
Molecular Epidemiology of Aspergillus fumigatus in Chronic Pulmonary Aspergillosis Patients

 ,
, 
 , and
, and
Abstract
1. Introduction
2. Epidemiology of Aspergillus fumigatus
3. Aspergillus fumigatus Genotyping Methods
4. Aspergillus fumigatus Genotyping Study Results
4.1. Distinguishing the Nature of Infection
4.2. Genomic Dynamics during Infection
4.3. Routes of Transmission
4.4. Geographical Distribution of the Genetic Diversity of Aspergillus fumigatus
5. Genetic Diversity and Geographical Distribution of Triazole-Resistant A. fumigatus Strains
6. Genotyping of Aspergillus fumigatus Isolates from the UK National Aspergillosis Centre
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kosmidis, C.; Denning, D.W. The clinical spectrum of pulmonary aspergillosis. Thorax 2015, 70, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Maghrabi, F.; Denning, D.W. The Management of Chronic Pulmonary Aspergillosis: The UK National Aspergillosis Centre Approach. Curr. Fungal Infect. Rep. 2017, 11, 242–251. [Google Scholar] [CrossRef]
- Denning, D.W.; Cadranel, J.; Beigelman-Aubry, C.; Ader, F.; Chakrabarti, A.; Blot, S.; Ullmann, A.J.; Dimopoulos, G.; Lange, C. Chronic pulmonary aspergillosis: Rationale and clinical guidelines for diagnosis and management. Eur. Respir. J. 2016, 47, 45–68. [Google Scholar] [CrossRef]
- Lestrade, P.P.A.; Meis, J.F.; Melchers, W.J.G.; Verweij, P.E. Triazole resistance in Aspergillus fumigatus: Recent insights and challenges for patient management. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2019, 25, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Riley, L.W.; Blanton, R.E. Advances in Molecular Epidemiology of Infectious Diseases: Definitions, Approaches, and Scope of the Field. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- King, P.; Pham, L.K.; Waltz, S.; Sphar, D.; Yamamoto, R.T.; Conrad, D.; Taplitz, R.; Torriani, F.; Forsyth, R.A. Longitudinal Metagenomic Analysis of Hospital Air Identifies Clinically Relevant Microbes. PLoS ONE 2016, 11, e0160124. [Google Scholar] [CrossRef]
- Van der Linden, J.W.; Camps, S.M.; Kampinga, G.A.; Arends, J.P.; Debets-Ossenkopp, Y.J.; Haas, P.J.; Rijnders, B.J.; Kuijper, E.J.; van Tiel, F.H.; Varga, J.; et al. Aspergillosis due to voriconazole highly resistant Aspergillus fumigatus and recovery of genetically related resistant isolates from domiciles. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 57, 513–520. [Google Scholar] [CrossRef]
- Chen, Y.C.; Kuo, S.F.; Wang, H.C.; Wu, C.J.; Lin, Y.S.; Li, W.S.; Lee, C.H. Azole resistance in Aspergillus species in Southern Taiwan: An epidemiological surveillance study. Mycoses 2019, 62, 1174–1181. [Google Scholar] [CrossRef]
- Dauchy, C.; Bautin, N.; Nseir, S.; Reboux, G.; Wintjens, R.; Le Rouzic, O.; Sendid, B.; Viscogliosi, E.; Le Pape, P.; Arendrup, M.C.; et al. Emergence of Aspergillus fumigatus azole resistance in azole-naïve patients with chronic obstructive pulmonary disease and their homes. Indoor Air 2018, 28, 298–306. [Google Scholar] [CrossRef]
- Godeau, C.; Reboux, G.; Scherer, E.; Laboissiere, A.; Lechenault-Bergerot, C.; Millon, L.; Rocchi, S. Azole-resistant Aspergillus fumigatus in the hospital: Surveillance from flower beds to corridors. Am. J. Infect. Control 2020, 48, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.M.; Cohen, J.; Rees, A.J.; Holden, D.W. Molecular epidemiological study of invasive pulmonary aspergillosis in a renal transplantation unit. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 1994, 13, 318–321. [Google Scholar] [CrossRef]
- Warris, A.; Klaassen, C.H.; Meis, J.F.; De Ruiter, M.T.; De Valk, H.A.; Abrahamsen, T.G.; Gaustad, P.; Verweij, P.E. Molecular epidemiology of Aspergillus fumigatus isolates recovered from water, air, and patients shows two clusters of genetically distinct strains. J. Clin. Microbiol. 2003, 41, 4101–4106. [Google Scholar] [CrossRef]
- Lavergne, R.A.; Chouaki, T.; Hagen, F.; Toublanc, B.; Dupont, H.; Jounieaux, V.; Meis, J.F.; Morio, F.; Le Pape, P. Home Environment as a Source of Life-Threatening Azole-Resistant Aspergillus fumigatus in Immunocompromised Patients. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 64, 76–78. [Google Scholar] [CrossRef]
- Verweij, P.E.; Zhang, J.; Debets, A.J.M.; Meis, J.F.; van de Veerdonk, F.L.; Schoustra, S.E.; Zwaan, B.J.; Melchers, W.J.G. In-host adaptation and acquired triazole resistance in Aspergillus fumigatus: A dilemma for clinical management. Lancet. Infect. Dis. 2016, 16, e251–e260. [Google Scholar] [CrossRef]
- Tekaia, F.; Latgé, J.P. Aspergillus fumigatus: Saprophyte or pathogen? Curr. Opin. Microbiol. 2005, 8, 385–392. [Google Scholar] [CrossRef]
