
Evidence of a New MoYpd1p Phosphotransferase Isoform in the Multistep Phosphorelay System of Magnaporthe oryzae


Abstract
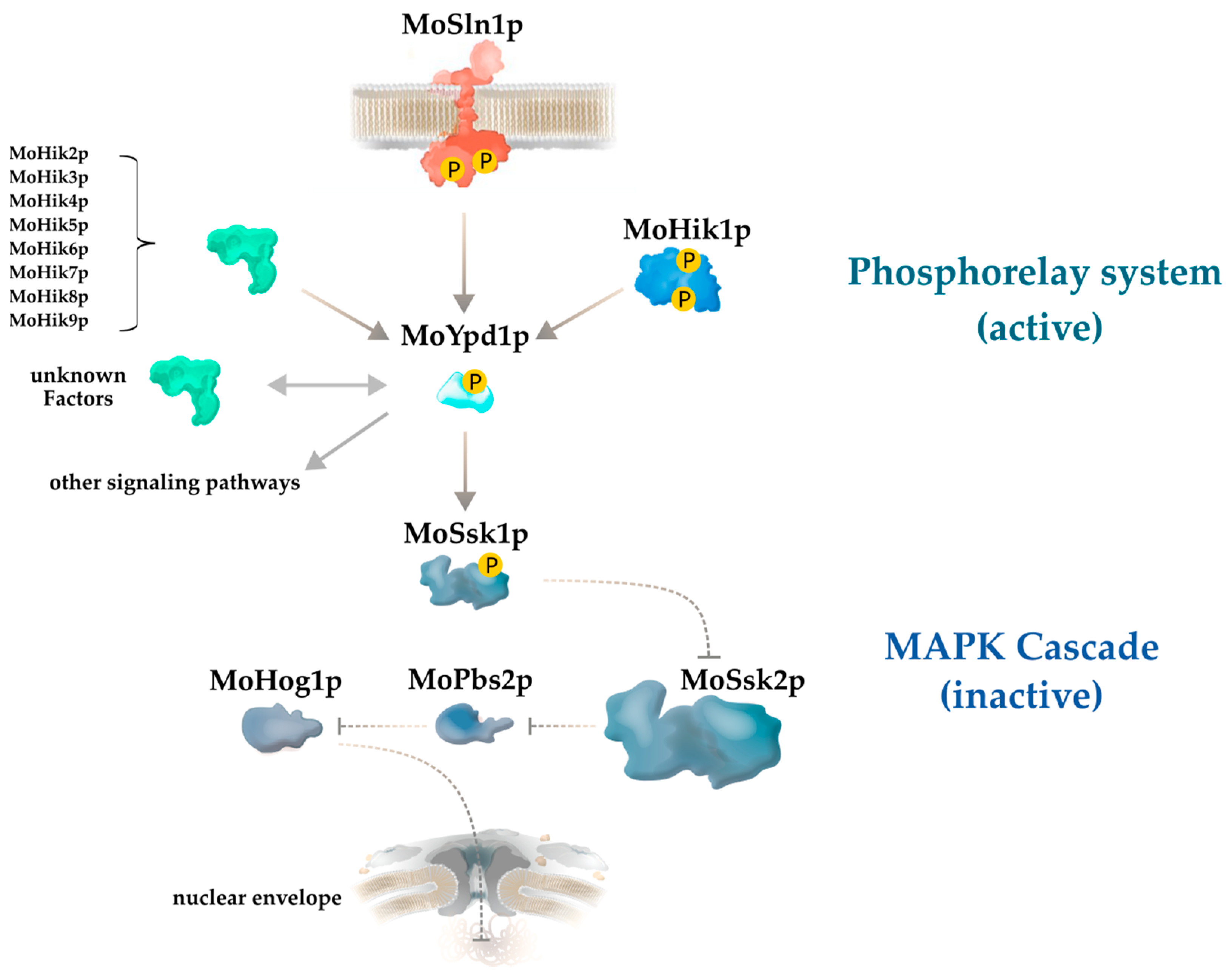
1. Introduction
2. Materials and Methods
2.1. Strains, Culture/Growth Conditions
2.2. RNA Extraction and Quality Control, cDNA Synthesis
2.3. PCR of MoYPD1 Isoforms on cDNA Level
2.4. DNA Manipulations, Construction of the Expression Vector, and Fungal Transformation
2.5. Protein Expression and Purification
2.6. In-Gel Tryptic Digest
2.7. LC-MS/MS Peptide Identification
2.8. Data Processing
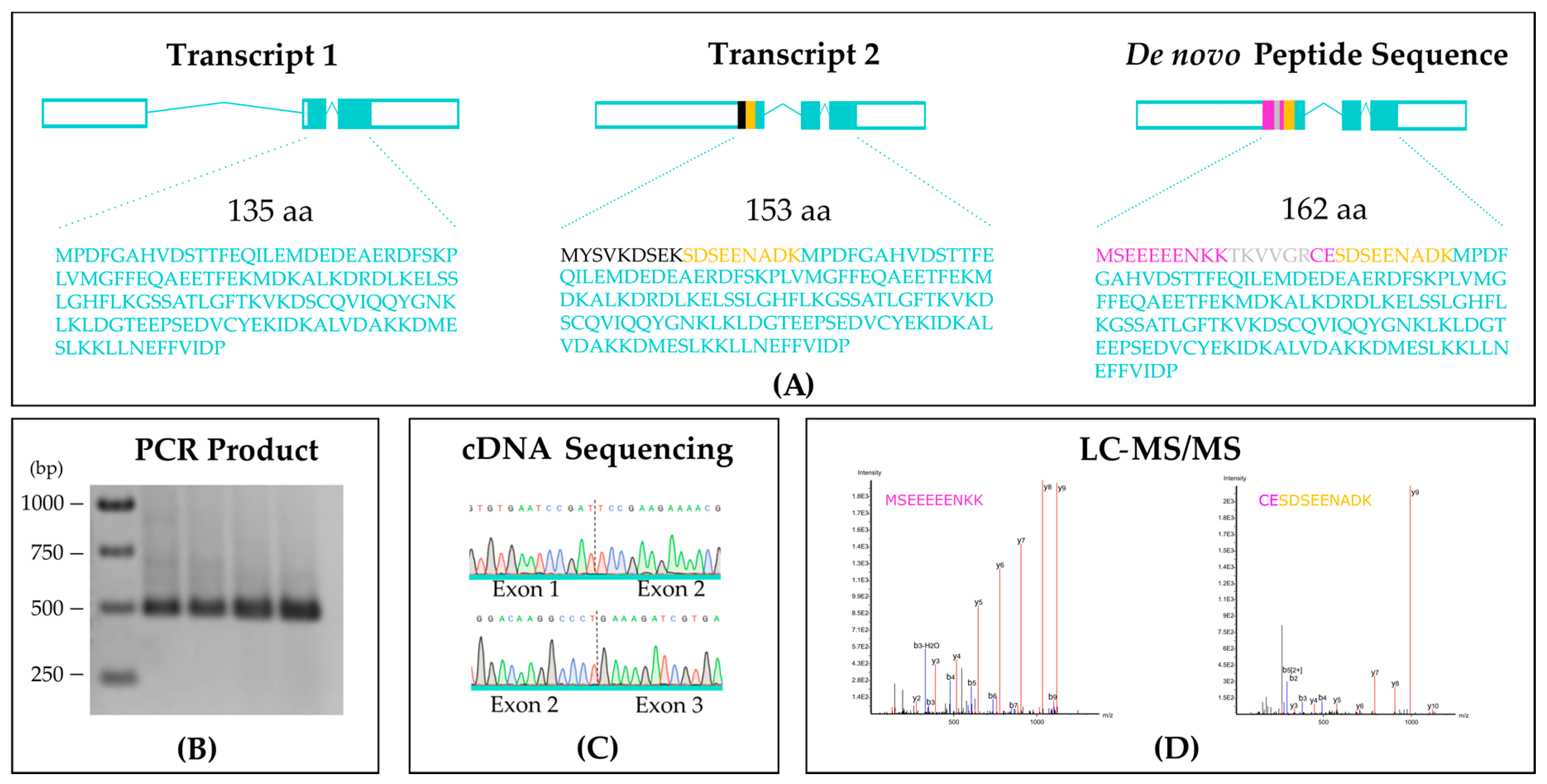
3. Results
A Novel MoYpd1p Isoform Enables Signal Diversity in the Multistep Phosphorelay System
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuo, C.-Y.; Chen, S.-A.; Hsueh, Y.-P. The high osmolarity glycerol (HOG) pathway functions in osmosensing, trap morphogenesis and conidiation of the nematode-trapping fungus Arthrobotrys oligospora. J. Fungi 2020, 6, 191. [Google Scholar] [CrossRef] [PubMed]
- De Nadal, E.; Alepuz, P.M.; Posas, F. Dealing with osmostress through MAP kinase activation. EMBO Rep. 2002, 3, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, H.; He, X.; Zhang, S.; Wang, J.; Wang, L.; Guo, D.; Guo, X. Scaffold protein GhMORG1 enhances the resistance of cotton to Fusarium oxysporum by facilitating the