Endophytic Fungi in Species of Artemisia


Abstract
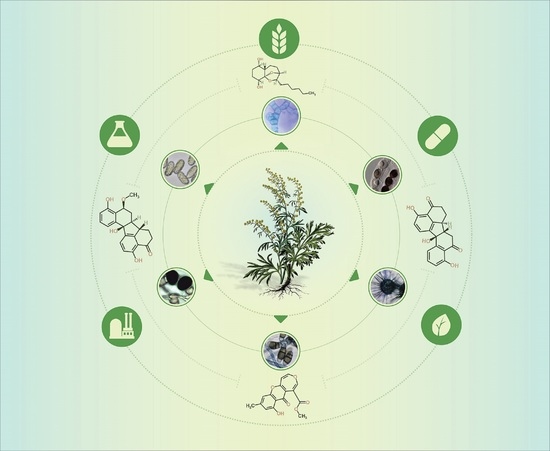
1. State of the Art
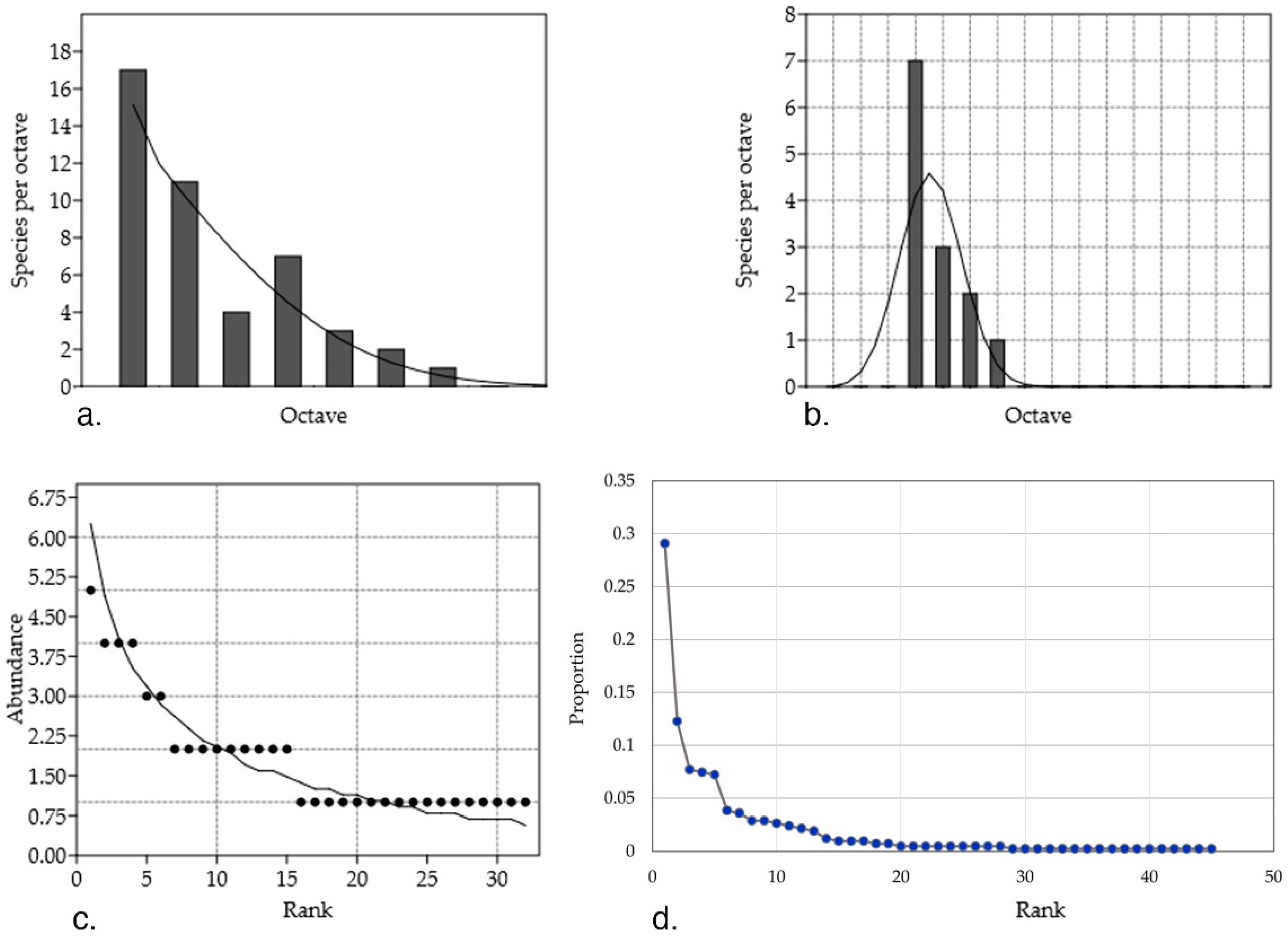
2. Artemisia Fungal Endophyte Identification and Diversity
3. Biological Activity of Artemisia Endophytes
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Haider, F.; Dwivedi, P.D.; Naqvi, A.A.; Bagchi, G.D. Essential oil composition of Artemisia vulgaris harvested at different growth periods under indo-gangetic plain conditions. J. Essent. Oil Res. 2003, 15, 376–378. [Google Scholar] [CrossRef]
- Judžentien, A.; Buzelyte, J. Chemical composition of essential oils of Artemisia vulgaris L. (mugwort) from North Lithuania. Chemija 2006, 17, 12–15. [Google Scholar]
- Blagojević, P.; Radulović, N.; Palić, R.; Stojanović, G. Chemical composition of the essential oils of Serbian wild-growing Artemisia absinthium and Artemisia vulgaris. J. Agric. Food Chem. 2006, 54, 4780–4789. [Google Scholar] [CrossRef] [PubMed]
- Abiri, R.; Silva, A.L.M.; de Mesquita, L.S.S.; de Mesquita, J.W.C.; Atabaki, N.; de Almeida, E.B.; Shaharuddin, S.M. Towards a better understanding of Artemisia vulgaris: Botany, phytochemistry, pharmacological and biotechnological potential. Food Res. Int. 2018. [Google Scholar] [CrossRef]
- Koul, B.; Taak, P. The Artemisia Genus: A Review on Traditional Uses, Phytochemical Constituents, Pharmacological Properties and Germplasm Conservation. J. Glycom. Lipidom. 2018, 7, 1–7. [Google Scholar] [CrossRef]
- Pandey, A.K.; Singh, P. The Genus Artemisia: A 2012–2017 Literature Review on Chemical Composition, Antimicrobial, Insecticidal and Antioxidant Activities of Essential Oils. Medicines 2017, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Tabari, M.A.; Youssefi, M.R.; Benelli, G. Eco-friendly control of the poultry red mite, Dermanyssus gallinae (Dermanyssidae), using the α-thujone-rich essential oil of Artemisia sieberi (Asteraceae): Toxic and repellent potential. Parasitol. Res. 2017, 116, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zou, K.; Zhang, W.; Guo, S.; Liu, H.; Sun, J.; Li, J.; Huang, D.; Wu, Y.; Du, S.; et al. Efficacy of Compounds Isolated from the Essential Oil of Artemisia lavandulaefolia in Control of the Cigarette Beetle, Lasioderma serricorne. Molecules 2018, 23, 343. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini-Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K. Anti-inflammatory effects of eriodictyol in lipopolysaccharidestimulated raw 264.7 murine macrophages. Arch. Pharm. Res. 2011, 34, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Bucolo, C.; Leggio, G.M.; Drago, F.; Salomone, S. Eriodictyol prevents early retinal and plasma abnormalities in streptozotocin-induced diabetic rats. Biochem. Pharmacol. 