Alternaria and Fusarium Fungi: Differences in Distribution and Spore Deposition in a Topographically Heterogeneous Wheat Field


Abstract
1. Introduction
2. Materials and Methods
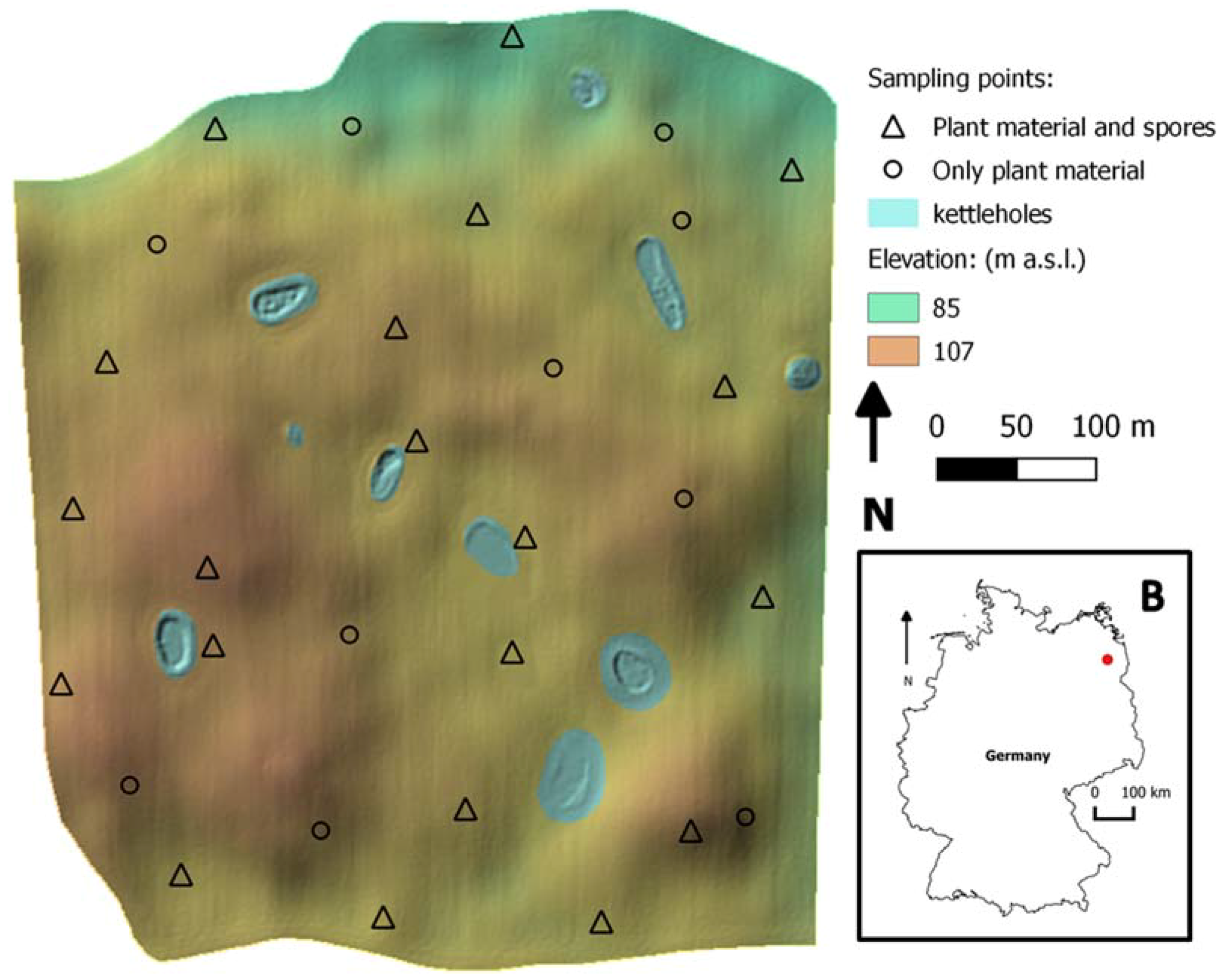
2.1. Field Study
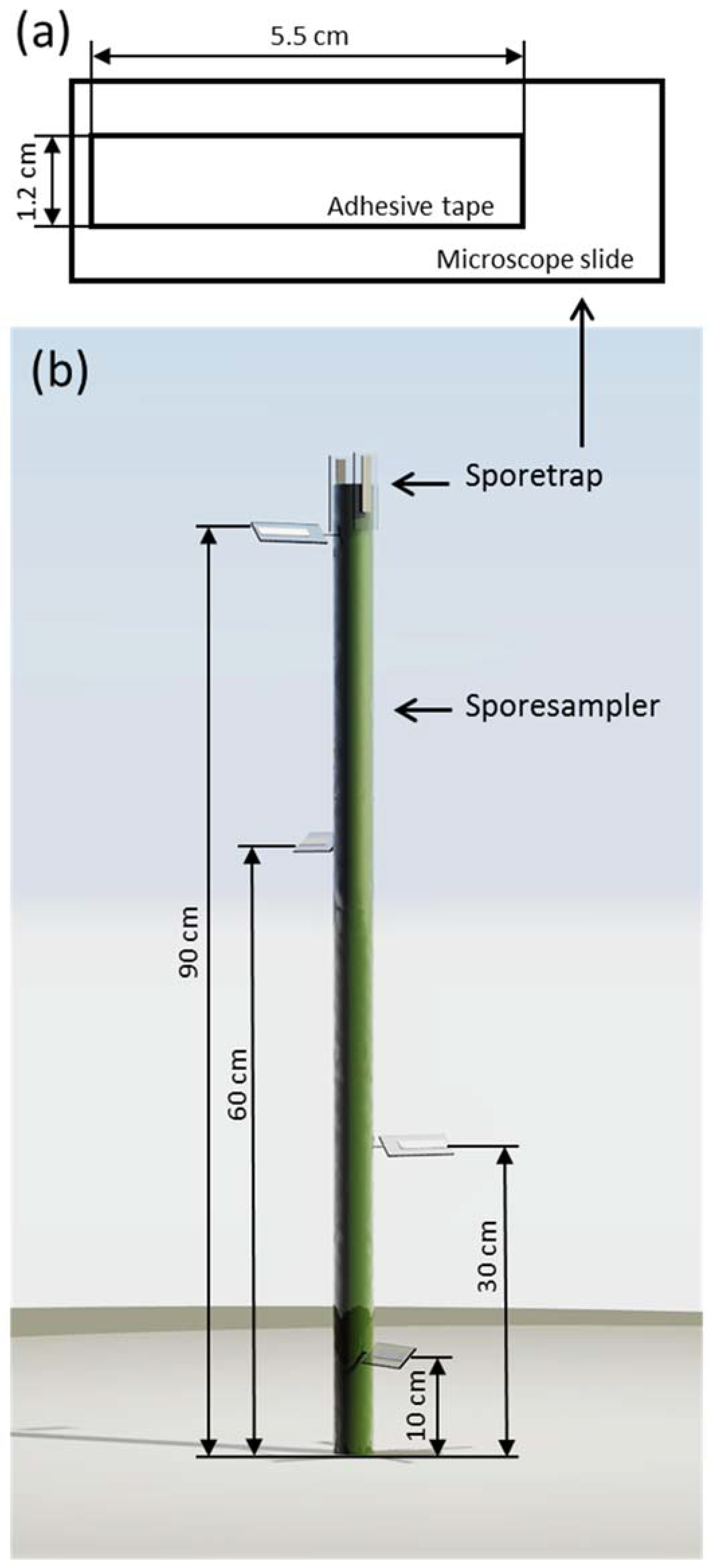
2.2. Spore Traps and Samplers
2.3. Spore Quantification
2.4. DNA Extraction
2.5. Fungal Quantification with qPCR
2.6. Statistical Analysis
3. Results
3.1. Microclimatic Measurements
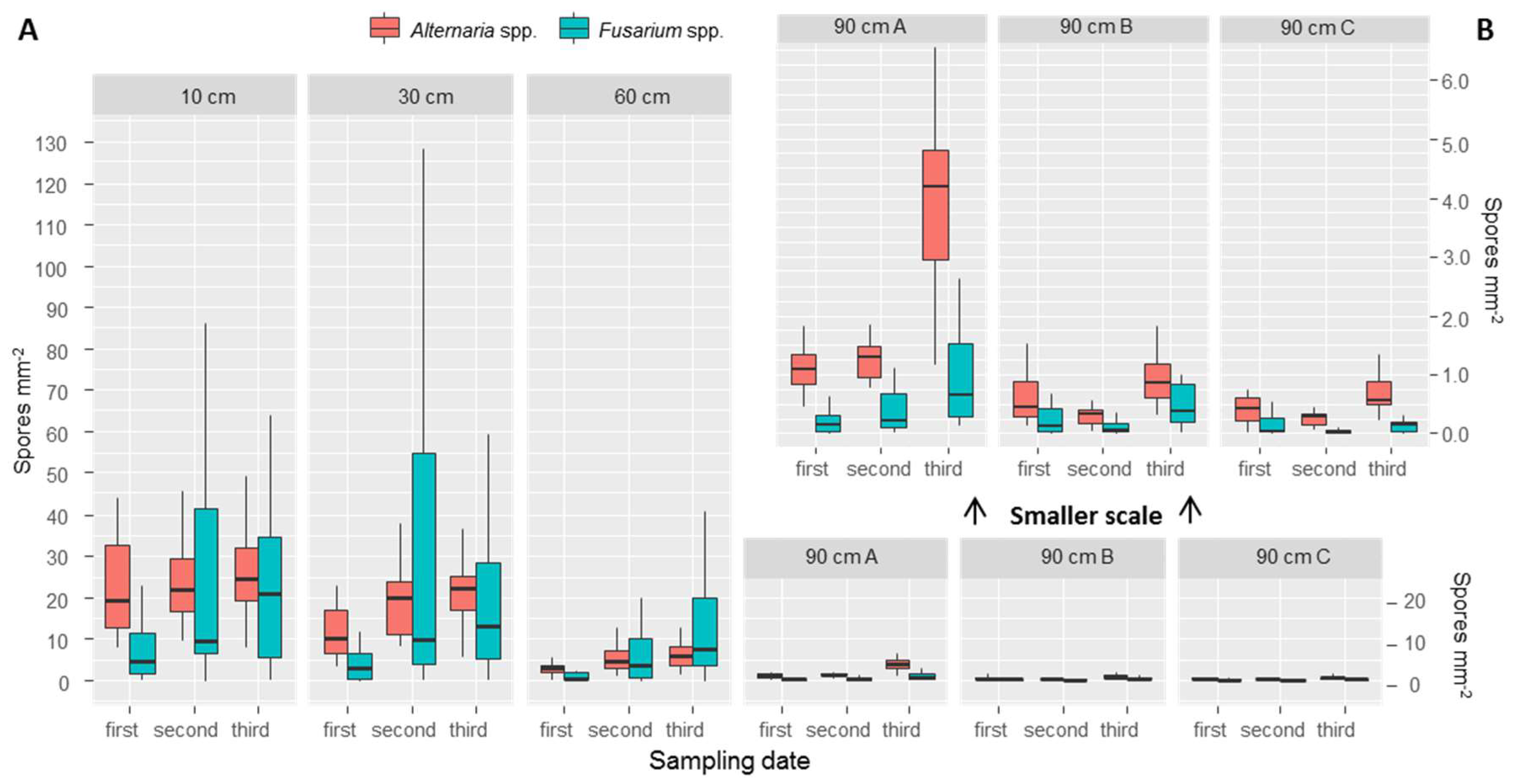
3.2. Spore Counts
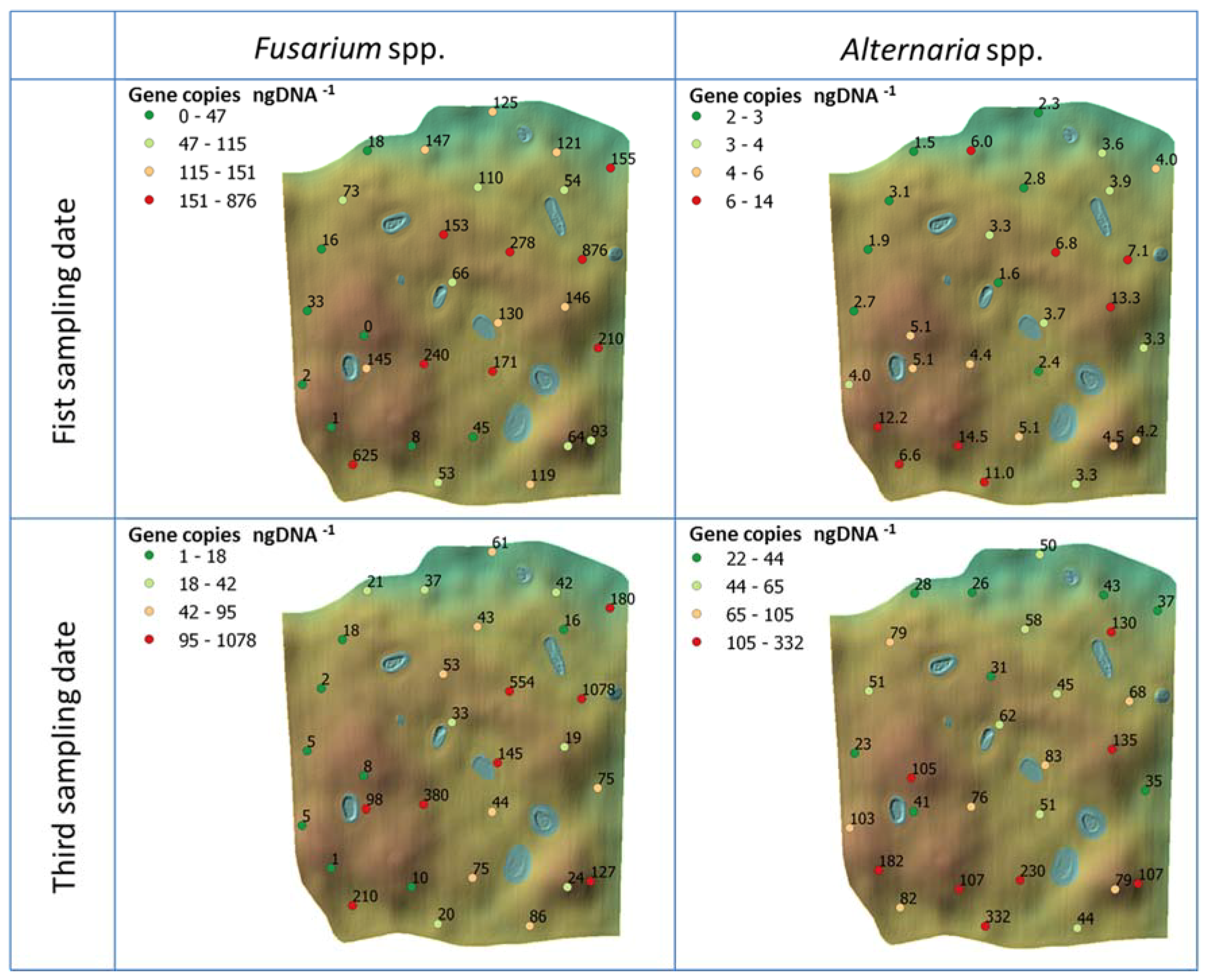
3.3. Results of qPCR Measurements
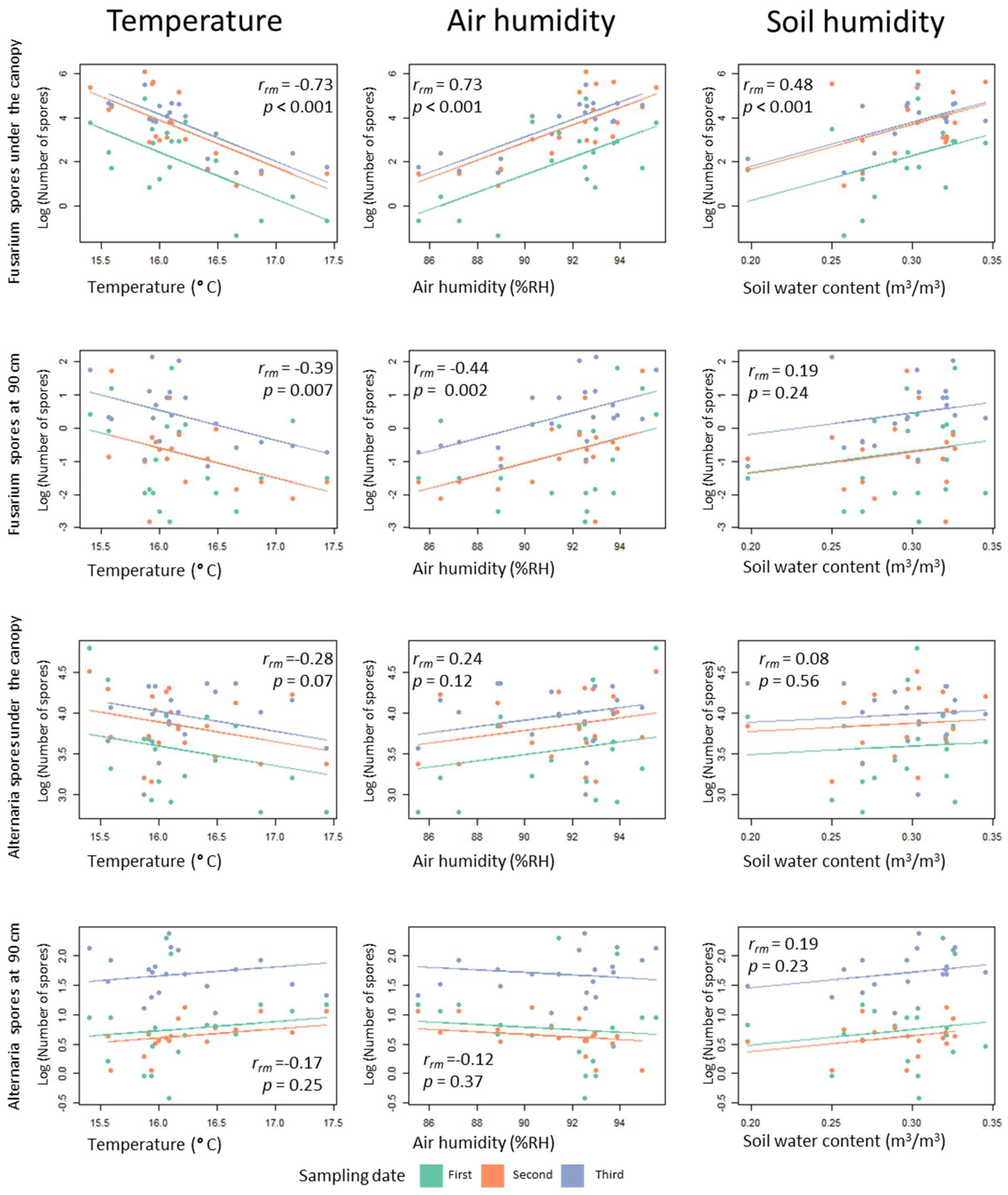
3.4. Relationship between Microclimatic Conditions and Spore Deposition
3.5. Relationship between Microclimatic Conditions and qPCR Results
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases-a field perspective. Mol. Plant Pathol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Luck, J.; Spackman, M.; Freeman, A.; TreBicki, P.; Griffiths, W.; Finlay, K.; Chakraborty, S. Climate change and diseases of food crops. Plant Pathol. 2011, 60, 113–121. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Savary, S.; Ficke, A.; Aubertot, J.N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Xu, X.; Nicholson, P. Community Ecology of Fungal Pathogens Causing Wheat Head Blight. Annu. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A.; Leslie, J.F. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Divers. 2011, 50, 135–144. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Pirgozliev, S.R.; Edwards, S.G.; Hare, M.C.; Jenkinson, P. Strategies for the control of Fusarium head blight in cereals. Eur. J. Plant Pathol. 2003, 109, 731–742. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium head blight of wheat and barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Xu, X. Effects of environmental conditions on the development of Fusarium ear blight. Eur. J. Plant Pathol. 2003, 109, 683–689. [Google Scholar] [CrossRef]
- Castiblanco, V.; Castillo, H.; Miedaner, T. Candidate Genes for Aggressiveness in a Natural Fusarium culmorum Population Greatly Differ between Wheat and Rye Head Blight. J. Fungi 2018, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Manstretta, V.; Rossi, V. Modelling the effect of weather on moisture fluctuations in maize stalk residues, an important inoculum source for plant diseases. Agric. For. Meteorol. 2015, 207, 83–93. [Google Scholar] [CrossRef]
- Maiorano, A.; Blandino, M.; Reyneri, A.; Vanara, F. Effects of maize residues on the Fusarium spp. infection and deoxynivalenol (DON) contamination of wheat grain. Crop Prot. 2008, 27, 182–188. [Google Scholar] [CrossRef]
- Pereyra, S.A.; Dill-Macky, R. Colonization of the Residues of Diverse Plant Species by Gibberella zeae and Their Contribution to Fusarium Head Blight Inoculum. Plant Dis. 2008, 92, 800–807. [Google Scholar] [CrossRef]
- Keller, M.D.; Thomason, W.E.; Schmale, D.G. The Spread of a Released Clone of Gibberella zeae from Different Amounts of Infested Corn Residue. Plant Dis. 2011, 95, 1458–1464. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Hoboken, NJ, USA, 2006; ISBN 9780470278376. [Google Scholar]
- Keller, M.D.; Bergstrom, G.C.; Shields, E.J. The aerobiology of Fusarium graminearum. Aerobiologia 2014, 30, 123–136. [Google Scholar] [CrossRef]
- Paul, P.A.; El-Allaf, S.M.; Lipps, P.E.; Madden, L. V Rain splash dispersal of Gibberella zeae within wheat canopies in Ohio. Phytopathology 2004, 94, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Manstretta, V.; Gourdain, E.; Rossi, V. Deposition patterns of Fusarium graminearum ascospores and conidia within a wheat canopy. Eur. J. Plant Pathol. 2015, 143, 873–880. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Patriarca, A.; Magan, N. Alternaria in Food: Ecophysiology, Mycotoxin Production and Toxicology. Mycobiology 2015, 43, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: An overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- Magan, N.; Cayley, G.R.; Lacey, J. Effect of Water Activity and Temperature on Mycotoxin Production by Alternaria alternata in Culture and on Wheat Grain. Appl. Environ. Microbiol. 1984, 47, 1113–1117. [Google Scholar] [PubMed]
- Perelló, A.; Moreno, M.; Sisterna, M. Alternaria infectoria species-group associated with black point of wheat in Argentina. Plant Pathol. 2008, 57, 379. [Google Scholar] [CrossRef]
- Andersen, B.; Sørensen, J.L.; Nielsen, K.F.; Gerrits van den Ende, B.; de Hoog, S. A polyphasic approach to the taxonomy of the Alternaria infectoria species-group. Fungal Genet. Biol. 2009, 46, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Maya-Manzano, J.M.; Fernández-Rodríguez, S.; Hernández-Trejo, F.; Díaz-Pérez, G.; Gonzalo-Garijo, Á.; Silva-Palacios, I.; Muñoz-Rodríguez, A.F.; Tormo-Molina, R. Seasonal Mediterranean pattern for airborne spores of Alternaria. Aerobiologia 2012, 28, 515–525. [Google Scholar] [CrossRef]
- Filali Ben Sidel, F.; Bouziane, H.; del Mar Trigo, M.; El Haskouri, F.; Bardei, F.; Redouane, A.; Kadiri, M.; Riadi, H.; Kazzaz, M. Airborne fungal spores of Alternaria, meteorological parameters and predicting variables. Int. J. Biometeorol. 