MKK6-MPK4 cascade. Plant. Biotechnol. J. 2020, 18, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Jogawat, A.; Vadassery, J.; Verma, N.; Oelmüller, R.; Dua, M.; Nevo, E.; Johri, A.K. PiHOG1, a stress regulator MAP kinase from the root endophyte fungus Piriformospora indica, confers salinity stress tolerance in rice plants. Sci. Rep. 2016, 6, 36765. [Google Scholar] [CrossRef]
- Foster, A.J.; Ryder, L.S.; Kershaw, M.J.; Talbot, N.J. The role of glycerol in the pathogenic lifestyle of the rice blast fungus Magnaporthe oryzae. Environ. Microbiol. 2017, 19, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef]
- Hérivaux, A.; So, Y.-S.; Gastebois, A.; Latgé, J.-P.; Bouchara, J.-P.; Bahn, Y.-S.; Papon, N. Major sensing proteins in pathogenic fungi: The hybrid histidine kinase family. PLoS Pathog. 2016, 12, e1005683. [Google Scholar] [CrossRef]
- Jacob, S.; Foster, A.J.; Yemelin, A.; Thines, E. High osmolarity glycerol (HOG) signalling in Magnaporthe oryzae: Identification of MoYPD1 and its role in osmoregulation, fungicide action, and pathogenicity. Fungal. Biol. 2015, 119, 580–594. [Google Scholar] [CrossRef]
- Joubert, A.; Bataille-Simoneau, N.; Campion, C.; Guillemette, T.; Hudhomme, P.; Iacomi-Vasilescu, B.; Leroy, T.; Pochon, S.; Poupard, P. Cell wall integrity and high osmolarity glycerol pathways are required for adaptation of Alternaria brassicicola to cell wall stress caused by brassicaceous indolic phytoalexins. Cell Microbiol. 2011, 13, 62–80. [Google Scholar] [CrossRef] [PubMed]
