The Squeaky Yeast Gets Greased: The Roles of Host Lipids in the Clearance of Pathogenic Fungi

Abstract
1. Introduction
2. The Offense
3. The Defense
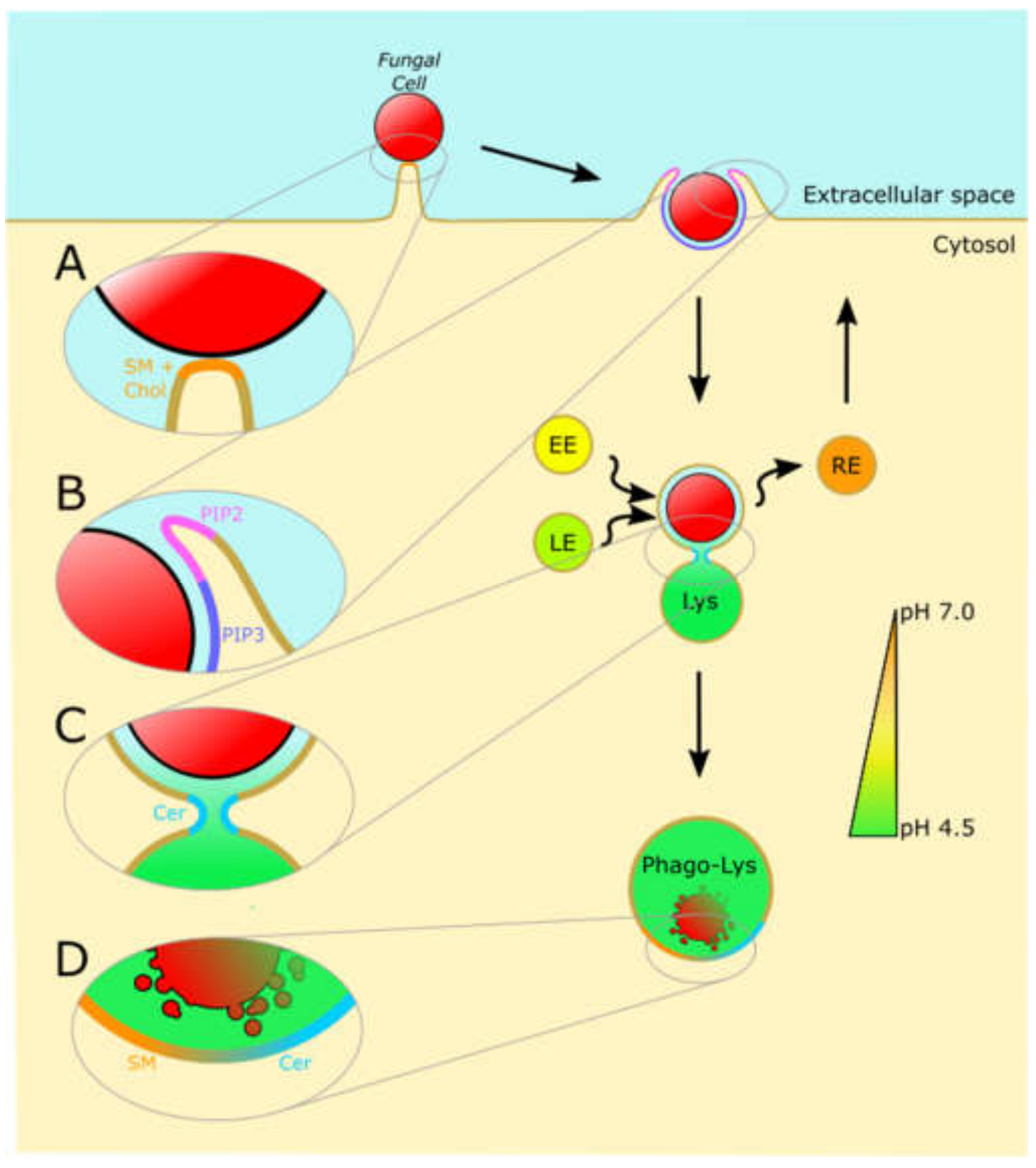
4. Phagocytosis: The Weapon of Choice
5. The Phagolysosome: The Final Stage for Eliminating Fungal Pathogens
6. Conclusions and Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Cauchie, M.; Desmet, S.; Lagrou, K. Candida and Its Dual Lifestyle as a Commensal and a Pathogen. Res. Microbiol. 2017, 168, 802–810. [Google Scholar] [CrossRef]
- Iliev, I.D.; Leonardi, I. Fungal Dysbiosis: Immunity and Interactions at Mucosal Barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida Albicans Pathogenicity Mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Benedict, K.; Richardson, M.; Vallabhaneni, S.; Jackson, B.R.; Chiller, T. Emerging Issues, Challenges, and Changing Epidemiology of Fungal Disease Outbreaks. Lancet Infect. Dis. 2017, 17, e403–e411. [Google Scholar] [CrossRef]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections; Humana Press: New York, NY, USA, 2017; pp. 17–65. [Google Scholar]
- Pustylnikov, S.; Sagar, D.; Jain, P.; Khan, Z.K. Targeting the C-Type Lectins-Mediated Host-Pathogen Interactions with Dextran. J. Pharm. Pharm. Sci. 2014, 17, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chandawarkar, R.Y. Phagocytosis of Fungal Agents and Yeast via Macrophage Cell Surface Scavenger Receptors. J. Surg. Res. 2010, 164, e273–e279. [Google Scholar] [CrossRef]
- Guo, M.; Härtlova, A.; Dill, B.D.; Prescott, A.R.; Gierliński, M.; Trost, M. High-Resolution Quantitative Proteome Analysis Reveals Substantial Differences between Phagosomes of RAW 264.7 and Bone Marrow Derived Macrophages. Proteomics 2015, 15, 3169–3174. [Google Scholar] [CrossRef]
- Richardson, J.P.; Moyes, D.L. Adaptive Immune Responses to Candida Albicans Infection. Virulence 2015, 6, 327–337. [Google Scholar] [CrossRef]
- Kerrigan, A.M.; Brown, G.D. Syk-Coupled C-Type Lectin Receptors That Mediate Cellular Activation via Single Tyrosine Based Activation Motifs. Immunol. Rev. 2010, 234, 335–352. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-Glucan Recognition by the Innate Immune System. Immunological Reviews 2009, 38–50. [Google Scholar] [CrossRef]
- Li, X.; Utomo, A.; Cullere, X.; Choi, M.M.; Milner, D.A.; Venkatesh, D.; Yun, S.H.; Mayadas, T.N. The β-Glucan Receptor Dectin-1 Activates the Integrin Mac-1 in Neutrophils via Vav Protein Signaling to Promote Candida Albicans Clearance. Cell Host Microbe 2011, 10, 603–615. [Google Scholar] [CrossRef]