- Hohl, T.M.; Feldmesser, M. Aspergillus fumigatus: Principles of pathogenesis and host defense. Eukaryot. Cell 2007, 6, 1953–1963. [Google Scholar] [CrossRef]
- Sharpe, R.A.; Cocq, K.L.; Nikolaou, V.; Osborne, N.J.; Thornton, C.R. Identifying risk factors for exposure to culturable allergenic moulds in energy efficient homes by using highly specific monoclonal antibodies. Environ. Res. 2016, 144, 32–42. [Google Scholar] [CrossRef]
- Richardson, M.; Rautemaa-Richardson, R. Exposure to Aspergillus in Home and Healthcare Facilities’ Water Environments: Focus on Biofilms. Microorganisms 2019, 7, 7. [Google Scholar] [CrossRef]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of chronic pulmonary aspergillosis as a sequel to pulmonary tuberculosis. Bull. World Health Organ. 2011, 89, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of chronic pulmonary aspergillosis complicating sarcoidosis. Eur. Respir. J. 2013, 41, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Barac, A.; Kosmidis, C.; Alastruey-Izquierdo, A.; Salzer, H.J.F. Chronic pulmonary aspergillosis update: A year in review. Med. Mycol. 2019, 57, S104–s109. [Google Scholar] [CrossRef]
- Van der Linden, J.W.; Arendrup, M.C.; Warris, A.; Lagrou, K.; Pelloux, H.; Hauser, P.M.; Chryssanthou, E.; Mellado, E.; Kidd, S.E.; Tortorano, A.M.; et al. Prospective multicenter international surveillance of azole resistance in Aspergillus fumigatus. Emerg. Infect. Dis. 2015, 21, 1041–1044. [Google Scholar] [CrossRef]
- Mondon, P.; Thélu, J.; Lebeau, B.; Ambroise-Thomas, P.; Grillot, R. Virulence of Aspergillus fumigatus strains investigated by random amplified polymorphic DNA analysis. J. Med. Microbiol. 1995, 42, 299–303. [Google Scholar] [CrossRef][Green Version]
- Mondon, P.; Brenier, M.P.; Symoens, F.; Rodriguez, E.; Coursange, E.; Chaib, F.; Lebeau, B.; Piens, M.A.; Tortorano, A.M.; Mallié, M.; et al. Molecular typing of Aspergillus fumigatus strains by sequence-specific DNA primer (SSDP) analysis. FEMS Immunol. Med. Microbiol. 1997, 17, 95–102. [Google Scholar] [CrossRef]
- Varga, J. Molecular typing of aspergilli: Recent developments and outcomes. Med. Mycol. 2006, 44, S149–S169. [Google Scholar] [CrossRef] [PubMed]
- Vanhee, L.M.; Nelis, H.J.; Coenye, T. What can be learned from genotyping of fungi? Med. Mycol. 2010, 48, S60–S69. [Google Scholar] [CrossRef][Green Version]
- Taylor, J.W.; Geiser, D.M.; Burt, A.; Koufopanou, V. The evolutionary biology and population genetics underlying fungal strain typing. Clin. Microbiol. Rev. 1999, 12, 126–146. [Google Scholar] [CrossRef]
- Balajee, S.A.; Gribskov, J.L.; Hanley, E.; Nickle, D.; Marr, K.A. Aspergillus lentulus sp. nov., a new sibling species of A. fumigatus. Eukaryot. Cell 2005, 4, 625–632. [Google Scholar] [CrossRef] [PubMed]
- De Valk, H.A.; Meis, J.F.; Curfs, I.M.; Muehlethaler, K.; Mouton, J.W.; Klaassen, C.H. Use of a novel panel of nine short tandem repeats for exact and high-resolution fingerprinting of Aspergillus fumigatus isolates. J. Clin. Microbiol. 2005, 43, 4112–4120. [Google Scholar] [CrossRef]
- Klaassen, C.H.; Osherov, N. Aspergillus strain typing in the genomics era. Stud. Mycol. 2007, 59, 47–51. [Google Scholar] [CrossRef]
- Alanio, A.; Desnos-Ollivier, M.; Garcia-Hermoso, D.; Bretagne, S. Investigating Clinical Issues by Genotyping of Medically Important Fungi: Why and How? Clin. Microbiol. Rev. 2017, 30, 671–707. [Google Scholar] [CrossRef]
- Vanhee, L.M.; Symoens, F.; Jacobsen, M.D.; Nelis, H.J.; Coenye, T. Comparison of multiple typing methods for Aspergillus fumigatus. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2009, 15, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.H.; de Valk, H.A.; Balajee, S.A.; Meis, J.F. Utility of CSP typing to sub-type clinical Aspergillus fumigatus isolates and proposal for a new CSP type nomenclature. J. Microbiol. Methods 2009, 77, 292–296. [Google Scholar] [CrossRef]
- Balajee, S.A.; Tay, S.T.; Lasker, B.A.; Hurst, S.F.; Rooney, A.P. Characterization of a novel gene for strain typing reveals substructuring of Aspergillus fumigatus across North America. Eukaryot. Cell 2007, 6, 1392–1399. [Google Scholar] [CrossRef]
- De Ruiter, M.T.; de Valk, H.A.; Meis, J.F.; Klaassen, C.H. Retrotransposon insertion-site context (RISC) typing: A novel typing method for Aspergillus fumigatus and a convenient PCR alternative to restriction fragment length polymorphism analysis. J. Microbiol. Methods 2007, 70, 528–534. [Google Scholar] [CrossRef] [PubMed]
- De Valk, H.A.; Klaassen, C.H.; Meis, J.F. Molecular typing of Aspergillus species. Mycoses 2008, 51, 463–476. [Google Scholar] [CrossRef] [PubMed]
- De Valk, H.A.; Meis, J.F.; Klaassen, C.H. Microsatellite based typing of Aspergillus fumigatus: Strengths, pitfalls and solutions. J. Microbiol. Methods 2007, 69, 268–272. [Google Scholar] [CrossRef]