- De Assis, L.J.; Silva, L.P.; Liu, L.; Schmitt, K.; Valerius, O.; Braus, G.H.; Ries, L.N.A.; Goldman, G.H. The high osmolarity glycerol mitogen-activated protein kinase regulates glucose catabolite repression in filamentous fungi. PLoS Genet. 2020, 16, e1008996. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yin, D.; Yin, Y.; Cao, Y.; Ma, Z. The response regulator BcSkn7 is required for vegetative differentiation and adaptation to oxidative and osmotic stresses in Botrytis cinerea. Mol. Plant. Pathol. 2015, 16, 276–287. [Google Scholar] [CrossRef]
- Tanaka, C.; Izumitsu, K. Two-component signaling system in filamentous fungi and the mode of action of dicarboximide and phenylpyrrole fungicides. In Fungicides; Carisse, O., Ed.; IntechOpen: London, UK, 2010; Available online: https://books.google.de/books?id=NuaODwAAQBAJ&pg=PA523&lpg=PA523&dq=Two-component+signaling+system+in+filamentous+fungi+and+the+mode+of+action+of+dicarboximide+and+phenylpyrrole+fungicides&source=bl&ots=aUwbFN0NQQ&sig=ACfU3U2Su_ChjXGLcXBc9YJzFAWP2giX8g&hl=en&sa=X&ved=2ahUKEwi34o6znOzvAhWTHuwKHR5HAyMQ6AEwBHoECAYQAw#v=onepage&q=Tanaka&f=false (accessed on 7 April 2021).