2012, 84, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Chung, H.Y.; Maier, C.G.; Wood, A.R.; Dixon, R.; Mabry, T. Estrogenic flavonoids from Artemisia vulgaris L. J. Agric. Food Chem. 1998, 46, 3325–3329. [Google Scholar] [CrossRef]
- Ikram, N.K.B.K.; Simonsen, H.T. A Review of Biotechnological Artemisinin Production in Plants. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.B.S.; Meurer, Y.S.R.; Oliveira, M.G.; Medeiros, W.M.T.Q.; Silva, F.O.N.; Brito, A.C.F.; Pontes, D.L.; Andrade-Neto, V. Comparative study on the antioxidant and anti-Toxoplasma activities of vanillin and its resorcinarene derivative. Molecules 2014, 19, 5898–5912. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.T.; Alshehri, M.A.; Panneerselvam, C.; Murugan, K.; Trivedi, S.; Mahyoub, J.A.; Hassan, M.M.; Maggi, F.; Sut, S.; Dall’Aqua, S.; et al. The desert wormwood (Artemisia herba-alba)—From Arabian folk medicine to a source of green and effective nanoinsecticides against mosquito vectors. J. Photochem. Photobiol. B Biol. 2018, 180, 225–234. [Google Scholar] [CrossRef] [PubMed]
- D’Addabbo, T.; Argentieri, M.P.; Radicci, V.; Grassi, F.; Avato, P. Artemisia annua compounds have potential to manage root-knot and potato cyst nematodes. Ind. Crops Prod. 2017, 108, 195–200. [Google Scholar] [CrossRef]
- Julio, L.F.; González-Coloma, A.; Burillo, J.; Diaz, C.E.; Andrés, M.F. Nematicidal activity of the hydrolate byproduct from the semi industrial vapor pressure extraction of domesticated Artemisia absinthium against Meloidogyne javanica. Crop Prot. 2017, 94, 33–37. [Google Scholar] [CrossRef]
- Chengala, L. Botanical pesticides—A major alternative to chemical pesticides: A review. Int. J. Life Sci. 2017, 5, 722–729. [Google Scholar]
- Esmaeily, M.; Bandani, A.; Zibaee, I.; Sharifian, I.; Zare, S. Sublethal effects of Artemisia annua L. and Rosmarinus officinalis L. essential oils on life table parameters of Tetranychus urticae (Acari: Tetranychidae). Persian J. Acarol. 2017, 6, 39–52. [Google Scholar]
- Sainz, P.; Cruz-Estrada, Á.; Díaz, C.E.; González-Coloma, A. The genus Artemisia: Distribution and phytochemistry in the Iberian Peninsula and the Canary and Balearic Islands. Phytochem. Rev. 2017, 16, 1023–1043. [Google Scholar] [CrossRef]
- Royal Botanic Gardens K and MBG. The Plant List 2010. Available online: http://www.theplantlist.org/tpl/search?q=artemisia (accessed on 3 February 2017).