- De Valk, H.A.; Meis, J.F.; Bretagne, S.; Costa, J.M.; Lasker, B.A.; Balajee, S.A.; Pasqualotto, A.C.; Anderson, M.J.; Alcázar-Fuoli, L.; Mellado, E.; et al. Interlaboratory reproducibility of a microsatellite-based typing assay for Aspergillus fumigatus through the use of allelic ladders: Proof of concept. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2009, 15, 180–187. [Google Scholar] [CrossRef]
- Araujo, R.; Pina-Vaz, C.; Rodrigues, A.G.; Amorim, A.; Gusmão, L. Simple and highly discriminatory microsatellite-based multiplex PCR for Aspergillus fumigatus strain typing. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2009, 15, 260–266. [Google Scholar] [CrossRef][Green Version]
- De Groot, T.; Meis, J.F. Microsatellite Stability in STR Analysis Aspergillus fumigatus Depends on Number of Repeat Units. Front. Cell. Infect. Microbiol. 2019, 9, 82. [Google Scholar] [CrossRef]
- Pringle, A.; Baker, D.M.; Platt, J.L.; Wares, J.P.; Latgé, J.P.; Taylor, J.W. Cryptic speciation in the cosmopolitan and clonal human pathogenic fungus Aspergillus fumigatus. Evol. Int. J. Org. Evol. 2005, 59, 1886–1899. [Google Scholar] [CrossRef]
- Sewell, T.R.; Zhu, J.; Rhodes, J.; Hagen, F.; Meis, J.F.; Fisher, M.C.; Jombart, T. Nonrandom Distribution of Azole Resistance across the Global Population of Aspergillus fumigatus. MBio 2019, 10. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; Gil, H.; Monteiro, M.C.; Pelaez, T.; Mellado, E. A New Aspergillus fumigatus Typing Method Based on Hypervariable Tandem Repeats Located within Exons of Surface Protein Coding Genes (TRESP). PLoS ONE 2016, 11, e0163869. [Google Scholar] [CrossRef] [PubMed]
- Levdansky, E.; Romano, J.; Shadkchan, Y.; Sharon, H.; Verstrepen, K.J.; Fink, G.R.; Osherov, N. Coding tandem repeats generate diversity in Aspergillus fumigatus genes. Eukaryot. Cell 2007, 6, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; Escribano, P.; Gomez, A.; Guinea, J.; Mellado, E. Comparison of Two Highly Discriminatory Typing Methods to Analyze Aspergillus fumigatus Azole Resistance. Front. Microbiol. 2018, 9, 1626. [Google Scholar] [CrossRef]
- Mortensen, K.L.; Jensen, R.H.; Johansen, H.K.; Skov, M.; Pressler, T.; Howard, S.J.; Leatherbarrow, H.; Mellado, E.; Arendrup, M.C. Aspergillus species and other molds in respiratory samples from patients with cystic fibrosis: A laboratory-based study with focus on Aspergillus fumigatus azole resistance. J. Clin. Microbiol. 2011, 49, 2243–2251. [Google Scholar] [CrossRef] [PubMed]
- Albarrag, A.M.; Anderson, M.J.; Howard, S.J.; Robson, G.D.; Warn, P.A.; Sanglard, D.; Denning, D.W. Interrogation of related clinical pan-azole-resistant Aspergillus fumigatus strains: G138C, Y431C, and G434C single nucleotide polymorphisms in cyp51A, upregulation of cyp51A, and integration and activation of transposon Atf1 in the cyp51A promoter. Antimicrob. Agents Chemother. 2011, 55, 5113–5121. [Google Scholar] [CrossRef]
- Howard, S.J.; Pasqualotto, A.C.; Anderson, M.J.; Leatherbarrow, H.; Albarrag, A.M.; Harrison, E.; Gregson, L.; Bowyer, P.; Denning, D.W. Major variations in Aspergillus fumigatus arising within aspergillomas in chronic pulmonary aspergillosis. Mycoses 2013, 56, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Camps, S.M.; Rijs, A.J.; Klaassen, C.H.; Meis, J.F.; O’Gorman, C.M.; Dyer, P.S.; Melchers, W.J.; Verweij, P.E. Molecular epidemiology of Aspergillus fumigatus isolates harboring the TR34/L98H azole resistance mechanism. J. Clin. Microbiol. 2012, 50, 2674–2680. [Google Scholar] [CrossRef]
- Bader, O.; Tünnermann, J.; Dudakova, A.; Tangwattanachuleeporn, M.; Weig, M.; Groß, U. Environmental isolates of azole-resistant Aspergillus fumigatus in Germany. Antimicrob. Agents Chemother. 2015, 59, 4356–4359. [Google Scholar] [CrossRef]
- Tsuchido, Y.; Tanaka, M.; Nakano, S.; Yamamoto, M.; Matsumura, Y.; Nagao, M. Prospective multicenter surveillance of clinically isolated Aspergillus species revealed azole-resistant Aspergillus fumigatus isolates with TR34/L98H mutation in the Kyoto and Shiga regions of Japan. Med. Mycol. 2019, 57, 997–1003. [Google Scholar] [CrossRef]