- Goyal, S.; Castrillón-Betancur, J.C.; Klaile, E.; Slevogt, H. The Interaction of Human Pathogenic Fungi With C-Type Lectin Receptors. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Wells, C.A.; Salvage-Jones, J.A.; Li, X.; Hitchens, K.; Butcher, S.; Murray, R.Z.; Beckhouse, A.G.; Lo, Y.-L.-S.; Manzanero, S.; Cobbold, C.; et al. The Macrophage-Inducible C-Type Lectin, Mincle, Is an Essential Component of the Innate Immune Response to Candida Albicans. J. Immunol. 2008, 180, 7404–7413. [Google Scholar] [CrossRef]
- Yamasaki, S.; Matsumoto, M.; Takeuchi, O.; Matsuzawa, T.; Ishikawa, E.; Sakuma, M.; Tateno, H.; Uno, J.; Hirabayashi, J.; Mikami, Y.; et al. C-Type Lectin Mincle Is an Activating Receptor for Pathogenic Fungus, Malassezia. Proc. Natl. Acad. Sci. USA. 2009, 106, 1897–1902. [Google Scholar] [CrossRef]
- Serrano-Gómez, D.; Antonio Leal, J.; Corbí, A.L. DC-SIGN Mediates the Binding of Aspergillus Fumigatus and Keratinophylic Fungi by Human Dendritic Cells. Immunobiology 2005, 210, 175–183. [Google Scholar] [CrossRef]
- Jouault, T.; El Abed-El Behi, M.; Martínez-Esparza, M.; Breuilh, L.; Trinel, P.-A.; Chamaillard, M.; Trottein, F.; Poulain, D. Specific Recognition of Candida Albicans by Macrophages Requires Galectin-3 to Discriminate Saccharomyces Cerevisiae and Needs Association with TLR2 for Signaling. J. Immunol. 2006, 177, 4679–4687. [Google Scholar] [CrossRef]
- Brown, G.D.; Taylor, P.R.; Reid, D.M.; Willment, J.A.; Williams, D.L.; Martinez-Pomares, L.; Wong, S.Y.C.; Gordon, S. Dectin-1 Is A Major β-Glucan Receptor On Macrophages. J. Exp. Med. 2002, 196, 407–412. [Google Scholar] [CrossRef]
- Gantner, B.N.; Simmons, R.M.; Canavera, S.J.; Akira, S.; Underhill, D.M. Collaborative Induction of Inflammatory Responses by Dectin-1 and Toll-like Receptor 2. J. Exp. Med. 2003, 197, 1107–1117. [Google Scholar] [CrossRef]
- Hawn, T.R.; Underhill, D.M. Toll-like Receptors in Innate Immunity. In Measuring Immunity: Basic Biology and Clinical Assessment; Academic Press: Cambridge, MA, USA, 2005; Volume 17, pp. 80–90. [Google Scholar]
- Inoue, M.; Shinohara, M.L. Clustering of Pattern Recognition Receptors for Fungal Detection. PLoS Pathog. 2014, 10, 2–4. [Google Scholar] [CrossRef]
- Skold, M.; Behar, S.M. Role of CD1d-Restricted NKT Cells in Microbial Immunity. Infect. Immun. 2003, 71, 5447–5455. [Google Scholar] [CrossRef]