- Huang, W.Y.; Cai, Y.Z.; Surveswaran, S.; Hyde, K.D.; Corke, H.; Sun, M. Molecular phylogenetic identification of endophytic fungi isolated from three Artemisia species. Fungal Divers. 2009, 36, 69–88. [Google Scholar]
- Cosoveanu, A.; Hernandez, M.; Iacomi-Vasilescu, B.; Zhang, X.; Shu, S.; Wang, M.; Cabrera, R. Fungi as endophytes in Chinese Artemisia spp.: Juxtaposed elements of phylogeny, diversity and bioactivity. Mycosphere 2016, 7, 102–117. [Google Scholar] [CrossRef]
- Cosoveanu, A.; Rodriguez Sabina, S.; Cabrera, R. Fungi as endophytes in Artemisia thuscula: Juxtaposed elements of diversity and phylogeny. J. Fungi 2018, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.L.; Chen, Y.C.; Ma, X.J. Symbiotic fungi in roots of Artemisia annua with special reference to endophytic colonizers. Plant Biosyst. 2011, 145, 495–502. [Google Scholar] [CrossRef]
- Ofek-lalzar, M.; Gur, Y.; Ben-Moshe, S.; Sharon, O.; Kosman, E.; Mochli, E.; Sharon, A. Diversity of fungal endophytes in recent and ancient wheat ancestors of Triticum dicoccoides and Aegilops sharonensis. FEMS Microbiol. Ecol. Adv. 2016, 92, 1–26. [Google Scholar] [CrossRef]
- Kernaghan, G.; Mayerhofer, M.; Griffin, A. Fungal endophytes of wild and hybrid Vitis leaves and their potential for vineyard biocontrol. Can. J. Microbiol. 2017, 63, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.M.C.; Hyde, K.D.; Corlett, R.T. Diversity of fungi on wild fruits in Hong Kong. Fungal Divers. 2003, 14, 165–185. [Google Scholar]
- Qian, Y.; Kang, J.; Geng, K.; Wang, L.; Lei, B. Endophytic fungi from Artemisia argyi Levl. et Vant. and their bioactivity. Chiang Mai J. Sci. 2014, 41, 910–921. [Google Scholar]
- Myrchiang, P.; Dkhar, M.S.; Devi, H.R. Studies on endophytic fungi associated with medicinally important aromatic plant Artemisia nilagirica (C.B. Clarke) Pamp. and their antagonistic activity against Phytophthora infestans. J. Adv. Lab. Res. Biol. 2014, 5, 112–119. [Google Scholar]
- Cosoveanu, A.; Da Silva, E.; Gimenez Marino, C.; Nunez Trujillo, G.; Gonzalez Coloma, A.; Frias Viera, I.; Cabrera, R. Artemisia thuscula Cav.: Antibacterial, antifungal activity of the plant extracts and associated endophytes. J. Hortic. For. Biotechnol. 2012, 16, 87–90. [Google Scholar]
- Gautam, A.K. Diversity of fungal endophytes in some medicinal plants of Himachal Pradesh, India. Arch. Phytopathol. Plant Prot. 2014, 47, 537–544. [Google Scholar] [CrossRef]
- Chowdhary, K.; Kaushik, N. Fungal endophyte diversity and bioactivity in the Indian medicinal plant Ocimum sanctum Linn. PLoS ONE 2015, 10, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Zhou, Y.; Zhou, X.; Xia, X.; Wei, Y.; He, L.; Tang, H.; Yu, L. Diversity and bioactive potential of culturable fungal endophytes of Dysosma versipellis; a rare medicinal plant endemic to China. Sci. Rep. 2018, 8, 5929. [Google Scholar] [CrossRef] [PubMed]
- Nalini, M.S.; Sunayana, N.; Prakash, H.S. Endophytic Fungal Diversity in Medicinal Plants of Western Ghats, India. Int. J. Biodivers. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Sánchez Márquez, S.; Bills, G.F.; Herrero, N.; Zabalgogeazcoa, Í. Non-systemic fungal endophytes of grasses. Fungal Ecol. 2012, 5, 289–297. [Google Scholar] [CrossRef]
- Banerjee, D. Endophytic fungal diversity in tropical and subtropical plants. Res. J. Microbiol. 2011, 6, 54–62. [Google Scholar] [CrossRef]
- Casieri, L.; Hofstetter, V.; Viret, O.; Gindro, K. Fungal communities in the wood of different cultivars of young Vitis vinifera plants. Phytopathol. Mediterr. 2009, 48, 73–83. [Google Scholar] [CrossRef]
- Qian, Y.; Kang, J.; Lei, B.; Wang, L.; Huang, Y. Screening and taxonomic identification of endophytic fungi with antitumor and antioxidant activities from Artemisia lactiflora. China J. Chin. Mater. Med. 2014, 39, 438–441. [Google Scholar]
- Lu, H.; Zou, W.X.; Meng, J.C.; Hu, J.; Tan, R.X. New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Sci. 2000, 151, 67–73. [Google Scholar] [CrossRef]
- Zhang, H.; Bai, X.; Wu, B. Evaluation of antimicrobial activities of extracts of endophytic fungi from Artemisia annua. Bangladesh J. Pharmacol. 2012, 7, 120–123. [Google Scholar] [CrossRef][Green Version]