- Badali, H.; Vaezi, A.; Haghani, I.; Yazdanparast, S.A.; Hedayati, M.T.; Mousavi, B.; Ansari, S.; Hagen, F.; Meis, J.F.; Chowdhary, A. Environmental study of azole-resistant Aspergillus fumigatus with TR34/L98H mutations in the cyp51A gene in Iran. Mycoses 2013, 56, 659–663. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Randhawa, H.S.; Gaur, S.N.; Klaassen, C.H.; Meis, J.F. Isolation of multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR/L98H mutations in the cyp51A gene in India. J. Antimicrob. Chemother. 2012, 67, 362–366. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Sharma, C.; Sundar, G.; Singh, P.K.; Gaur, S.N.; Hagen, F.; Klaassen, C.H.; Meis, J.F. Clonal expansion and emergence of environmental multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR34/L98H mutations in the cyp51A gene in India. PLoS ONE 2012, 7, e52871. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Kathuria, S.; Hagen, F.; Meis, J.F. Prevalence and mechanism of triazole resistance in Aspergillus fumigatus in a referral chest hospital in Delhi, India and an update of the situation in Asia. Front. Microbiol. 2015, 6, 428. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Khan, Z.; Hagen, F.; Meis, J.F. Occurrence of triazole-resistant Aspergillus fumigatus with TR34/L98H mutations in outdoor and hospital environment in Kuwait. Environ. Res. 2014, 133, 20–26. [Google Scholar] [CrossRef]
- Ahangarkani, F.; Puts, Y.; Nabili, M.; Khodavaisy, S.; Moazeni, M.; Salehi, Z.; Laal Kargar, M.; Badali, H.; Meis, J.F. First azole-resistant Aspergillus fumigatus isolates with the environmental TR(46)/Y121F/T289A mutation in Iran. Mycoses 2020, 63, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; Han, X.; Tian, S.; Zhao, J.; Chen, F.; Su, X.; Zhao, J.; Zou, Z.; Gong, Y.; et al. Elevated MIC Values of Imidazole Drugs against Aspergillus fumigatus Isolates with TR(34)/L98H/S297T/F495I Mutation. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Ashu, E.E.; Korfanty, G.A.; Xu, J. Evidence of unique genetic diversity in Aspergillus fumigatus isolates from Cameroon. Mycoses 2017, 60, 739–748. [Google Scholar] [CrossRef]
- Wang, H.C.; Huang, J.C.; Lin, Y.H.; Chen, Y.H.; Hsieh, M.I.; Choi, P.C.; Lo, H.J.; Liu, W.L.; Hsu, C.S.; Shih, H.I.; et al. Prevalence, mechanisms and genetic relatedness of the human pathogenic fungus Aspergillus fumigatus exhibiting resistance to medical azoles in the environment of Taiwan. Environ. Microbiol. 2018, 20, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Takahashi, H.; Watanabe, A.; Takahashi-Nakaguchi, A.; Kawamoto, S.; Kamei, K.; Gonoi, T. Whole-genome comparison of Aspergillus fumigatus strains serially isolated from patients with aspergillosis. J. Clin. Microbiol. 2014, 52, 4202–4209. [Google Scholar] [CrossRef]
- Abdolrasouli, A.; Rhodes, J.; Beale, M.A.; Hagen, F.; Rogers, T.R.; Chowdhary, A.; Meis, J.F.; Armstrong-James, D.; Fisher, M.C. Genomic Context of Azole Resistance Mutations in Aspergillus fumigatus Determined Using Whole-Genome Sequencing. MBio 2015, 6, e00536. [Google Scholar] [CrossRef]
- Ballard, E.; Melchers, W.J.G.; Zoll, J.; Brown, A.J.P.; Verweij, P.E.; Warris, A. In-host microevolution of Aspergillus fumigatus: A phenotypic and genotypic analysis. Fungal Genet. Biol. 2018, 113, 1–13. [Google Scholar] [CrossRef]
- Ballard, E.; Zoll, J.; Melchers, W.J.G.; Brown, A.J.P.; Warris, A.; Verweij, P.E. Raw genome sequence data for 13 isogenic Aspergillus fumigatus strains isolated over a 2 year period from a patient with chronic granulomatous disease. Data Brief. 2019, 25, 104021. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Nakaguchi, A.; Muraosa, Y.; Hagiwara, D.; Sakai, K.; Toyotome, T.; Watanabe, A.; Kawamoto, S.; Kamei, K.; Gonoi, T.; Takahashi, H. Genome sequence comparison of Aspergillus fumigatus strains isolated from patients with pulmonary aspergilloma and chronic necrotizing pulmonary aspergillosis. Med. Mycol. 2015, 53, 353–360. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, B.; Walsh, T.J.; Donnelly, J.P.; Stevens, D.A.; Edwards, J.E.; Calandra, T.; Pappas, P.G.; Maertens, J.; Lortholary, O.; Kauffman, C.A.; et al. Revised definitions of invasive fungal disease from the European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group and the National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) Consensus Group. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2008, 46, 1813–1821. [Google Scholar] [CrossRef]
- Bart-Delabesse, E.; Humbert, J.F.; Delabesse, E.; Bretagne, S. Microsatellite markers for typing Aspergillus fumigatus isolates. J. Clin. Microbiol. 1998, 36, 2413–2418. [Google Scholar] [CrossRef]
- Alvarez-Perez, S.; Garcia, M.E.; Bouza, E.; Pelaez, T.; Blanco, J.L. Characterization of multiple isolates of Aspergillus fumigatus from patients: Genotype, mating type and invasiveness. Med. Mycol. 2009, 47, 601–608. [Google Scholar] [CrossRef][Green Version]
- Escribano, P.; Peláez, T.; Bouza, E.; Guinea, J. Microsatellite (STRAf) genotyping cannot differentiate between invasive and colonizing Aspergillus fumigatus isolates. J. Clin. Microbiol. 2015, 53, 667–670. [Google Scholar] [CrossRef]
- Obar, J.J. Sensing the threat posed by Aspergillus infection. Curr. Opin. Microbiol. 2020, 58, 47–55. [Google Scholar] [CrossRef] [PubMed]
