Sporothrix schenckii Cell Wall Proteins-Stimulated BMDCs Are Able to Induce a Th1-Prone Cytokine Profile In Vitro


 and
and 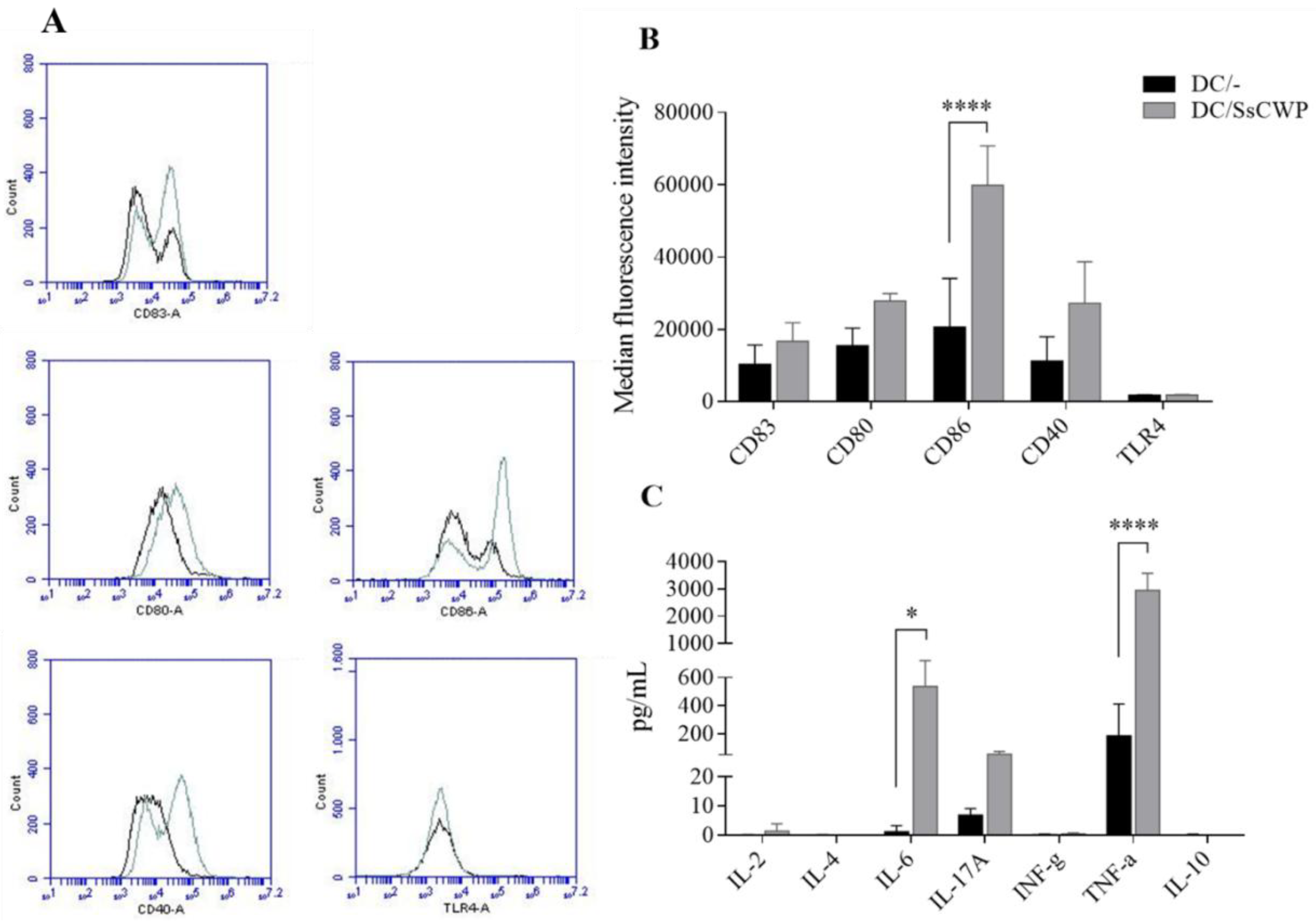{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Microorganism and Culture Conditions
2.3. Extraction of the SsCWP
2.4. BMDCs Generation
2.5. Flow Cytometry
2.6. Splenocytes
2.7. Cocultivation of BMDCs and Splenocytes
2.8. Cytokine Measurement
2.9. Statistical Analysis
3. Results
3.1. Activation and Cytokine Profile of SsCWP-Stimulated BMDCs
3.2. Cytokine Profile Induced by BMDCs Upon Cocultivation with Splenocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sasaki, A.A.; Fernandes, G.F.; Rodrigues, A.M.; Lima, F.M.; Marini, M.M.; Dos S Feitosa, L.; de Melo Teixeira, M.; Felipe, M.S.; da Silveira, J.F.; de Camargo, Z.P. Chromosomal polymorphism in the Sporothrix schenckii complex. PLoS ONE 2014, 9, e86819. [Google Scholar] [CrossRef] [PubMed]
- Gremião, I.D.; Menezes, R.C.; Schubach, T.M.; Figueiredo, A.B.; Cavalcanti, M.C.; Pereira, S.A. Feline sporotrichosis: Epidemiological and clinical aspects. Med. Mycol. 2015, 53, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Carlos, I.Z.; Batista-Duharte, A. Sporotrichosis; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–23. [Google Scholar]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Cassone, A. Fungal vaccines: Real progress from real challenges. Lancet Infect. Dis. 2008, 8, 114–124. [Google Scholar] [CrossRef]
- Edwards, J.E., Jr. Fungal cell wall vaccines: An update. J. Med. Microbiol. 2012, 61, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Portuondo, D.L.; Ferreira, L.S.; Urbaczek, A.C.; Batista-Duharte, A.; Carlos, I.Z. Adjuvants and delivery systems for antifungal vaccines: Current state and future developments. Med. Mycol. 2015, 53, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Alba-Fierro, C.A.; Pérez-Torres, A.; López-Romero, E.; Cuéllar-Cruz, M.; Ruiz-Baca, E. Cell wall proteins of Sporothrix schenckii as immunoprotective agents. Rev. Iberoam. Micol. 2014, 31, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Blander, J.M.; Medzhitov, R. Toll-dependent selection of microbial antigens for presentation by dendritic cells. Nature 2006, 440, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Benach, J.L.; Habicht, G.S.; Holbrook, T.W.; Cook, J.A. Glucan as an adjuvant for a murine Babesia microti immunization trial. Infect. Immun. 1982, 35, 947–951. [Google Scholar] [PubMed]