- Gu, W.; Ge, H.M.; Song, Y.C.; Ding, H.; Zhu, H.L.; Zhao, X.A.; Tan, R.X. Cytotoxic benzo[j]fluoranthene metabolites from Hypoxylon truncatum IFB-18, an endophyte of Artemisia annua. J. Nat. Prod. 2007, 70, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Liu, C.H.; Zou, W.X.; Tan, R.X. Leptosphaerone, a metabolite with a novel skeleton from Leptosphaeria sp. IV403, an endophytic fungus in Artemisia annua. Helv. Chim. Acta 2002, 85, 2662–2667. [Google Scholar] [CrossRef]
- Wilco, E.P.; Verberk, E.P. Explaining general patterns in species abundance and distributions. Nat. Educ. Knowl. 2011, 3, 38. [Google Scholar]
- Galand, P.E.; Casamayor, E.O.; Kirchman, D.L.; Lovejoy, C. Ecology of the rare microbial biosphere of the Arctic Ocean. Proc. Natl. Acad. Sci. USA 2009, 106, 22427–22432. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [PubMed]
- Dastogeer, K.M.G.; Li, H.; Sivasithamparam, K.; Jones, M.G.K.; Wylie, S.J. Host Specificity of Endophytic Mycobiota of Wild Nicotiana Plants from Arid Regions of Northern Australia. Microb. Ecol. 2018, 75, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ding, Q.; Hyde, K.D.; Guo, L.D. Community structure and preference of endophytic fungi of three woody plants in a mixed forest. Fungal Ecol. 2012, 5, 624–632. [Google Scholar] [CrossRef]
- Zhou, D.; Hyde, K.D. Host specificity, hos exclusivity and host-recurrence in saprobic fungi. Mycol. Res. 2001, 105, 1449–1457. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Li, S.; Shi, D.H.; Song, Y.C.; Tan, R.X. Chemical constituents of liquid culture of endophyte IFB-E012 in Artemisia annua. Chin. J. Nat. Med. 2009, 7, 354–356. [Google Scholar] [CrossRef]
- Liu, C.H.; Zou, W.X.; Lu, H.; Tan, R.X. Antifungal activity of Artemisia annua endophyte cultures against phytopathogenic fungi. J. Biotechnol. 2001, 88, 277–282. [Google Scholar] [CrossRef]
- Wang, J.W.; Zhang, Z.; Tan, R.X. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Wang, J.W.; Zheng, L.P.; Tan, R.X. The preparation of an elicitor from a fungal endophyte to enhance artemisinin production in hairy root culture of Artemisia annua L. Chin. J. Biotechnol. 2006, 22, 829–834. [Google Scholar]
- Liu, J.Y.; Liu, C.H.; Zou, W.X.; Tan, R.X. Leptosphaeric acid, a metabolite with a novel carbon skeleton from Leptosphaeria sp. IV403, an endophytic fungus in Artemisia annua. Helv. Chim. Acta 2003, 86, 657–660. [Google Scholar] [CrossRef]
- Arora, M.; Saxena, P.; Choudhary, D.K.; Abdin, M.Z.; Varma, A. Dual symbiosis between Piriformospora indica and Azotobacter chroococcum enhances the artemisinin content in Artemisia annua L. World J. Microbiol. Biotechnol. 2016, 32, 19. [Google Scholar] [CrossRef] [PubMed]
- Baishya, D.; Deka, P.; Kalita, M.C. In vitro co-cultivation of Piriformospora indica filtrate for improve biomass productivity in Artemisia annua (L.). Symbiosis 2015, 66, 37–46. [Google Scholar] [CrossRef]
- Jogawat, A.; Saha, S.; Bakshi, M.; Dayaman, V.; Kumar, M.; Dua, M.; Varma, M.; Oelmuller, R.; Tuteja, N.; Johri, A.K. Piriformospora indica rescues growth diminution of rice seedlings during high salt stress. Plant Signal Behav. 2013, 8, e26891. [Google Scholar] [CrossRef]
- Hussain, M.; Mahajan, V.; Rather, I.A.; Awasthi, P.; Chouhan, R.; Dutt, P.; Sharma, Y.P.; Bedi, Y.S.; Gandhi, S.G. Isolation and identification of growth promoting endophytic fungi from Artemisia annua L. and its effects on artemisinin content. Trends Phytochem. Res. 2017, 1, 207–214. [Google Scholar]
- Jessing, K.K.; Duke, S.O.; Cedergreeen, N. Potential Ecological Roles of Artemisinin Produced by Artemisia annua L. J. Chem. Ecol. 2014, 40, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Das, A.K. Anticancer effect of Antimalarial artemisinin compounds. Ann. Med. Health Sci. Res. 2015, 5, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Xia, Z.H.; Tan, R.X. Elicitation on Artemisinin Biosynthesis in Artemisia annua Hairy Roots by the Oligosaccharide extract from the Endophytic Colletotrichum sp. B501. Acta Bot. Sin. 2002, 44, 1233–1238. [Google Scholar]
- Zheng, L.P.; Tian, H.; Yuan, Y.F.; Wang, J.W. The influence of endophytic Penicillium oxalicum B4 on growth and artemisinin biosynthesis of in vitro propagated plantlets of Artemisia annua L. Plant Growth Regul. 2016, 80, 93–102. [Google Scholar] [CrossRef]