- De Valk, H.A.; Klaassen, C.H.; Yntema, J.B.; Hebestreit, A.; Seidler, M.; Haase, G.; Müller, F.M.; Meis, J.F. Molecular typing and colonization patterns of Aspergillus fumigatus in patients with cystic fibrosis. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 2009, 8, 110–114. [Google Scholar] [CrossRef]
- Fraczek, M.G.; Chishimba, L.; Niven, R.M.; Bromley, M.; Simpson, A.; Smyth, L.; Denning, D.W.; Bowyer, P. Corticosteroid treatment is associated with increased filamentous fungal burden in allergic fungal disease. J. Allergy Clin. Immunol. 2018, 142, 407–414. [Google Scholar] [CrossRef]
- Latgé, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33. [Google Scholar] [CrossRef]
- O’Gorman, C.M. Airborne Aspergillus fumigatus conidia: A risk factor for aspergillosis. Fungal Biol. Rev. 2011, 25, 151–157. [Google Scholar] [CrossRef]
- Kanamori, H.; Rutala, W.A.; Sickbert-Bennett, E.E.; Weber, D.J. Review of fungal outbreaks and infection prevention in healthcare settings during construction and renovation. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 61, 433–444. [Google Scholar] [CrossRef]
- Vonberg, R.P.; Gastmeier, P. Nosocomial aspergillosis in outbreak settings. J. Hosp. Infect. 2006, 63, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.; Amorim, A.; Gusmão, L. Genetic diversity of Aspergillus fumigatus in indoor hospital environments. Med. Mycol. 2010, 48, 832–838. [Google Scholar] [CrossRef]
- Symoens, F.; Burnod, J.; Lebeau, B.; Viviani, M.A.; Piens, M.A.; Tortorano, A.M.; Nolard, N.; Chapuis, F.; Grillot, R. Hospital-acquired Aspergillus fumigatus infection: Can molecular typing methods identify an environmental source? J. Hosp. Infect. 2002, 52, 60–67. [Google Scholar] [CrossRef]
- Diba, K.; Jangi, F.; Makhdoomi, K.; Moshiri, N.; Mansouri, F. Aspergillus diversity in the environments of nosocomial infection cases at a university hospital. J. Med. Life 2019, 12, 128–132. [Google Scholar] [CrossRef]
- Loeffert, S.T.; Melloul, E.; Gustin, M.P.; Hénaff, L.; Guillot, C.; Dupont, D.; Wallon, M.; Cassier, P.; Dananché, C.; Bénet, T.; et al. Investigation of the Relationships Between Clinical and Environmental Isolates of Aspergillus fumigatus by Multiple-locus Variable Number Tandem Repeat Analysis During Major Demolition Work in a French Hospital. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 68, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Mellado, E.; Diaz-Guerra, T.M.; Cuenca-Estrella, M.; Buendia, V.; Aspa, J.; Prieto, E.; Villagrasa, J.R.; Rodriguez-Tudela, J.L. Characterization of a possible nosocomial aspergillosis outbreak. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2000, 6, 543–548. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Etienne, K.A.; Subudhi, C.P.; Chadwick, P.R.; Settle, P.; Moise, J.; Magill, S.S.; Chiller, T.; Balajee, S.A. Investigation of a cluster of cutaneous aspergillosis in a neonatal intensive care unit. J. Hosp. Infect. 2011, 79, 344–348. [Google Scholar] [CrossRef]
- Lutz, B.D.; Jin, J.; Rinaldi, M.G.; Wickes, B.L.; Huycke, M.M. Outbreak of invasive Aspergillus infection in surgical patients, associated with a contaminated air-handling system. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2003, 37, 786–793. [Google Scholar] [CrossRef]
- Peláez, T.; Muñoz, P.; Guinea, J.; Valerio, M.; Giannella, M.; Klaassen, C.H.; Bouza, E. Outbreak of invasive aspergillosis after major heart surgery caused by spores in the air of the intensive care unit. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2012, 54, e24–e31. [Google Scholar] [CrossRef] [PubMed]
- Wirmann, L.; Ross, B.; Reimann, O.; Steinmann, J.; Rath, P.M. Airborne Aspergillus fumigatus spore concentration during demolition of a building on a hospital site, and patient risk determination for invasive aspergillosis including azole resistance. J. Hosp. Infect. 2018, 100, e91–e97. [Google Scholar] [CrossRef]
- Bonnal, C.; Leleu, C.; Brugière, O.; Chochillon, C.; Porcher, R.; Boelle, P.Y.; Menotti, J.; Houze, S.; Lucet, J.C.; Derouin, F. Relationship between Fungal Colonisation of the Respiratory Tract in Lung Transplant Recipients and Fungal Contamination of the Hospital Environment. PLoS ONE 2015, 10, e0144044. [Google Scholar] [CrossRef]
- Talento, A.F.; Fitzgerald, M.; Redington, B.; O’Sullivan, N.; Fenelon, L.; Rogers, T.R. Prevention of healthcare-associated invasive aspergillosis during hospital construction/renovation works. J. Hosp. Infect. 2019, 103, 1–12. [Google Scholar] [CrossRef]
- Weber, D.J.; Peppercorn, A.; Miller, M.B.; Sickbert-Benett, E.; Rutala, W.A. Preventing healthcare-associated Aspergillus infections: Review of recent CDC/HICPAC recommendations. Med. Mycol. 2009, 47, S199–S209. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, E.; Bausman, D.; Fallah Yakhdani, H. Renovation in hospitals: An experimental study of negative pressure and filtration. Sci. Technol. Built Environ. 2020, 26, 377–386. [Google Scholar] [CrossRef]