- Pereira, C.S.; Macedo, M.F. CD1-Restricted T Cells at the Crossroad of Innate and Adaptive Immunity. J. Immunol. Res. 2016, 2016. [Google Scholar] [CrossRef]
- Kawakami, K.; Kinjo, Y.; Yara, S.; Koguchi, Y.; Uezu, K.; Nakayama, T.; Taniguchi, M.; Saito, A. Activation of Vα14+ Natural Killer T Cells by α-Galactosylceramide Results in Development of Th1 Response and Local Host Resistance in Mice Infected with Cryptococcus Neoformans. Infect. Immun. 2001, 69, 213–220. [Google Scholar] [CrossRef]
- Induces, R.; Disease, A. A Fungal Glycosphingolipid Directly Activates Natural Killer T Cells and Rapidly Induces Airways Disease. Nat. Med. 2014, 19, 1297–1304. [Google Scholar] [CrossRef]
- Bird, D.C.; Sheagren, J.N. Evaluation of Reticuloendothelial System Phagocytic Activity During Systemic Candida Albicans Infection in Mice. Exp. Biol. Med. 1970, 133, 34–37. [Google Scholar] [CrossRef]
- Baine, W.B.; Koenig, M.G.; Goodman, J.S. Clearance of Candida Albicans from the Bloodstream of Rabbits. Infect. Immun. 1974, 10, 1420–1425. [Google Scholar] [CrossRef]
- Sawyer, R.T.; Moon, R.J.; Beneke, E.S. Hepatic Clearance of Candida Albicans in Rats. Infect. Immun. 1976, 14, 1348–1355. [Google Scholar] [CrossRef]
- Stuart, L.M.; Ezekowitz, R.A.B. Phagocytosis: Elegant Complexity. Immunity 2005, 22, 539–550. [Google Scholar] [CrossRef]
- Rosales, C.; Uribe-Querol, E. Phagocytosis: A Fundamental Process in Immunity. Biomed Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Zhao, H.; Lappalainen, P. A Simple Guide to Biochemical Approaches for Analyzing Protein–Lipid Interactions. Mol. Biol. Cell 2012, 23, 2823–2830. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Reyes, C.N.; Becker, C.A.; Katsumoto, T.R.; Ma, J.; Wolf, A.J.; Bose, N.; Chan, A.S.H.; Magee, A.S.; Danielson, M.E.; et al. Activation of the Innate Immune Receptor Dectin-1 upon Formation of a ‘Phagocytic Synapse’. Nature 2011, 472, 471–475. [Google Scholar] [CrossRef]
- Yoshizaki, F.; Nakayama, H.; Iwahara, C.; Takamori, K.; Ogawa, H.; Iwabuchi, K. Role of Glycosphingolipid-Enriched Microdomains in Innate Immunity: Microdomain-Dependent Phagocytic Cell Functions. Biochim. Biophys. Acta - Gen. Subj. 2008, 1780, 383–392. [Google Scholar] [CrossRef]
- Iwabuchi, K.; Nagaoka, I. Lactosylceramide-Enriched Glycosphingolipid Signaling Domain Mediates Superoxide Generation from Human Neutrophils. Blood 2002, 100, 1454–1464. [Google Scholar] [CrossRef]
- Xu, S.; Huo, J.; Gunawan, M.; Su, I.-H.; Lam, K.-P. Activated Dectin-1 Localizes to Lipid Raft Microdomains for Signaling and Activation of Phagocytosis and Cytokine Production in Dendritic Cells. J. Biol. Chem. 2009, 284, 22005–22011. [Google Scholar] [CrossRef]