- LeibundGut-Landmann, S.; Gross, O.; Robinson, M.J.; Osorio, F.; Slack, E.C.; Tsoni, S.V.; Schweighoffer, E.; Tybulewicz, V.; Brown, G.D.; Ruland, J.; et al. Syk- and CARD9-dependent coupling of innate immunity to the induction of T helper cells that produce interleukin 17. Nat. Immunol. 2007, 8, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Liu, X.; Sun, Y.; Zhou, P.; Wang, Y.; Zhang, Y. Dendritic cell targeted vaccines: Recent progresses and challenges. Hum. Vaccin. Immunother. 2016, 12, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.; Ferreira, K.S.; Almeida, S.R.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Prophylactic and therapeutic vaccination using dendritic cells primed with peptide 10 derived from the 43-kilodalton glycoprotein of Paracoccidioides brasiliensis. Clin. Vaccine Immunol. 2012, 19, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Tacken, P.J.; Zeelenberg, I.S.; Cruz, L.J.; van Hout-Kuijer, M.A.; van de Glind, G.; Fokkink, R.G.; Lambeck, A.J.; Figdor, C.G. Targeted delivery of TLR ligands to human and mouse dendritic cells strongly enhances adjuvanticity. Blood 2011, 118, 6836–6844. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Liu, Y.J.; Kadowaki, N. Functional diversity and plasticity of human dendritic cell subsets. Int. J. Hematol. 2005, 81, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Bozza, S.; Montagnoli, C.; Gaziano, R.; Rossi, G.; Nkwanyuo, G.; Bellocchio, S.; Romani, L. Dendritic cell-based vaccination against opportunistic fungi. Vaccine 2004, 22, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, J.E.; Luft, V.D.; Amorim, J.; Magalhães, A.; Thomaz, L.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Immunization with P10 peptide increases specific immunity and protects immunosuppressed BALB/c mice infected with virulent yeasts of Paracoccidioides brasiliensis. Mycopathologia 2014, 178, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Verdan, F.F.; Faleiros, J.C.; Ferreira, L.S.; Monnazzi, L.G.; Maia, D.C.; Tansine, A.; Placeres, M.C.; Carlos, I.Z.; Santos-Junior, R.R. Dendritic cell are able to differentially recognize Sporothrix schenckii antigens and promote Th1/Th17 response in vitro. Immunobiology 2012, 217, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Portuondo, D.L.; Batista-Duharte, A.; Ferreira, L.S.; Martínez, D.T.; Polesi, M.C.; Duarte, R.A.; de Paula E Silva, A.C.; Marcos, C.M.; Almeida, A.M.; Carlos, I.Z. A cell wall protein-based vaccine candidate induce protective immune response against Sporothrix schenckii infection. Immunobiology 2016, 221, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Portuondo, D.L.; Batista-Duharte, A.; Ferreira, L.S.; de Andrade, C.R.; Quinello, C.; Téllez-Martínez, D.; de Aguiar Loesch, M.L.; Carlos, I.Z. Comparative efficacy and toxicity of two vaccine candidates against Sporothrix schenckii using either Montanide™ Pet Gel A or aluminum hydroxide adjuvants in mice. Vaccine 2017, 35, 4430–4436. [Google Scholar] [CrossRef] [PubMed]