- Mousavi, E.S.; Grosskopf, K.R. Renovation in hospitals: A case study of source control ventilation in work zones. Adv. Build. Energy Res. 2020, 14, 115–128. [Google Scholar] [CrossRef]
- Pouvaret, A.; Tavernier, E.; Cornillon, J.; Daguenet, E.; Raberin, H.; Grattard, F.; Berthelot, P.; Guyotat, D. Performance evaluation of a new mobile air-treatment technology at-rest and under normal work conditions in a conventional hematology room. Health Technol. 2020, 10, 1591–1602. [Google Scholar] [CrossRef]
- Lemaire, B.; Normand, A.C.; Forel, J.M.; Cassir, N.; Piarroux, R.; Ranque, S. Hospitalized Patient as Source of Aspergillus fumigatus, 2015. Emerg. Infect. Dis. 2018, 24, 1524–1527. [Google Scholar] [CrossRef]
- Pegues, D.A.; Lasker, B.A.; McNeil, M.M.; Hamm, P.M.; Lundal, J.L.; Kubak, B.M. Cluster of cases of invasive aspergillosis in a transplant intensive care unit: Evidence of person-to-person airborne transmission. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2002, 34, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.G.P.; Erren, E.; Vanden Driessche, K.S.J.; Melchers, W.J.G.; Reijers, M.H.; Merkus, P.; Verweij, P.E. Aerosol Transmission of Aspergillus fumigatus in Cystic Fibrosis Patients in the Netherlands. Emerg. Infect. Dis. 2019, 25, 797–799. [Google Scholar] [CrossRef]
- Bertout, S.; Renaud, F.; De Meeüs, T.; Piens, M.A.; Lebeau, B.; Viviani, M.A.; Mallié, M.; Bastide, J.M.; The Ebga Network. Multilocus enzyme electrophoresis analysis of Aspergillus fumigatus strains isolated from the first clinical sample from patients with invasive aspergillosis. J. Med. Microbiol. 2000, 49, 375–381. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Klaassen, C.H.; Gibbons, J.G.; Fedorova, N.D.; Meis, J.F.; Rokas, A. Evidence for genetic differentiation and variable recombination rates among Dutch populations of the opportunistic human pathogen Aspergillus fumigatus. Mol. Ecol. 2012, 21, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Ashu, E.; Sharma, C.; Kathuria, S.; Chowdhary, A.; Xu, J. Diversity and origins of Indian multi-triazole resistant strains of Aspergillus fumigatus. Mycoses 2016, 59, 450–466. [Google Scholar] [CrossRef]
- Duarte-Escalante, E.; Frías-De-León, M.G.; Martínez-Herrera, E.; Acosta-Altamirano, G.; de Paz, E.R.; Reséndiz-Sánchez, J.; Refojo, N.; Reyes-Montes, M.D.R. Identification of CSP Types and Genotypic Variability of Clinical and Environmental Isolates of Aspergillus fumigatus from Different Geographic Origins. Microorganisms 2020, 8, 688. [Google Scholar] [CrossRef]
- Ashu, E.E.; Hagen, F.; Chowdhary, A.; Meis, J.F.; Xu, J. Global Population Genetic Analysis of Aspergillus fumigatus. mSphere 2017, 2. [Google Scholar] [CrossRef]
- O’Gorman, C.M.; Fuller, H.; Dyer, P.S. Discovery of a sexual cycle in the opportunistic fungal pathogen Aspergillus fumigatus. Nature 2009, 457, 471–474. [Google Scholar] [CrossRef]
- Teixeira, J.; Amorim, A.; Araujo, R. Recombination detection in Aspergillus fumigatus through single nucleotide polymorphisms typing. Environ. Microbiol. Rep. 2015, 7, 881–886. [Google Scholar] [CrossRef]
- Bain, J.M.; Tavanti, A.; Davidson, A.D.; Jacobsen, M.D.; Shaw, D.; Gow, N.A.; Odds, F.C. Multilocus sequence typing of the pathogenic fungus Aspergillus fumigatus. J. Clin. Microbiol. 2007, 45, 1469–1477. [Google Scholar] [CrossRef]
- Losada, L.; Sugui, J.A.; Eckhaus, M.A.; Chang, Y.C.; Mounaud, S.; Figat, A.; Joardar, V.; Pakala, S.B.; Pakala, S.; Venepally, P.; et al. Genetic Analysis Using an Isogenic Mating Pair of Aspergillus fumigatus Identifies Azole Resistance Genes and Lack of MAT Locus’s Role in Virulence. PLoS Pathog. 2015, 11, e1004834. [Google Scholar] [CrossRef]
- Pugliese, M.; Matić, S.; Prethi, S.; Gisi, U.; Gullino, M.L. Molecular characterization and sensitivity to demethylation inhibitor fungicides of Aspergillus fumigatus from orange-based compost. PLoS ONE 2018, 13, e0200569. [Google Scholar] [CrossRef]
- Zhang, J.; Snelders, E.; Zwaan, B.J.; Schoustra, S.E.; Meis, J.F.; van Dijk, K.; Hagen, F.; van der Beek, M.T.; Kampinga, G.A.; Zoll, J.; et al. A Novel Environmental Azole Resistance Mutation in Aspergillus fumigatus and a Possible Role of Sexual Reproduction in Its Emergence. MBio 2017, 8. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–s444. [Google Scholar] [CrossRef]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2018, 24, e1–e38. [Google Scholar] [CrossRef]
- Hagiwara, D.; Watanabe, A.; Kamei, K.; Goldman, G.H. Epidemiological and Genomic Landscape of Azole Resistance Mechanisms in Aspergillus Fungi. Front. Microbiol. 2016, 7, 1382. [Google Scholar] [CrossRef]