2015, 59, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Skjøth, C.A.; Damialis, A.; Belmonte, J.; De Linares, C.; Fernández-Rodríguez, S.; Grinn-Gofroń, A.; Jędryczka, M.; Kasprzyk, I.; Magyar, D.; Myszkowska, D.; et al. Alternaria spores in the air across Europe: Abundance, seasonality and relationships with climate, meteorology and local environment. Aerobiologia 2016, 32, 3–22. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Justesen, A.F.; Knorr, K.; Wang, J.; Pinnschmidt, H.O. Fungal communities in wheat grain show significant co-existence patterns among species. Fungal Ecol. 2014, 11, 145–153. [Google Scholar] [CrossRef]
- Hertz, M.; Jensen, I.R.; Jensen, L.Ø.; Thomsen, S.N.; Winde, J.; Dueholm, M.S.; Sørensen, L.H.; Wollenberg, R.D.; Sørensen, H.O.; Sondergaard, T.E.; Sørensen, J.L. The fungal community changes over time in developing wheat heads. Int. J. Food Microbiol. 2016, 222, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.E.H.; Urban, K.; Köppen, R.; Siegel, D.; Korn, U.; Koch, M. Mycotoxins as antagonistic or supporting agents in the interaction between phytopathogenic Fusarium and Alternaria fungi. World Mycotoxin J. 2015, 8, 311–321. [Google Scholar] [CrossRef]
- Saß, V.; Milles, J.; Krämer, J.; Prange, A. Competitive interactions of Fusarium graminearum and Alternaria alternata in vitro in relation to deoxynivalenol and zearalenone production. J. Food Agric. Environ. 2007, 5, 257–261. [Google Scholar]