- Kinoshita, M.; Suzuki, K.G.N.; Matsumori, N.; Takada, M.; Ano, H.; Morigaki, K.; Abe, M.; Makino, A.; Kobayashi, T.; Hirosawa, K.M.; et al. Raft-Based Sphingomyelin Interactions Revealed by New Fluorescent Sphingomyelin Analogs. J. Cell Biol. 2017, 216, 1183–1204. [Google Scholar] [CrossRef]
- Kinoshita, M.; Suzuki, K.G.N.; Murata, M.; Matsumori, N. Evidence of Lipid Rafts Based on the Partition and Dynamic Behavior of Sphingomyelins. Chem. Phys. Lipids 2018, 215, 84–95. [Google Scholar] [CrossRef]
- Tafesse, F.G.; Rashidfarrokhi, A.; Schmidt, F.I.; Freinkman, E.; Dougan, S.; Dougan, M.; Esteban, A.; Maruyama, T.; Strijbis, K.; Ploegh, H.L. Disruption of Sphingolipid Biosynthesis Blocks Phagocytosis of Candida Albicans. PLOS Pathog. 2015, 11, e1005188. [Google Scholar] [CrossRef]
- Niekamp, P.; Guzman, G.; Leier, H.; Rashidfarrokhi, A.; Richina, V.; Pott, F.; Holthuis, J.C.M.; Tafesse, F.G. Sphingomyelin Biosynthesis Is Essential for Phagocytic Signaling during Mycobacterium Tuberculosis Host Cell Entry. bioRxiv 2019, 565226. [Google Scholar] [CrossRef]
- Viswanathan, G.; Jafurulla, M.; Kumar, G.A.; Raghunand, T.R.; Chattopadhyay, A. Macrophage Sphingolipids Are Essential for the Entry of Mycobacteria. Chem. Phys. Lipids 2018, 213, 25–31. [Google Scholar] [CrossRef]
- Bryan, A.M.; Del Poeta, M.; Luberto, C. Sphingolipids as Regulators of the Phagocytic Response to Fungal Infections. Mediat. Inflamm. 2015, 2015. [Google Scholar] [CrossRef]
- Brown, G.D. Dectin-1: A Signalling Non-TLR Pattern-Recognition Receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Hemmings, B.A.; Restuccia, D.F. PI3K-PKB/Akt Pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011189. [Google Scholar] [CrossRef]
- Razzinit, G.; Brancaccio, A.; Lemmon, M.A.; Guarnieri, S.; Falasca, M. The Role of the Pleckstrin Homology Domain in Membrane Targeting and Activation of Phospholipase Cβ1. J. Biol. Chem. 2000, 275, 14873–14881. [Google Scholar] [CrossRef][Green Version]
- Levin, R.; Grinstein, S.; Schlam, D. Phosphoinositides in Phagocytosis and Macropinocytosis. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 2015, 1851, 805–823. [Google Scholar] [CrossRef]
- Mendelez, A.J. Calcium Signaling During Phagocytosis. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA; Available online: https://www.ncbi.nlm.nih.gov/books/NBK5971/ (accessed on 26 January 2020).