- Zou, W.X.; Meng, J.C.; Lu, H.; Chen, G.X.; Shi, G.X.; Zhang, T.Y.; Tan, R.X. Metabolites of Colletotrichum gloeosporioides, an endophytic fungus in Artemisia mongolica. J. Nat. Prod. 2000, 63, 1529–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Ying, C.; Tang, Y.F. Four ardeemin analogs from endophytic Aspergillus fumigatus SPS-02 and their reversal effects on multidrug-resistant tumor cells. Chem. Biodivers. 2014, 11, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Purwantini, I.; Wahyono, M.; Ratna, A. Isolation of endophytic fungi from Artemisia annua, L and identification of their antimicrobial compound using bioautography method. Int. J. Phrmacy Pharm. Sci. 2015, 7, 95–99. [Google Scholar]
- Hsieh, P.W.; Hsu, L.C.; Lai, C.H.; Wu, C.C.; Hwang, T.L.; Lin, Y.K.; Wu, Y.C. Evaluation of the bioactivities of extracts of endophytes isolated from Taiwanese herbal plants. World J. Microbiol. Biotechnol. 2009, 25, 1461–1469. [Google Scholar] [CrossRef]
- Shi, X.S.; Wang, D.J.; Li, X.M.; Li, H.L.; Meng, L.H.; Li, X.; Pi, Y.; Zhou, X.W.; Wang, B.G. Antimicrobial polyketides from Trichoderma koningiopsis QA-3, an endophytic fungus obtained from the medicinal plant Artemisia argyi. RSC Adv. 2017, 7, 51335–51342. [Google Scholar] [CrossRef]
- Shen, L.; Zhu, L.; Tan, Q.; Wan, D.; Xie, J.; Peng, J. New cytotoxic trichothecene macrolide epimers from endophytic Myrothecium roridum IFB-E012. J. Antibiot. 2016, 69, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Jiao, R.H.; Ye, Y.H.; Wang, X.T.; Xu, C.; Song, Y.C.; Zhu, H.L.; Tan, R.X. Absolute configuration of new cytotoxic and other bioactive trichothecene macrolides. Chem. A Eur. J. 2006, 12, 5596–5602. [Google Scholar] [CrossRef] [PubMed]
- Quang, D.N.; Hashimoto, T.; Nomura, Y.; Wollweber, H.; Hellwig, V.; Fournier, J.; Staedler, M.; Asakawa, Y. Cohaerins A and B, azaphilones from the fungus Hypoxylon cohaerens, and comparison of HPLC-based metabolite profiles in Hypoxylon sect. Annulata. Phytochemistry 2005, 66, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Cai, Y.Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Noumeur, S.R.; Mancini, V.; Romanazzi, G. Activity of endophytic fungi from Artemisia absinthium on Botrytis cinerea. Acta Hortic. 2016, 1144, 101–104. [Google Scholar] [CrossRef]
- Tian, H.; Ma, Y.J.; Li, W.Y.; Wang, J.W. Efficient degradation of triclosan by an endophytic fungus Penicillium oxalicum B4. Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Zhang, J.; Hu, S.; Zhang, Z.J.; Zhu, C.J.; Ng, S.W.; Tan, R.W. Ardeemins and cytochalasins from Aspergillus terreus residing in Artemisia annua. Planta Med. 2010, 76, 1616–1621. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Wang, J.S.; Shen, H.J.; Song, Y.C.; Tan, R.X. A new cytotoxic trichothecene macrolide from the endophyte Myrothecium roridum. Planta Med. 2010, 76, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.M.; Song, Y.C.; Chen, J.R.; Hu, S.; Wu, J.Y.; Tan, R.X. Paranolin: A new xanthene-based metabolite from Paraphaeosphaeria nolinae. Helv. Chim. Acta 2006, 89, 502–506. [Google Scholar] [CrossRef]
- Ngankaranatikarn, P.; Chuanasa, T.; Sriubolmas, N.; Suwanborirux, K. Antileukemic activity and secondary metabolites of an endophytics fungus Phomopsis sp. from Artemisia annua. Thai J. Pharm. Sci. 2013, 38, 195–198. [Google Scholar]
- Lösgen, S.; Magull, J.; Schulz, B.; Draeger, S.; Zeeck, A. Isofusidienols: Novel chromone-3-oxepines produced by the endophytic fungus Chalara sp. Eur. J. Org. Chem. 2008, 698–703. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Order RA % | Family RA % | Genus | Genus RA % Per Host Plant/Plants | Host Plants | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amphisphaeriales 1.92 | Pestalotiopsidaceae 2.13 | Pestalotiopsis | 0.9 | 1.19 | 11.76 | A. capillaris* + A. indica* + A. lactiflora* (HK); A. thuscula (TF); A. argy* (CH) | ||||||||
| Botryosphaeriales 7.69 | Aplosporellaceae 2.13 | Aplosporella | 1.22 | A. thuscula (TF) | ||||||||||
| Botryosphaeriaceae 6.38 | Botryosphaeria | 41.67 | A. lavandulifolia (CH) | |||||||||||
| Macrophomina | 2.44 | A. thuscula (TF) | ||||||||||||
| Neofusicoccum | 2.78 | 30.3 | 10.98 | A. lavandulifolia; A. thuscula (LP); A. thuscula (TF) | ||||||||||