- Verweij, P.E.; Ananda-Rajah, M.; Andes, D.; Arendrup, M.C.; Brüggemann, R.J.; Chowdhary, A.; Cornely, O.A.; Denning, D.W.; Groll, A.H.; Izumikawa, K.; et al. International expert opinion on the management of infection caused by azole-resistant Aspergillus fumigatus. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer Chemother. 2015, 21–22, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Hokken, M.W.J.; Zoll, J.; Coolen, J.P.M.; Zwaan, B.J.; Verweij, P.E.; Melchers, W.J.G. Phenotypic plasticity and the evolution of azole resistance in Aspergillus fumigatus; an expression profile of clinical isolates upon exposure to itraconazole. BMC Genom. 2019, 20, 28. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.J.; Cerar, D.; Anderson, M.J.; Albarrag, A.; Fisher, M.C.; Pasqualotto, A.C.; Laverdiere, M.; Arendrup, M.C.; Perlin, D.S.; Denning, D.W. Frequency and Evolution of Azole Resistance in Aspergillus fumigatus Associated with Treatment Failure. Emerg. Infect. Dis. J. 2009, 15, 1068. [Google Scholar] [CrossRef]
- Nywening, A.V.; Rybak, J.M.; Rogers, P.D.; Fortwendel, J.R. Mechanisms of triazole resistance in Aspergillus fumigatus. Environ. Microbiol. 2020, 22, 4934–4952. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Lucas, J.A.; Arendrup, M.C.; Bowyer, P.; Brinkmann, A.J.F.; Denning, D.W.; Dyer, P.S.; Fisher, M.C.; Geenen, P.L.; Gisi, U.; et al. The one health problem of azole resistance in Aspergillus fumigatus: Current insights and future research agenda. Fungal Biol. Rev. 2020, 34, 202–214. [Google Scholar] [CrossRef]
- Hare, R.K.; Gertsen, J.B.; Astvad, K.M.T.; Degn, K.B.; Løkke, A.; Stegger, M.; Andersen, P.S.; Kristensen, L.; Arendrup, M.C. In Vivo Selection of a Unique Tandem Repeat Mediated Azole Resistance Mechanism (TR(120)) in Aspergillus fumigatus cyp51A, Denmark. Emerg. Infect. Dis. 2019, 25, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Ziogas, B.N.; Malandrakis, A.A. Sterol Biosynthesis Inhibitors: C14 Demethylation (DMIs). In Fungicide Resistance in Plant Pathogens: Principles and a Guide to Practical Management; Ishii, H., Hollomon, D.W., Eds.; Springer: Japan, Tokyo, 2015; pp. 199–216. [Google Scholar] [CrossRef]
- Snelders, E.; Huis In’t Veld, R.A.; Rijs, A.J.; Kema, G.H.; Melchers, W.J.; Verweij, P.E. Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl. Environ. Microbiol. 2009, 75, 4053–4057. [Google Scholar] [CrossRef]
- Snelders, E.; Camps, S.M.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.; Verweij, P.E. Triazole fungicides can induce cross-resistance to medical triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef] [PubMed]
- Beer, K.D.; Farnon, E.C.; Jain, S.; Jamerson, C.; Lineberger, S.; Miller, J.; Berkow, E.L.; Lockhart, S.R.; Chiller, T.; Jackson, B.R. Multidrug-Resistant Aspergillus fumigatus Carrying Mutations Linked to Environmental Fungicide Exposure-Three States, 2010–2017. Mmwr. Morb. Mortal. Wkly. Rep. 2018, 67, 1064–1067. [Google Scholar] [CrossRef]
- Prigitano, A.; Esposto, M.C.; Biffi, A.; De Lorenzis, G.; Favuzzi, V.; Koncan, R.; Lo Cascio, G.; Barao Ocampo, M.; Colombo, C.; Pizzamiglio, G.; et al. Triazole resistance in Aspergillus fumigatus isolates from patients with cystic fibrosis in Italy. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 2017, 16, 64–69. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Riat, A.; Plojoux, J.; Gindro, K.; Schrenzel, J.; Sanglard, D. Azole Resistance of Environmental and Clinical Aspergillus fumigatus; Isolates from Switzerland. Antimicrob. Agents Chemother. 2018, 62, e02088-17. [Google Scholar] [CrossRef]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole Resistance in Aspergillus fumigatus: A Consequence of Antifungal Use in Agriculture? Front. Microbiol. 2017, 8, 1024. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of Azole-Resistant Aspergillus fumigatus Strains due to Agricultural Azole Use Creates an Increasing Threat to Human Health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Infect. Dis. 2019, 25, 1347–1353. [CrossRef] [PubMed]
- Resendiz Sharpe, A.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, P.E.; on behalf of the ISHAM/ECMM Aspergillus Resistance Surveillance Working Group. Triazole resistance surveillance in Aspergillus fumigatus. Med. Mycol. 2018, 56, S83–S92. [Google Scholar] [CrossRef]
- Buil, J.B.; Hare, R.K.; Zwaan, B.J.; Arendrup, M.C.; Melchers, W.J.G.; Verweij, P.E. The fading boundaries between patient and environmental routes of triazole resistance selection in Aspergillus fumigatus. PLoS Pathog. 2019, 15, e1007858. [Google Scholar] [CrossRef]