- Kosiak, B.; Torp, M.; Skjerve, E.; Andersen, B. Alternaria and Fusarium in Norwegian grains of reduced quality—a matched pair sample study. Int. J. Food Microbiol. 2004, 93, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Lacey, J.; Bateman, G.L.; Mirocha, C.J. Effects of infection time and moisture on development of ear blight and deoxynivalenol production by Fusarium spp. in wheat. Ann. Appl. Biol. 1999, 134, 277–283. [Google Scholar] [CrossRef]
- Magan, N.; Lacey, J. Effect of water activity, temperature and substrate on interactions between field and storage fungi. Trans. Br. Mycol. Soc. 1984, 82, 83–93. [Google Scholar] [CrossRef]
- Müller, M.E.H.; Koszinski, S.; Bangs, D.E.; Wehrhan, M.; Ulrich, A.; Verch, G.; Brenning, A. Crop biomass and humidity related factors reflect the spatial distribution of phytopathogenic Fusarium fungi and their mycotoxins in heterogeneous fields and landscapes. Precis. Agric. 2016, 1–23. [Google Scholar] [CrossRef]
- Federal Office of Plant Varieties (Bundessortenamt) Beschreibende Sortenliste Getreide, Mais, Öl- und Faserpflanzen, Leguminosen, Rüben, Zwischenfrüchte. Hannover, Germany, 2017. Available online: https://www.bundessortenamt.de/internet30/fileadmin/Files/PDF/bsl_getreide_2017.pdf (accessed on 18 May 2018).