| Cantharellales 1.92 | Ceratobasidiaceae 2.13 | Rhizoctonia | 16.67 | A. brachyloba (CH) | ||||||||||
| Capnodiales 3.85 | Cladosporiaceae 2.13 | Cladosporium | 2.44 | 2.77 | 5.88 | 8.69 | A. thuscula (TF); A. annua* (CH); A. argy*; A. nilagirica* (IN) | |||||||
| Dissoconiaceae 2.13 | Ramichloridium | 2.77 | A. annua* | |||||||||||
| Diaporthales 3.85 | Diaporthaceae 4.26 | Diaporthe | 5.56 | 16.67 | 8.33 | 9.09 | 5.88 | A. lavandulifolia; A. brachyloba; A. scoparia (CH); A. thuscula (LP); A. argy* | ||||||
| Phomopsis | 6.3 | 5.88 | 36.36 | A. capillaris +A. indica + A. lactiflora*; A. argy*; A. lactiflora* (CH) | ||||||||||
| Dothideales 1.92 | Dothioraceae 2.13 | Aureobasidium | 0.9 | 3.66 | A. capillaris + A. indica + A. lactiflora*; A. thuscula (TF) | |||||||||
| Eurotiales 7.69 | Trichocomaceae 8.51 | Aspergillus | 8.33 | 4.55 | 8.69 | A. lavandulifolia; A. thuscula (LP); A. nilagirica* | ||||||||
| Eupenicillium | 5.55 | A. annua* | ||||||||||||
| Paecilomyces | 2.77 | A. annua* | ||||||||||||
| Penicillium | 2.78 | 1.52 | 13.8 | 5.88 | 33.33 | 13.4 | A. lavandulifolia; A. thuscula (LP); A. annua*; A. argy*; A. nilagirica*; A. annua | |||||||
| Glomerellales 1.92 | Glomerellaceae 2.13 | Colletotrichum | 19.81 | 11.1 | 25 | 5.55 | 5.88 | 10.11 | 9.09 | A. capillaris + A. indica + A. lactiflora*; A. lavandulifolia; A. brachyloba; A. annua*; A. argy*; A. annua; A. lactiflora* (CH) | ||||
| Hypocreales 17.31 | Clavicipitaceae 2.13 | Ephelis | 1.8 | A. capillaris + A. indica + A. lactiflora* | ||||||||||
| Hypocreaceae 4.26 | Hypocrea | 5.88 | 9.09 | A. argy*; A. lactiflora* | ||||||||||
| Trichoderma | 4.34 | A. nilagirica* | ||||||||||||
| Nectriaceae 4.26 | Fusarium | 5.55 | 5.88 | 4.34 | A. annua*; A. argy*; A. nilagirica* | |||||||||
| Nectria | 2.44 | A. thuscula (TF) | ||||||||||||
| Stachybotryaceae 4.26 | Stachybotrys | 1.22 | A. thuscula (TF) | |||||||||||
| Myrothecium | N.A. | A. nilagirica* | ||||||||||||
| Acremonium | 13 | A. annua | ||||||||||||
| Cephalosporium | N.A. | A. annua | ||||||||||||
| Mucorales 3.85 | Mucoraceae4.26 | Rhizopus | 4.34 | A. nilagirica* | ||||||||||
| Mucor | N.A. | A. annua | ||||||||||||
| Onygenales 1.92 | Arthrodermataceae 2.13 | Arthroderma | 4.34 | A. nilagirica* | ||||||||||
| Pleosporales 25.00 | Sporormiaceae 2.13 | Preussia | 10.61 | 10.98 | A. thuscula (LP); A. thuscula (TF) | |||||||||
| Didymosphaeriaceae 2.13 | Tremateia | 1.52 | A. thuscula (LP) | |||||||||||
| Leptosphaeriaceae 4.26 | Coniothyrium | 1.52 | A. thuscula (LP) | |||||||||||
| Leptosphaeria | N.A. | A. annua | ||||||||||||
| Pleosporaceae 14.89 | Alternaria | 24.30 | 13.89 | 75 | 25 | 66.67 | 33.33 | 33.33 | 23.5 | 36.36 | 28.79 | 41.46 | A. capillaris + A. indica + A. lactiflora*; A. lavandulifolia; A. tangutica; A. brachyloba; A. subulata; A. argy; A. scoparia; A. argy*; A. lactiflora*; A. thuscula (LP); A. thuscula (TF) | |
| Curvularia | 16.66 | 33.33 | 16.67 | 8.33 | 1.52 | A. tangutica (CH); A. brachyloba (CH); A. subulata (CH); A. argy; A. thuscula (LP) | ||||||||
| Drechslera | 0.9 | A. capillaris + A. indica + A. lactiflora* | ||||||||||||
| Edenia | 2.77 | A. annua* | ||||||||||||
| Neoplatysporoides | 3.03 | A. thuscula (LP) | ||||||||||||
| Stemphylium | 0.65 | 4.88 | A. thuscula (LP); A. thuscula (TF) | |||||||||||
| Paraphoma | 1.52 | A. thuscula (LP) | ||||||||||||
| Phoma | 1.52 | 7.32 | 2.77 | 4.34 | A. thuscula (LP); A. thuscula (TF); A. annua*; A. nilagirica* | |||||||||
| Pleosporineae 1.92 | Camarosporiaceae 2.13 | Camarosporium | 1.52 | 1.22 | A. thuscula (LP); A. thuscula (TF) | |||||||||
| Pythiales 1.92 | Pythiaceae 2.13 | Pythium | 4.34 | A. nilagirica* | ||||||||||
| Saccharomycetales 1.92 | Debaryomycetaceae 2.13 | Meyerozyma | 9.09 | A. lactiflora* | ||||||||||
| Sordariales 3.85 | Chaetomiaceae 4.26 | Chaetomium | 2.44 | 11.76 | A. thuscula (TF); A. argy* | |||||||||
| Thielavia | 2.44 | A. thuscula (TF) | ||||||||||||
| Sporidiobolales 1.92 | Sporidiobolaceae 2.13 | Rhodotorula | 5.88 | A. argy* | ||||||||||
| Xylariales 9.62 | Apiosporaceae 4.26 | Arthrinium | 5.88 | A. argy* | ||||||||||
| Nigrospora | 5.56 | 33.33 | 75 | 25 | 3.03 | A. lavandulifolia; A. brachyloba; A. argy; A. scoparia; A. thuscula (LP) | ||||||||