- Wilsson, å.; Lundqvist, H.; Gustafsson, M.; Stendahl, O. Killing of Phagocytosed Staphylococcus Aureus by Human Neutrophils Requires Intracellular Free Calcium. J. Leukoc. Biol. 1996, 59, 902–907. [Google Scholar] [CrossRef]
- Hauck, C.R.; Grassmé, H.; Bock, J.; Jendrossek, V.; Ferlinz, K.; Meyer, T.F.; Gulbins, E. Acid Sphingomyelinase Is Involved in CEACAM Receptor-Mediated Phagocytosis of Neisseria Gonorrhoeae. FEBS Lett. 2000, 478, 260–266. [Google Scholar] [CrossRef]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brugger, B.; Simons, M. Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Utermöhlen, O.; Herz, J.; Schramm, M.; Krönke, M. Fusogenicity of Membranes: The Impact of Acid Sphingomyelinase on Innate Immune Responses. Immunobiology 2008, 213, 307–314. [Google Scholar] [CrossRef]
- Grassmé, H.; Jendrossek, V.; Riehle, A.; von Kürthy, G.; Berger, J.; Schwarz, H.; Weller, M.; Kolesnick, R.; Gulbins, E. Host Defense against Pseudomonas Aeruginosa Requires Ceramide-Rich Membrane Rafts. Nat. Med. 2003, 9, 322–330. [Google Scholar] [CrossRef]
- Flannagan, R.S.; Jaumouillé, V.; Grinstein, S. The Cell Biology of Phagocytosis. Annu. Rev. Pathol. Mech. Dis. 2012, 7, 61–98. [Google Scholar] [CrossRef]
- Jordens, I.; Fernandez-Borja, M.; Marsman, M.; Dusseljee, S.; Janssen, L.; Calafat, J.; Janssen, H.; Wubbolts, R.; Neefjes, J. The Rab7 Effector Protein RILP Controls Lysosomal Transport by Inducing the Recruitment of Dynein-Dynactin Motors. Curr. Biol. 2001, 11, 1680–1685. [Google Scholar] [CrossRef]
- Harrison, R.E.; Bucci, C.; Vieira, O.V.; Schroer, T.A.; Grinstein, S. Phagosomes Fuse with Late Endosomes and/or Lysosomes by Extension of Membrane Protrusions along Microtubules: Role of Rab7 and RILP. Mol. Cell. Biol. 2003, 23, 6494–6506. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Cosío, G.; Grinstein, S. Antimicrobial Mechanisms of Phagocytes and Bacterial Evasion Strategies. Nat. Rev. Microbiol. 2009, 7, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Teichgräber, V.; Ulrich, M.; Endlich, N.; Riethmüller, J.; Wilker, B.; De Oliveira–Munding, C.C.; van Heeckeren, A.M.; Barr, M.L.; von Kürthy, G.; Schmid, K.W.; et al. Ceramide Accumulation Mediates Inflammation, Cell Death and Infection Susceptibility in Cystic Fibrosis. Nat. Med. 2008, 14, 382–391. [Google Scholar] [CrossRef]
- Smith, E.L.; Schuchman, E.H. The Unexpected Role of Acid Sphingomyelinase in Cell Death and the Pathophysiology of Common Diseases. FASEB J. 2008, 22, 3419–3431. [Google Scholar] [CrossRef]
- Teng, O.; Ang, C.K.E.; Guan, X.L. Macrophage–Bacteria Interactions—A Lipid-Centric Relationship. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Kinchen, J.M.; Ravichandran, K.S. Phagosome Maturation: Going through the Acid Test. Nature Reviews Molecular Cell Biology 2008, 9, 781–795. [Google Scholar] [CrossRef]
- Heinrich, M.; Wickel, M.; Schneider-Brachert, W.; Sandberg, C.; Gahr, J.; Schwandner, R.; Weber, T.; Brunner, J.; Krönke, M.; Schütze, S. Cathepsin D Targeted by Acid Sphingomyelinase-Derived Ceramide. EMBO J. 1999, 18, 5252–5263. [Google Scholar] [CrossRef]