- Batista-Duharte, A.; Téllez-Martínez, D.; Aparecida Jellmayer, J.; Leandro Portuondo Fuentes, D.; Campos Polesi, M.; Martins Baviera, A.; Zeppone Carlos, I. Repeated exposition to mercury (II) chloride enhances susceptibility to S. schenckii sensu stricto infection in mice. J. Fungi 2018, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.M.; Klein, B.S. Dendritic cells in antifungal immunity and vaccine design. Cell Host Microbe 2012, 11, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Van Stipdonk, M.J.; Lemmens, E.E.; Schoenberger, S.P. Naïve CTLs require a single brief period of antigenic stimulation for clonal expansion and differentiation. Nat. Immunol. 2001, 2, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Bacci, A.; Montagnoli, C.; Perruccio, K.; Bozza, S.; Gaziano, R.; Pitzurra, L.; Velardi, A.; d’Ostiani, C.F.; Cutler, J.E.; Romani, L. Dendritic cells pulsed with fungal RNA induce protective immunity to Candida albicans in hematopoietic transplantation. J. Immunol. 2002, 168, 2904–2913. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, G.; Nisini, R.; Chiani, P.; Mariotti, S.; Teloni, R.; Cassone, A.; Torosantucci, A. The interaction of human dendritic cells with yeast and germ-tube forms of Candida albicans leads to efficient fungal processing, dendritic cell maturation, and acquisition of a Th1 response-promoting function. J. Leukoc. Biol. 2004, 75, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Jellmayer, J.A.; Ferreira, L.S.; Manente, F.A.; Gonçalves, A.C.; Polesi, M.C.; Batista-Duharte, A.; Carlos, I.Z. Dectin-1 expression by macrophages and related antifungal mechanisms in a murine model of Sporothrix schenckii sensu stricto systemic infection. Microb. Pathog. 2017, 110, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martínez-Duncker, I.; Lópes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and sporothrix brasiliensis are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Uenotsuchi, T.; Takeuchi, S.; Matsuda, T.; Urabe, K.; Koga, T.; Uchi, H.; Nakahara, T.; Fukagawa, S.; Kawasaki, M.; Kajiwara, H.; et al. Differential induction of Th1-prone immunity by human dendritic cells activated with Sporothrix schenckii of cutaneous and visceral origins to determine their different virulence. Int. Immunol. 2006, 18, 1637–1646. [Google Scholar] [CrossRef] [PubMed]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Miranda, L.H.M.; Santiago, M.A.; Schubach, T.M.P.; Morgado, F.N.; Pereira, S.A.; Oliveira, R.V.C.; Conceição-Silva, F. Severe feline sporotrichosis associated with an increased population of CD8low cells and a decrease in CD4+ cells. Med. Mycol. 2016, 54, 29–39. [Google Scholar] [PubMed]