| Graphostromataceae 2.13 | Biscogniauxia | 2.44 | A. thuscula (TF) | |||||||||||
| Hypoxylaceae 2.13 | Hypoxylon | N.A. | A. annua | |||||||||||
| Artemisia Species | Plant Part | EF Taxa | EF Identification Method | Compound/Extract | Class of Compounds | Bioactivity | Reference |
|---|---|---|---|---|---|---|---|
| A. absinthium | root | Penicillium spp. (strains B6C18 and B14C36) | N.A. | spore solution (106 mL−1) | N.A. | Antifungal inhibition of diameter of lesions on vine berries: 75% and 91% | [73] |
| A. annua | stem | Aspergillus spp. | morphology | ethyl acetate crude extract | N.A. | Antibacterial, antifungal | [41] |
| Mucor spp. | |||||||
| Cephalosporium spp. | |||||||
| Fusarium spp. | Antibacterial | ||||||
| Aspergillus terreus (strain IFB-E030) | N.A. | 10-phenyl-[12]-cytochalasins Z17 | Cytochalasan, alkaloids | Cytotoxic IC50 = 26.2 µM | [75] | ||
| Colletotrichum sp. | morphology | 3β-hydroxy-ergosta-5-ene | Ergosterol derivatives | Antifungal %I (200 µg mL−1): 77–85% | [40] | ||
| Antimicrobial MIC (µg mL−1): 50–75 | |||||||
| 3-oxo-ergosta-4,6,8(11),22-tetraene | Triunsaturated steroids | Antifungal %I (200 µg mL−1): 75% | |||||
| Antimicrobial MIC (µg mL−1): 25–75% | |||||||
| 3β-hydroxy-5α,8α-epidioxy-ergosta-6,22-diene | Ergosterol peroxides | Antimicrobial MIC (µg mL−1): 50–75 | |||||
| 6-isoprenylindole-3-carboxylic acid | indoles | Antifungal %I (200 µg mL−1): 57–77% | |||||
| Antimicrobial MIC (µg mL−1): 25–75 | |||||||
| 3β,5α-dihydroxy-6β-acetoxy-ergosta-7,22-diene | sterols | Antifungal %I (200 µg mL−1): 75–77% | |||||
| Antimicrobial MIC (µg mL−1): 50–100% | |||||||
| 3β,5α-dihydroxy-6β-phenylacety- loxy-ergosta-7,22-diene | Antifungal %I (200 µg mL−1): 50–66% | ||||||
| Antimicrobial MIC (µg mL−1): 50–75 | |||||||
| Hypoxylon trunctatum | morphology and 18S rDNA sequence | daldinone C | Benzo[j]fluoranthenes (polycyclic aromatic hydrocarbons) | Cytotoxic IC50 = 49.5 µM | [42] | ||
| daldinone D | Cytotoxic IC50 = 41 µM | ||||||
| Myrothecium roridum (strain IFB-E012) | myrothecine A | Sesquiterpene-based trichothecenes | Cytotoxic IC50 = 8.5 µg mL−1 | [69,76] | |||
| myrothecine B | Cytotoxic IC50 = 0.76 µg mL−1 | ||||||
| myrothecine C | Cytotoxic IC50 = 32.21 µg mL−1 | ||||||
| roridin E | Macrocyclic trychothecenes | Cytotoxic IC50 = 0.03 µg mL−1 | |||||
| mytoxin B | Trychothecene macrolides | Cytotoxic IC50 = 0.002 µg mL−1 | |||||
| dihydromyrothecine C | Cytotoxic IC50 = 44.48 µM | ||||||
| roritoxin E | Cytotoxic IC50 = 0.26 µg mL−1, 10.54 µg mL−1 | ||||||
| Aspergillus fumigatus | 5-N-acetylardeemin | Pyrimidinones (aromatic heterocyclic diazenes) | Cytotoxic RF = 1.5–6 µM | [65] | |||
| 5-N-acetyl-15bβ-hydroxyardeemin | Cytotoxic RF = 2–9.5 µM | ||||||
| 5-N-acetyl-15b-didehydroardeemin | Cytotoxic RF = 2.5–8 µM | ||||||
| 5-N-acetyl-16α-hydroxyardeemin | Cytotoxic RF = 1.5–7 µM | ||||||
| Colletotrichum gloeosporioides | morphology | oligosaccharides extract elicitor for artemisinin | oligosaccharides | increment of artemisinin = 64.29% | [62] | ||
| Penicillium oxalicum | ITS1, 5.8S, and ITS2 | heat-killed mycelium degrading triclosan (TCS) | N.A. | max. adsorbing capacity of TCS = 17.60 mg g−1 | [74] | ||
| leaves | Curvularia pallescens | elicitor extract for artemisinin | N.A. | 2–6% elicitor extract (w/v): artemisinin content = 1.21–3.47-times more than control | [59] | ||
| Cladosporium sp. | morphology | ethyl acetate extract | N.A. | Antibacterial | [66] | ||
| stem | Paraphaeosphaeria nolinae (strain IFB-E011) | N.A. | paranolin | polycyclic aromatic compounds (xanthene-based) | Cytotoxic IC50 values > 50 µg mL−1 | [77] | |
| Phomopsis sp. | crude EtOAc extract = BB1, fraction Hex/EtOAc/MeOH = BB4, fractions MeOH = BB8, BB9, BB10 | N.A. | Cytotoxic: IC50 (µg mL−1) BB1 = 17.11, BB4 = 0.7; %I (20 µg/mL) BB8 = 1.97, BB9 = 0.53, BB10 = 52.98 | [78] | |||
| tyrosol (purified from BB10) | Phenolic compounds | ||||||
| A. argyi | N.A. | Phomopsis sp. (strain H31) | ITS1, 5.8S, and ITS2 | ethyl acetate + ethyl acetate/water extract | N.A. | Antifungal %I (1 mg mL−1) = 34.57; Cytotoxic %I (20 µg mL−1) = 81.69 | [29] |
| Cladosporium sp. (strain H23) | Antifungal %I (1 mg mL−1) = 23.46 | ||||||
| Chaetomium sp. (strain H40) | Antibacterial | ||||||