- Kempken, F. Alternative splicing in ascomycetes. Appl. Microbiol. Biotechnol. 2013, 97, 4235–4241. [Google Scholar] [CrossRef]
- Stamm, S.; Ben-Ari, S.; Rafalska, I.; Tang, Y.; Zhang, Z.; Toiber, D.; Thanaraj, T.A.; Soreq, H. Function of alternative splicing. Gene 2004, 344, 1–20. [Google Scholar] [CrossRef]
- Schmucker, D.; Clemens, J.C.; Shu, H.; Worby, C.A.; Xiao, J.; Muda, M.; Dixon, J.E.; Zipursky, S.L. Drosophila Dscam is an axon guidance receptor exhibiting extraordinary molecular diversity. Cell 2000, 101, 671–684. [Google Scholar] [CrossRef]
- Li, Z.; Wu, L.; Wu, H.; Zhang, X.; Mei, J.; Zhou, X.; Wang, G.-L.; Liu, W. Arginine methylation is required for remodelling pre-mRNA splicing and induction of autophagy in rice blast fungus. New Phytol. 2020, 225, 413–429. [Google Scholar] [CrossRef]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef]
- Liu, X.Y.; Fan, L.; Gao, J.; Shen, X.Y.; Hou, C.L. Global identification of alternative splicing in Shiraia bambusicola and analysis of its regulation in hypocrellin biosynthesis. Appl. Microbiol. Biotechnol. 2020, 104, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, G.; Yu, D.; Huang, W.; Cheng, C.; Liao, S.; Wu, Q.; Zhang, Y. Transcriptome analysis reveals the complexity of alternative splicing regulation in the fungus Verticillium dahliae. BMC Genom. 2017, 18, 130. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.-B.; Li, D.; Shi, W.-L.; Qin, Q.-L.; Wang, X.-W.; Rong, J.-C.; Sun, C.-Y.; Huang, F.; Zhang, X.-Y.; Dong, X.-W.; et al. Deep RNA sequencing reveals a high frequency of alternative splicing events in the fungus Trichoderma longibrachiatum. BMC Genom. 2015, 16, 54. [Google Scholar] [CrossRef]
- Odenbach, D.; Breth, B.; Thines, E.; Weber, R.W.S.; Anke, H.; Foster, A.J. The transcription factor Con7p is a central regulator of infection-related morphogenesis in the rice blast fungus Magnaporthe grisea. Mol. Microbiol. 2007, 64, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Bohnert, S.; Neumann, H.; Thines, E.; Jacob, S. Visualizing fungicide action: An in vivo tool for rapid validation of fungicides with target location HOG pathway. Pest. Manag. Sci. 2019, 75, 772–778. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Thines, E. Multistep phosphorelay in fungi: The enigma of multiple signals and a limited number of signaling pathways. Mycol. Prog. 2017, 16, 1007–1013. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Qiu, K.; He, R.; Feng, X.; Liang, X. The occurrence and function of alternative splicing in fungi. Fungal. Biol. Rev. 2020, 34, 178–188. [Google Scholar] [CrossRef]
- Gao, X.; Yin, C.; Liu, X.; Peng, J.; Chen, D.; He, D.; Shi, W.; Zhao, W.; Yang, J.; Peng, Y.-L. A glycine-rich protein MoGrp1 functions as a novel splicing factor to regulate fungal virulence and growth in Magnaporthe oryzae. Phytopathol. Res. 2019, 1, 2. [Google Scholar] [CrossRef]