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed]
- Kahl, S.M.; Ulrich, A.; Kirichenko, A.A.; Müller, M.E.H. Phenotypic and phylogenetic segregation of Alternaria infectoria from small-spored Alternaria species isolated from wheat in Germany and Russia. J. Appl. Microbiol. 2015, 119, 1637–1650. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.X.; Pryor, B.; Peever, T.; Lawrence, C.B. The Alternaria genomes database: A comprehensive resource for a fungal genus comprised of saprophytes, plant pathogens, and allergenic species. BMC Genom. 2015, 16, 239. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, I.; Gautheron, N.; Kolseth, A.; Steinberg, C.; Persson, P.; Friberg, H. Genus-Specific Primers for Study of Fusarium Communities in Field. Appl. Environ. Microbil. 2016, 82, 491–501. [Google Scholar] [CrossRef] [PubMed]
- King, R.; Urban, M.; Hammond-Kosack, M.C.U.; Hassani-Pak, K.; Hammond-Kosack, K.E. The completed genome sequence of the pathogenic ascomycete fungus Fusarium graminearum. BMC Genom. 2015, 16, 1–21. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 18 May 2018).
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research. 2017. Available online: https://cran.r-project.org/package=psych (accessed on 18 May 2018).
- Bakdash, J.Z.; Marusich, L.R. Repeated measures correlation. Front. Psychol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, S. lm.beta: Add Standardized Regression Coefficients to lm-Objects. 2014. Available online: https://CRAN.R-project.org/package=lm.beta (accessed on 18 May 2018).
- Stack, R.W. A comparison of the inoculum potential of ascospores and conidia of Gibberella zeae. Can. J. Plant Pathol. 1989, 11, 137–142. [Google Scholar] [CrossRef]
- Paulitz, T.C. Diurnal release of ascospores by Gibberella zeae in inoculated wheat plots. Plant Dis. 1996, 80, 674–678. [Google Scholar] [CrossRef]
- Rossi, V.; Languasco, L.; Pattori, E.; Giosuè, S. Dynamics of airborne Fusarium macroconidia in wheat fields naturally affected by head blight. J. Plant Pathol. 2002, 84, 53–64. [Google Scholar] [CrossRef]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- De Aldana, B.R.V.; Bills, G.; Zabalgogeazcoa, I. Are endophytes an important link between airborne spores and allergen exposure? Fungal Divers. 2013, 60, 33–42. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Logrieco, A. Mycotoxin Reduction in Grain Chain. Mycotoxin Reduct. Grain Chain 2014, 376. [Google Scholar] [CrossRef]
- Andersen, B.; Thrane, U.; Svendsen, A.; Rasmussen, L.A. Associated field mycobiota on malt barley. Can. J. Bot. 1996, 74, 854–858. [Google Scholar] [CrossRef]
- Andersen, B.; Thrane, U. Differentiation of Altemaria infectoria and Alternaria alternata based on morphology, metabolite profiles, and cultural characteristics. Can. J. Microbiol. 1996, 42, 685–689. [Google Scholar] [CrossRef]
- Ofek-Lalzar, M.; Gur, Y.; Ben-Moshe, S.; Sharon, O.; Kosman, E.; Mochli, E.; Sharon, A. Diversity of fungal endophytes in recent and ancient wheat ancestors triticum dicoccoides and aegilops sharonensis. FEMS Microbiol. Ecol. 2016, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Humidity | Soil Humidity | Plant Height | |
|---|---|---|---|
| Temperature | 0.91133 *** | −0.48714 * | −0.89771 *** |