| Penicillium sp. (strain H9) | Antibacterial | ||||||
| Pestalotiopsis sp. (strain H14) | Antibacterial; Cytotoxic %I (20 µg mL−1) = 53.47, 71.43, 97.03 | ||||||
| Diaporthe sp. (strain H26) | Antibacterial | ||||||
| Trichoderma sp. (strain H17) | Antibacterial | ||||||
| Rhodotorula sp. (strain H13) | Cytotoxic %I (20 µg mL−1) = 16.41, 23.56, 78.28 | ||||||
| Colletotrichum sp. (strain H42) | Cytotoxic %I (20 µg mL−1) = 11.22, 26.95, 71.99 | ||||||
| stem | Nigrospora sphaerica | morphology and ITS1, 5.8S, and ITS2 | ethyl acetate crude extract | N.A. | Antifungal %I (1 mg mL−1): 48.37, 11.94, 24.92 | [23] | |
| Trichoderma koningiopsis (strain QA-3) | ITS1, 5.8S, and ITS2 | ent-koninginin A | Tricyclic polyketides | Antifungal MIC (µg mL−1): 8–64 Antibacterial MIC (µg mL−1): 8–64 | [68] | ||
| 1,6-di-epi-koninginin A | Antifungal MIC (µg mL−1): 64 Antibacterial MIC (µg mL−1): 32–64 | ||||||
| 15-hydroxykoninginin A | Antifungal MIC (µg mL−1): 64 Antibacterial MIC (µg mL−1): 16–64 | ||||||
| 10-deacetylkoningiopisin | Bicyclic polyketides | Antifungal MIC (µg mL−1): 8–64 Antibacterial MIC (µg mL−1): 32–64 | |||||
| koninginin T | Tricyclic polyketides | Antifungal MIC (µg mL−1): 16 Antibacterial MIC (µg mL−1): 32–64 | |||||
| koninginin L | Antifungal MIC (µg mL−1): 16 Antibacterial MIC (µg mL−1): 32–64 | ||||||
| trichoketide A | Bicyclic polyketides | Antifungal MIC (µg mL−1): 4–64 Antibacterial MIC (µg mL−1): 16–64 | |||||
| A. brachyloba | Alternaria alternata | morphology and ITS1, 5.8S, and ITS2 | ethyl acetate crude extract | N.A. | Antifungal %I (1 mg mL−1): 29.8, 33.2, 42.4 | [23] | |
| A. capillaris | flower | Alternaria sp. (strain Acap F1) | ITS1, 5.8S, and ITS2 | ethyl acetate extract of mycelia; Trolox equivalent antioxidant content (TEAC), Total phenolic content (TPC) | phenolic acids, flavonoids | TEAC (µM trolox equivalent/100 mL culture) = 127.41; TPC (mg gallic acid equivalent/100 mL culture) = 13.71 | [22,72] |
| Phomopsis sp. (strain Acap. F3) | TEAC (µM trolox equivalent/100 mL culture) = 526.93; TPC (mg gallic acid equivalent/100 mL culture) = 49.22 | ||||||
| Phomopsis sp. (strain AcapF4) | TEAC (µM trolox equivalent/100 mL culture) = 177.12; TPC (mg gallic acid equivalent/100 mL culture) = 18.43 | ||||||
| A. indica | leaves | Colletotrichum sp. (strain AiL1) | TEAC (µM trolox equivalent/100 mL culture) = 100.2; TPC (mg gallic acid equivalent/100 mL culture) = 6.88 | ||||
| Xylaria sp. (strain AiL3) | TEAC (µM trolox equivalent/100 mL culture) = 102.21; TPC (mg gallic acid equivalent/100 mL culture) = 8.69 | ||||||
| Diaporthe sp. (strain AiL4) | TEAC (µM trolox equivalent/100 mL culture) = 217.76; TPC (mg gallic acid equivalent/100 mL culture) = 21.27 | ||||||
| A. lactiflora | N.A. | Phomopsis sp. (strain GYBH42) | ethyl acetate crude extract | N.A. | Cytotoxic, antioxidant | [39] | |
| Alternaria sp. (strain GYBH47) | |||||||
| A. lavandulifolia | stem | Cochliobolus geniculatus | morphology and ITS1, 5.8S, and ITS2 | Antifungal EC50 = 0.03; %I (1 mg mL−1) = 31.5 and 100 | [23] | ||
| Botryosphaeria dothidea | Antifungal %I (1 mg mL−1): 85.79, 22.4 | ||||||
| A. mongolica | Colletotrichum gloeosporioides | morphology | colletotric acid | Phenolic acids | Antibacterial MIC (µg mL−1): 25–50 | [64] | |
| A. subulata | Curvularia spicifera | morphology and ITS1, 5.8S, and ITS2 | ethyl acetate crude extract | N.A. | Antifungal %I (1 mg mL−1): 8.95, 52.09, 77.9 | [23] | |
| A. vulgaris | N.A. | Chalara sp. (strain 6661) | N.A. | isofusidienol A, B, C, and D | chromones (benzopyran derivatives) | Antibacterial | [79] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cosoveanu, A.; Cabrera, R. Endophytic Fungi in Species of Artemisia. J. Fungi 2018, 4, 53. https://doi.org/10.3390/jof4020053
Cosoveanu A, Cabrera R. Endophytic Fungi in Species of Artemisia. Journal of Fungi. 2018; 4(2):53. https://doi.org/10.3390/jof4020053
Chicago/Turabian StyleCosoveanu, Andreea, and Raimundo Cabrera. 2018. "Endophytic Fungi in Species of Artemisia" Journal of Fungi 4, no. 2: 53. https://doi.org/10.3390/jof4020053
APA StyleCosoveanu, A., & Cabrera, R. (2018). Endophytic Fungi in Species of Artemisia. Journal of Fungi, 4(2), 53. https://doi.org/10.3390/jof4020053