- Roca, F.J.; Ramakrishnan, L. TNF Dually Mediates Resistance and Susceptibility to Mycobacteria via Mitochondrial Reactive Oxygen Species. Cell 2013, 153, 521–534. [Google Scholar] [CrossRef]
- Roca, F.J.; Whitworth, L.J.; Redmond, S.; Jones, A.A.; Ramakrishnan, L. TNF Induces Pathogenic Programmed Macrophage Necrosis in Tuberculosis through a Mitochondrial-Lysosomal-Endoplasmic Reticulum Circuit. Cell 2019, 178, 1344–1361. [Google Scholar] [CrossRef]
- Schroeder, F. Phagosomal Membrane Lipids of LM Fibroblasts. J. Membr. Biol. 1982, 68, 141–150. [Google Scholar] [CrossRef]
- Augenstreich, J.; Haanappel, E.; Ferré, G.; Czaplicki, G.; Jolibois, F.; Destainville, N.; Guilhot, C.; Milon, A.; Astarie-Dequeker, C.; Chavent, M. The Conical Shape of DIM Lipids Promotes Mycobacterium Tuberculosis Infection of Macrophages. bioRxiv 2019, 649541. [Google Scholar] [CrossRef]
- Wilburn, K.M.; Fieweger, R.A.; VanderVen, B.C. Cholesterol and Fatty Acids Grease the Wheels of Mycobacterium Tuberculosis Pathogenesis. Pathog. Dis. 2018, 76, 21. [Google Scholar] [CrossRef] [PubMed]
- Leier, H.C.; Messer, W.B.; Tafesse, F.G. Lipids and Pathogenic Flaviviruses: An Intimate Union. PLoS ONE 2018, 14, e1006952. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.; Maamar, H.; Deb, C.; Sirakova, T.D.; Kolattukudy, P.E. Mycobacterium Tuberculosis Uses Host Triacylglycerol to Accumulate Lipid Droplets and Acquires a Dormancy-like Phenotype in Lipid-Loaded Macrophages. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| PRR | PAMP | References |
|---|---|---|
| Mannose receptor | Mannan, mannoproteins | [6,7,8,9] |
| Dectin-1 | β-1,3 glucan | [9,10,11,12,13] |
| Dectin-2 | α-mannans | [2,13] |
| Mincle | α-mannose | [9,13,14,15] |
| DC-SIGN | Galactomannans | [9,13,16] |
| Galectin-3 | β-1,2 mannosides | [9,17] |
| Complement receptor 3 | C3b, β-glucans | [9,18] |
| Toll-like receptor 2 | Chitin and other polysaccharides | [9,19,20] |
| Toll-like receptor 4 | O-linked mannosyl residues | [21] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzman, G.; Niekamp, P.; Tafesse, F.G. The Squeaky Yeast Gets Greased: The Roles of Host Lipids in the Clearance of Pathogenic Fungi. J. Fungi 2020, 6, 19. https://doi.org/10.3390/jof6010019
Guzman G, Niekamp P, Tafesse FG. The Squeaky Yeast Gets Greased: The Roles of Host Lipids in the Clearance of Pathogenic Fungi. Journal of Fungi. 2020; 6(1):19. https://doi.org/10.3390/jof6010019
Chicago/Turabian StyleGuzman, Gaelen, Patrick Niekamp, and Fikadu Geta Tafesse. 2020. "The Squeaky Yeast Gets Greased: The Roles of Host Lipids in the Clearance of Pathogenic Fungi" Journal of Fungi 6, no. 1: 19. https://doi.org/10.3390/jof6010019
APA StyleGuzman, G., Niekamp, P., & Tafesse, F. G. (2020). The Squeaky Yeast Gets Greased: The Roles of Host Lipids in the Clearance of Pathogenic Fungi. Journal of Fungi, 6(1), 19. https://doi.org/10.3390/jof6010019