| Air humidity | 0.599343 *** | 0.801085 *** | |
| Plant height | 0.385648 * |
| Fusariumspp. (Spores mm−2) | |||||||||
| First Sampling Date | Second Sampling Date | Third Sampling Date | |||||||
| height (cm) | AVG | STD | CV | AVG | STD | CV | AVG | STD | CV |
| 10 | 13.22 | 28.23 | 2.13 | 34.56 | 57.91 | 1.68 | 31.93 | 45.83 | 1.44 |
| 30 | 3.55 | 3.78 | 1.06 | 46.29 | 69.53 | 1.50 | 22.31 | 25.04 | 1.12 |
| 60 | 3.40 | 9.14 | 2.69 | 8.03 | 10.49 | 1.31 | 17.53 | 24.16 | 1.38 |
| 90 A | 0.81 | 2.00 | 2.47 | 0.64 | 1.12 | 1.74 | 1.46 | 1.98 | 1.36 |
| 90 B | 0.31 | 0.49 | 1.59 | 0.14 | 0.16 | 1.16 | 0.60 | 0.63 | 1.05 |
| 90 C | 0.20 | 0.29 | 1.45 | 0.06 | 0.10 | 1.80 | 0.18 | 0.21 | 1.16 |
| Alternariaspp. (Spores mm−2) | |||||||||
| First Sampling Date | Second Sampling Date | Third Sampling Date | |||||||
| height (cm) | AVG | STD | CV | AVG | STD | CV | AVG | STD | CV |
| 10 | 22.17 | 12.06 | 0.54 | 23.96 | 10.46 | 0.44 | 25.81 | 10.72 | 0.41 |
| 30 | 15.11 | 15.92 | 1.05 | 21.06 | 11.99 | 0.57 | 24.91 | 12.19 | 0.50 |
| 60 | 2.75 | 1.47 | 0.53 | 5.13 | 3.35 | 0.65 | 6.13 | 3.05 | 0.50 |
| 90 A | 1.34 | 1.04 | 0.78 | 1.30 | 0.33 | 0.25 | 3.98 | 1.51 | 0.38 |
| 90 B | 0.86 | 1.27 | 1.48 | 0.33 | 0.20 | 0.59 | 1.00 | 0.52 | 0.52 |
| 90 C | 0.46 | 0.36 | 0.78 | 0.31 | 0.25 | 0.79 | 0.77 | 0.63 | 0.82 |
| First Sampling Date | Second Sampling Date | Third Sampling Date | |
|---|---|---|---|
| Alternaria | 0.08152 | 0.9409 | 0.074 |
| Fusarium | 0.2904 | 0.04276 | 0.001962 |
| Fungus | Sampling Date | Temperature | Humidity | Soil Humidity |
|---|---|---|---|---|
| Fusarium | First | −0.41675 * | 0.42266 * | 0.579639 ** |
| Fusarium | Third | −0.36059 | 0.345813 | 0.385878 |
| Alternaria | First | 0.608374 ** | −0.60788 ** | −0.35742 |
| Alternaria | Third | 0.482266 * | −0.50049 * | −0.39409 |
| Response Variable: Gene Copies Number | Sampling Date | Adjusted r2 | β Values (Significance Codes) | ||
|---|---|---|---|---|---|
| Temperature | Soil humidity | Other Genus’ Abundance | |||
| Fusarium spp. | First | 0.45 *** | −0.47 * | 0.54 ** | 0.40 |
| Third | 0.16 | −0.25 | 0.38 | 0.12 | |
| Alternaria spp. | First | 0.5 *** | 0.75 *** | −0.21 | 0.36 |
| Third | 0.22 * | 0.42 * | −0.25 | 0.11 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiro, G.; Verch, G.; Grimm, V.; Müller, M.E.H. Alternaria and Fusarium Fungi: Differences in Distribution and Spore Deposition in a Topographically Heterogeneous Wheat Field. J. Fungi 2018, 4, 63. https://doi.org/10.3390/jof4020063
Schiro G, Verch G, Grimm V, Müller MEH. Alternaria and Fusarium Fungi: Differences in Distribution and Spore Deposition in a Topographically Heterogeneous Wheat Field. Journal of Fungi. 2018; 4(2):63. https://doi.org/10.3390/jof4020063
Chicago/Turabian StyleSchiro, Gabriele, Gernot Verch, Volker Grimm, and Marina E. H. Müller. 2018. "Alternaria and Fusarium Fungi: Differences in Distribution and Spore Deposition in a Topographically Heterogeneous Wheat Field" Journal of Fungi 4, no. 2: 63. https://doi.org/10.3390/jof4020063
APA StyleSchiro, G., Verch, G., Grimm, V., & Müller, M. E. H. (2018). Alternaria and Fusarium Fungi: Differences in Distribution and Spore Deposition in a Topographically Heterogeneous Wheat Field. Journal of Fungi, 4(2), 63. https://doi.org/10.3390/jof4020063