- Sieber, P.; Voigt, K.; Kämmer, P.; Brunke, S.; Schuster, S.; Linde, J. Comparative study on alternative splicing in human fungal pathogens suggests its involvement during host invasion. Front. Microbiol. 2018, 9, 2313. [Google Scholar] [CrossRef]
- Ibrahim, H.M.M.; Kusch, S.; Didelon, M.; Raffaele, S. Genome-wide alternative splicing profiling in the fungal plant pathogen Sclerotinia sclerotiorum during the colonization of diverse host families. Mol. Plant. Pathol. 2021, 22, 31–47. [Google Scholar] [CrossRef]
- Mastrangelo, A.M.; Marone, D.; Laidò, G.; De Leonardis, A.M.; De Vita, P. Alternative splicing: Enhancing ability to cope with stress via transcriptome plasticity. Plant Sci. 2012, 185–186, 40–49. [Google Scholar] [CrossRef]
- Mavrianos, J.; Desai, C.; Chauhan, N. Two-component histidine phosphotransfer protein Ypd1 is not essential for viability in Candida albicans. Eukaryot Cell 2014, 13, 452–460. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Name | Sequence (5′→ 3′) |
|---|---|
| SJ-632 (EF1prom-for) | ttactgatcactgattaagtCTGAGAGCGAGAAAAAAAAACTCTTC |
| SJ-633 (EF1prom-rev) | tgaggatgactccacatGGTGGCGGTTTGGTGCTC |
| SJ-634 (GFP-for) | aaccgccaccatgtggagtcatcctcaattcgagaaaggtggaggttctggcggtggatcgggaggttcagcgtggagccacccgcagttcgaaaaagatctGTGAGCAAGGGCGAGGAG |
| SJ-635 (GFP-rev) | gccgggcggccgctttacttTTACTTGTACAGCTCGTCCATG |
| SJ-651(Ypd1-twinstrep-for) | gaggttcagcgtggagccacccgcagttcgaaaaaATGTCGGAGGAGGAGGAGGAGAACA |
| SJ-652(Ypd1-twinstrep-rev) | cgatctgcagccgggcggccgctttacttttacttCTAAGGATCGATCACGAAAAACTCATTG |
| SJ-955 (Ypd1-var3-rev) | CTAAGGATCGATCACGAAAAAC |
| SJ-956 (Ypd1-var4-for) | ATGTCGGAGGAGGAGGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bühring, S.; Yemelin, A.; Michna, T.; Tenzer, S.; Jacob, S. Evidence of a New MoYpd1p Phosphotransferase Isoform in the Multistep Phosphorelay System of Magnaporthe oryzae. J. Fungi 2021, 7, 389. https://doi.org/10.3390/jof7050389
Bühring S, Yemelin A, Michna T, Tenzer S, Jacob S. Evidence of a New MoYpd1p Phosphotransferase Isoform in the Multistep Phosphorelay System of Magnaporthe oryzae. Journal of Fungi. 2021; 7(5):389. https://doi.org/10.3390/jof7050389
Chicago/Turabian StyleBühring, Sri, Alexander Yemelin, Thomas Michna, Stefan Tenzer, and Stefan Jacob. 2021. "Evidence of a New MoYpd1p Phosphotransferase Isoform in the Multistep Phosphorelay System of Magnaporthe oryzae" Journal of Fungi 7, no. 5: 389. https://doi.org/10.3390/jof7050389
APA StyleBühring, S., Yemelin, A., Michna, T., Tenzer, S., & Jacob, S. (2021). Evidence of a New MoYpd1p Phosphotransferase Isoform in the Multistep Phosphorelay System of Magnaporthe oryzae. Journal of Fungi, 7(5), 389. https://doi.org/10.3390/jof7050389