weight (g) | 245.8 | 17.1 | 261.2 | 19.5 | 242.8 | 31.5 | 210.5 | 25.2 |
| Health index (1–4 | 1.2 | 0.2 | 1.2 | 0.2 | 1.3 | 0.2 | 1.5 | 0.2 |
| qPCR (Mm/Cox ratio) | 7.3 × 10−4 | 1.5 × 10−3 | 5.3 × 10−4 | 1.1 × 10−3 | 1.9 × 10−3 | 3.0 × 10−3 | 1.6 × 10−3 | 1.9 × 10−3 |
| Treatment | SI 2 | Wet Weight (g) | Number of Leaves | Plant Height (cm) | qPCR (Mm/Cox) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | S.E. | Mean | S.E. | Mean | S.E. | Mean | S.E. | ||
| Control- | − | 255.7 | 10.6 | 9.87 | 0.2 | 115.4 | 2.1 | 5.5 × 10−4 | 3.3 × 10−4 |
| Control- | Stabbing | 240.2 | 17.1 | 9.80 | 0.3 | 111.7 | 4.1 | 3.1 × 10−3 | 2.4 × 10−3 |
| Control+ | − | 240.5 | 11.0 | 9.79 | 0.2 | 113.7 | 2.3 | 2.5 × 10−4 | 1.8 × 10−4 |
| Control+ | + | 243.3 | 16.0 | 9.75 | 0.2 | 112.2 | 3.0 | 7.2 × 10−4 | 6.6 × 10−4 |
| P1 | − | 245.1 | 13.6 | 9.00 | 0.2 | 116.9 | 2.5 | 3.0 × 10−4 | 1.9 × 10−4 |
| P1 | + | 248.8 | 16.3 | 10.50 | 0.2 | 118.7 | 3.6 | 5.1 × 10−3 | 2.8 × 10−3 |
| T7407 | − | 258.6 | 15.3 | 9.90 | 0.2 | 119.9 | 2.3 | 3.1 × 10−4 | 2.1 × 10−4 |
| T7407 | + | 246.9 | 9.7 | 10.08 | 0.2 | 118.6 | 2.3 | 1.3 × 10−2 | 1.0 × 10−2 |
| TREATMENT | SI 2 | YIELD (KG/M2) | A-CLASS (GR’) | B-CLASS (GR’) | |||
|---|---|---|---|---|---|---|---|
| Mean | S.E. | Mean | S.E. | Mean | S.E. | ||
| CONTROL- | − | 1.20 | 0.08 | 369.2 | 14.2 | 160.3 | 17.3 |
| CONTROL- | Stabbing | 1.11 | 0.15 | 345.9 | 12.9 | 158.3 | 5.5 |
| CONTROL+ | − | 1.14 | 0.08 | 341.9 | 20.3 | 152.4 | 14.9 |
| CONTROL+ | + | 1.19 | 0.08 | 347.8 | 17.2 | 195.8 | 33.1 |
| P1 | − | 1.09 | 0.05 | 333.1 | 20.9 | 180.2 | 16.1 |
| P1 | + | 1.22 | 0.04 | 363.3 | 25.6 | 187.3 | 16.9 |
| T7407 | − | 1.16 | 0.17 | 365.5 | 43.8 | 167.8 | 24.4 |
| T7407 | + | 1.20 | 0.05 | 354.5 | 13.9 | 156.9 | 19.2 |
| Location and Year | Dates | Average Temp. | Precipitation | Dehydration | M. maydis DNA | Reference |
|---|---|---|---|---|---|---|
| Amir 2016 (spring–summer) | 25 May–2 August (75 DAS) | 27 °C | 0.6 mm | 60% (69 DAS) 100% (75 DAS) | 0.05 (60 DAS) 7.8 × 10−05 (75 DAS) | [17] |
| Amir 2017 (spring–summer) | 24 May–2 August (70 DAS) | 27 °C | 0 mm | 73% | 6.26 | [16] |
| Neot Mordechai 2018 (spring–Summer) | 23 April–5 July (73 DAS) | 25 °C | 30 mm | Less than 10% | 6.5 × 10−05 | [15] |
| Amir 2018 (summer) | 21 June–5 September (71 DAS) | 28 °C | 3 mm | 72% | 0.02 | [15] |
| Gadash farm 2019 (summer) | 6 August–30 October (85 DAS) | 26 °C | 53 mm | 30% | 5.3 × 10−04 | This study |
| Gadash farm 2020 (autumn) | 10 September–1 December (82 DAS) | 23 °C | 141 mm | Less than 10% | 4.5 × 10−04 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum Confer Growth Promotion and Protection against Late Wilt Disease in the Field. J. Fungi 2021, 7, 444. https://doi.org/10.3390/jof7060444
Degani O, Rabinovitz O, Becher P, Gordani A, Chen A. Trichoderma longibrachiatum and Trichoderma asperellum Confer Growth Promotion and Protection against Late Wilt Disease in the Field. Journal of Fungi. 2021; 7(6):444. https://doi.org/10.3390/jof7060444
Chicago/Turabian StyleDegani, Ofir, Onn Rabinovitz, Paz Becher, Asaf Gordani, and Assaf Chen. 2021. "Trichoderma longibrachiatum and Trichoderma asperellum Confer Growth Promotion and Protection against Late Wilt Disease in the Field" Journal of Fungi 7, no. 6: 444. https://doi.org/10.3390/jof7060444
APA StyleDegani, O., Rabinovitz, O., Becher, P., Gordani, A., & Chen, A. (2021). Trichoderma longibrachiatum and Trichoderma asperellum Confer Growth Promotion and Protection against Late Wilt Disease in the Field. Journal of Fungi, 7(6), 444. https://doi.org/10.3390/jof7060444