- Batista-Duharte, A.; Téllez-Martínez, D.; Andrade, R.C.; Portuondo, D.F.; Jellmayer, J.A.; Polesi, M.C.; Carlos, I.Z. Sporothrix brasiliensis induces a more severe disease associated with sustained Th17 and Treg cells responses than Sporothrix schenckii sensu stricto in mice. Fungal Biol. 2018, in press. [Google Scholar] [CrossRef]
- Kusuhara, M.; Qian, H.; Li, X.; Tsuruta, D.; Tsuchisaka, A.; Ishii, N.; Ohata, C.; Furumura, M.; Hashimoto, T. Mouse bone marrow-derived dendritic cells can phagocytize the Sporothrix schenckii, and mature and activate the immune response by secreting interleukin-12 and presenting antigens to T lymphocytes. J. Dermatol. 2014, 41, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Batista-Duharte, A.; Martínez, D.T.; Carlos, I.Z. Efficacy and safety of immunological adjuvants. Where is the cut-off? Biomed. Pharmacother. 2018, 105, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Batista-Duharte, A.; Téllez-Martínez, D.; Fuentes, D.L.P.; Carlos, I.Z. Molecular adjuvants that modulate regulatory T cell function in vaccination: A critical appraisal. Pharmacol. Res. 2018, 129, 237–250. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinello, C.; Souza Ferreira, L.; Picolli, I.; Loesch, M.L.; Portuondo, D.L.; Batista-Duharte, A.; Zeppone Carlos, I. Sporothrix schenckii Cell Wall Proteins-Stimulated BMDCs Are Able to Induce a Th1-Prone Cytokine Profile In Vitro. J. Fungi 2018, 4, 106. https://doi.org/10.3390/jof4030106
Quinello C, Souza Ferreira L, Picolli I, Loesch ML, Portuondo DL, Batista-Duharte A, Zeppone Carlos I. Sporothrix schenckii Cell Wall Proteins-Stimulated BMDCs Are Able to Induce a Th1-Prone Cytokine Profile In Vitro. Journal of Fungi. 2018; 4(3):106. https://doi.org/10.3390/jof4030106
Chicago/Turabian StyleQuinello, Camila, Lucas Souza Ferreira, Isabella Picolli, Maria Luiza Loesch, Deivys Leandro Portuondo, Alexander Batista-Duharte, and Iracilda Zeppone Carlos. 2018. "Sporothrix schenckii Cell Wall Proteins-Stimulated BMDCs Are Able to Induce a Th1-Prone Cytokine Profile In Vitro" Journal of Fungi 4, no. 3: 106. https://doi.org/10.3390/jof4030106
APA StyleQuinello, C., Souza Ferreira, L., Picolli, I., Loesch, M. L., Portuondo, D. L., Batista-Duharte, A., & Zeppone Carlos, I. (2018). Sporothrix schenckii Cell Wall Proteins-Stimulated BMDCs Are Able to Induce a Th1-Prone Cytokine Profile In Vitro. Journal of Fungi, 4(3), 106. https://doi.org/10.3390/jof4030106