- Berkow, E.L.; Nunnally, N.S.; Bandea, A.; Kuykendall, R.; Beer, K.; Lockhart, S.R. Detection of TR(34)/L98H CYP51A Mutation through Passive Surveillance for Azole-Resistant Aspergillus fumigatus in the United States from 2015 to 2017. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Ren, J.; Jin, X.; Zhang, Q.; Zheng, Y.; Lin, D.; Yu, Y. Fungicides induced triazole-resistance in Aspergillus fumigatus associated with mutations of TR46/Y121F/T289A and its appearance in agricultural fields. J. Hazard. Mater. 2017, 326, 54–60. [Google Scholar] [CrossRef]
- Barber, A.E.; Riedel, J.; Sae-Ong, T.; Kang, K.; Brabetz, W.; Panagiotou, G.; Deising, H.B.; Kurzai, O. Effects of Agricultural Fungicide Use on Aspergillus fumigatus Abundance, Antifungal Susceptibility, and Population Structure. MBio 2020, 11, e02213–e02220. [Google Scholar] [CrossRef]
- Deng, S.; Zhang, L.; Ji, Y.; Verweij, P.E.; Tsui, K.M.; Hagen, F.; Houbraken, J.; Meis, J.F.; Abliz, P.; Wang, X.; et al. Triazole phenotypes and genotypic characterization of clinical Aspergillus fumigatus isolates in China. Emerg. Microbes Infect. 2017, 6, e109. [Google Scholar] [CrossRef]
- Sharma, C.; Hagen, F.; Moroti, R.; Meis, J.F.; Chowdhary, A. Triazole-resistant Aspergillus fumigatus harbouring G54 mutation: Is it de novo or environmentally acquired? J. Glob. Antimicrob. Resist. 2015, 3, 69–74. [Google Scholar] [CrossRef]
- Korfanty, G.A.; Teng, L.; Pum, N.; Xu, J. Contemporary Gene Flow is a Major Force Shaping the Aspergillus fumigatus Population in Auckland, New Zealand. Mycopathologia 2019, 184, 479–492. [Google Scholar] [CrossRef]
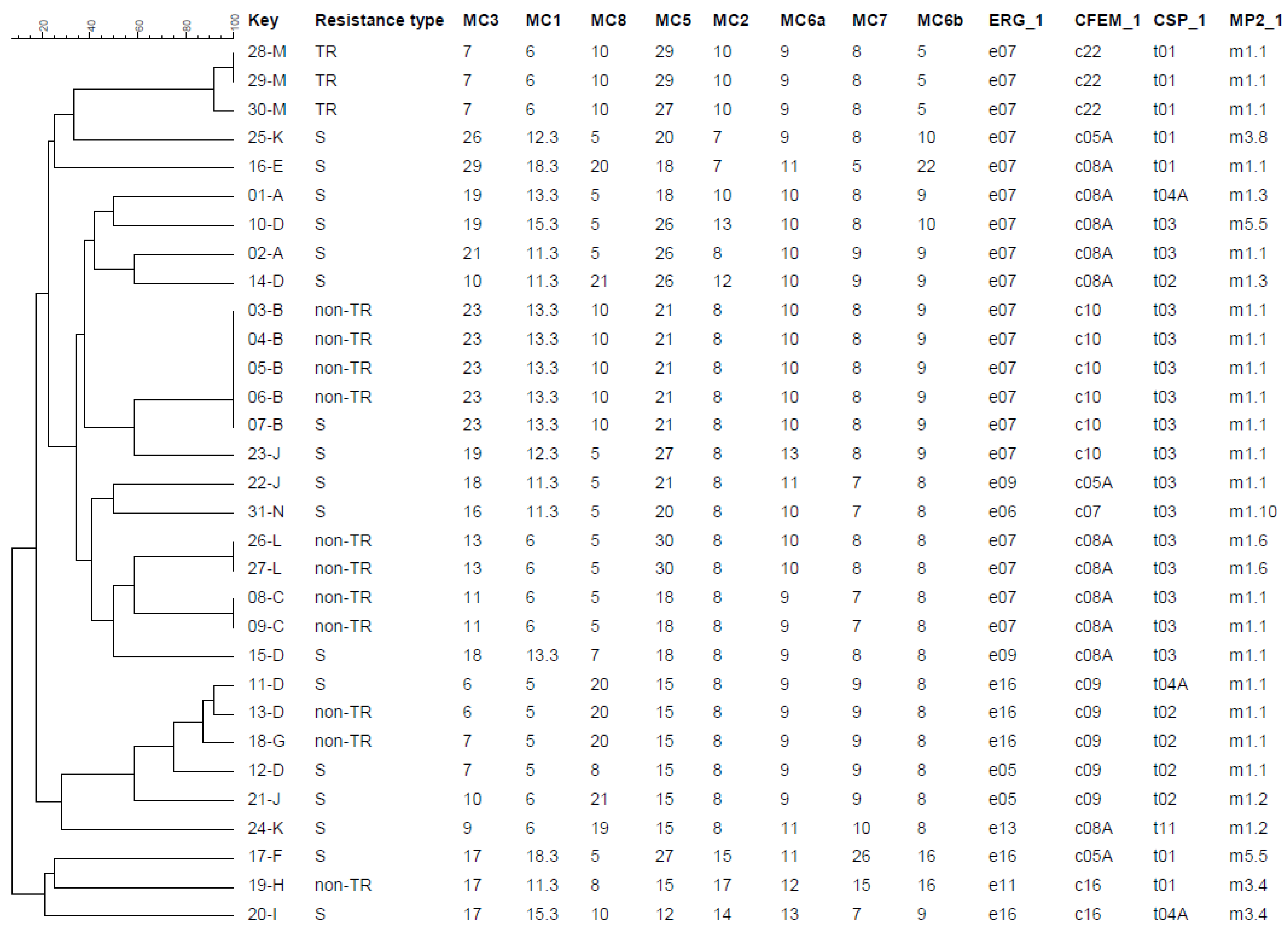
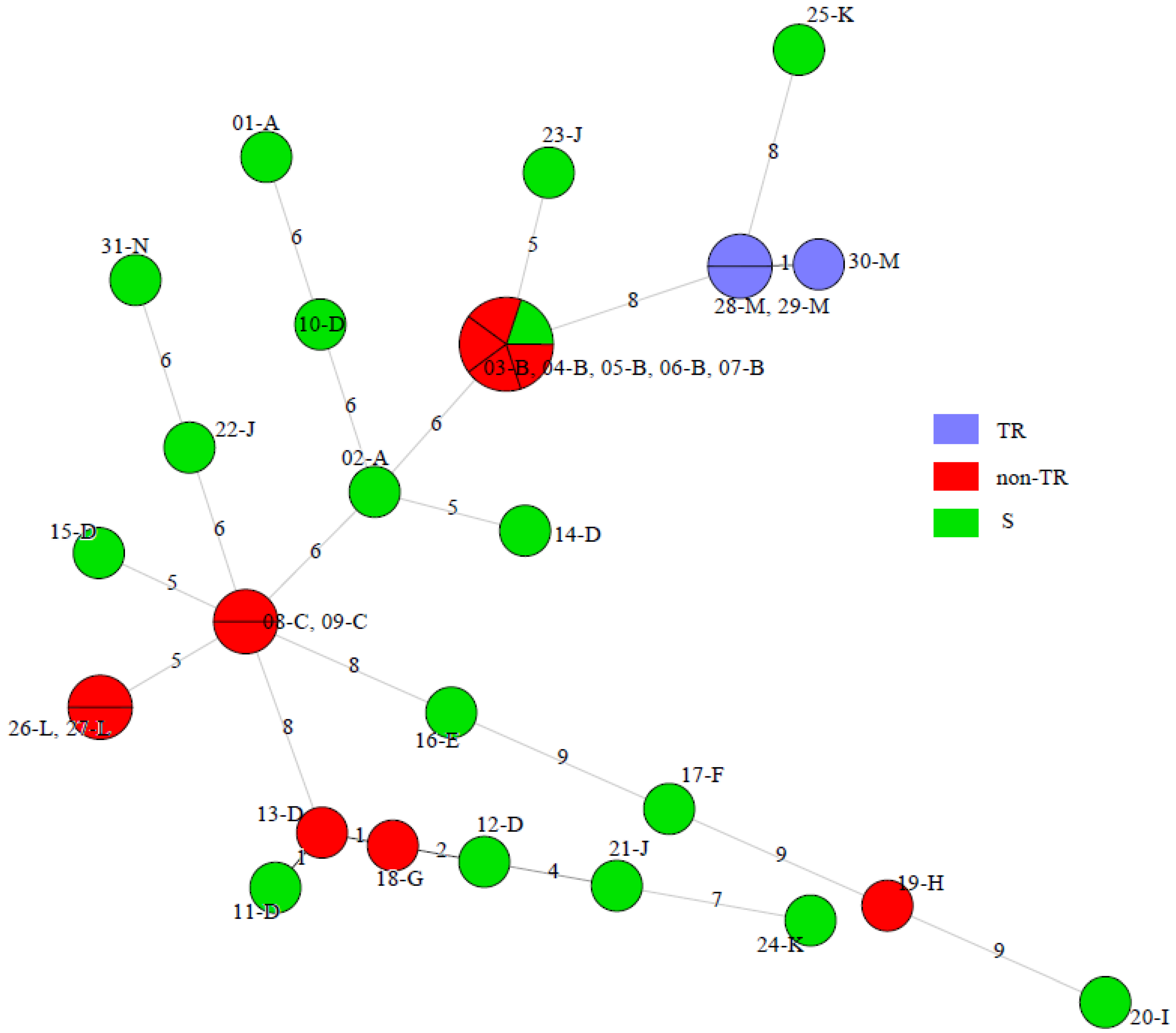
- Novak-Frazer, L.; Shen, H.; Masania, R.; Bowyer, P.; Rautemaa-Richardson, R.; Richardson, M.D. Comparison of two genotyping methods reveals clusters of resistant Aspergillus fumigatus from chronic pulmonary aspergillosis patients in the UK. In Proceedings of the Congress of the International Society for Human and Animal Mycology, Amsterdam, The Netherlands, 30 June–4 July 2018; p. 250. [Google Scholar]
- Rodriquez-Tudela, J.L.; Donnelly, J.P.; Arendrup, M.C.; Arikan, S.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Denning, D.; et al. EUCAST Technical Note on the method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia-forming moulds. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2008, 14, 982–984. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Typing Technique | Principle | Discriminatory Power 1 | Advantage(s) | Disadvantage(s) | Reference(s) |
|---|---|---|---|---|---|
| TRESP | hypervariable TRs 2 in 3 genes | >0.997 | specialist equipment not required ease of interpretation, reproducibility | lower resolution | [44] |
| including cyp51A resistance markers | 0.890 | ||||
| TRESPERG | hypervariable TRs in 4 genes | >0.997 | [46] | ||
| including cyp51A resistance markers | >0.993 | ||||
| TRESPERG + STRAf | hypervariable TRs in 4 genes plus 9 STRs 3 and cyp51A resistance markers | >0.999 | high-resolution fingerprinting | technical expertise specialist equipment | [46] |
| STRAf | 9 STRs | >0.999 | high-resolution fingerprinting | technical expertise specialist equipment reproducibility interlaboratory variation | [30] |
| nSTRAf | 8 STRs | 0.9997 | high-resolution fingerprinting single multiplex reaction longer, more accurate STRs | technical expertise specialist equipment | [40] |
| STRAf+ | 9 STRs and cyp51A resistance markers | na 4 | high-resolution fingerprinting public database (AfumID) | technical expertise specialist equipment | [43] |
| NGS/WGS 5 | genome | >0.999 | maximum resolution | technical expertise specialist equipment bioinformatics capabilities | [65,66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Torre, M.H.; Shen, H.; Rautemaa-Richardson, R.; Richardson, M.D.; Novak-Frazer, L. Molecular Epidemiology of Aspergillus fumigatus in Chronic Pulmonary Aspergillosis Patients. J. Fungi 2021, 7, 152. https://doi.org/10.3390/jof7020152
van der Torre MH, Shen H, Rautemaa-Richardson R, Richardson MD, Novak-Frazer L. Molecular Epidemiology of Aspergillus fumigatus in Chronic Pulmonary Aspergillosis Patients. Journal of Fungi. 2021; 7(2):152. https://doi.org/10.3390/jof7020152
Chicago/Turabian Stylevan der Torre, Mireille H., Hongwei Shen, Riina Rautemaa-Richardson, Malcolm D. Richardson, and Lilyann Novak-Frazer. 2021. "Molecular Epidemiology of Aspergillus fumigatus in Chronic Pulmonary Aspergillosis Patients" Journal of Fungi 7, no. 2: 152. https://doi.org/10.3390/jof7020152
APA Stylevan der Torre, M. H., Shen, H., Rautemaa-Richardson, R., Richardson, M. D., & Novak-Frazer, L. (2021). Molecular Epidemiology of Aspergillus fumigatus in Chronic Pulmonary Aspergillosis Patients. Journal of Fungi, 7(2), 152. https://doi.org/10.3390/jof7020152