
Disentangling a Complex of Violet, Endangered Species of Clavaria Subsumed Under the Misapplied Name Clavaria zollingeri Lév. (Clavariaceae, Fungi)

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Herbarium Specimens and Morphological Study
2.2. Nomenclature
2.3. DNA Extraction, PCR Amplification, Sequencing and Alignment
2.4. Phylogenetic Analyses and OTU Delimitation
3. Results
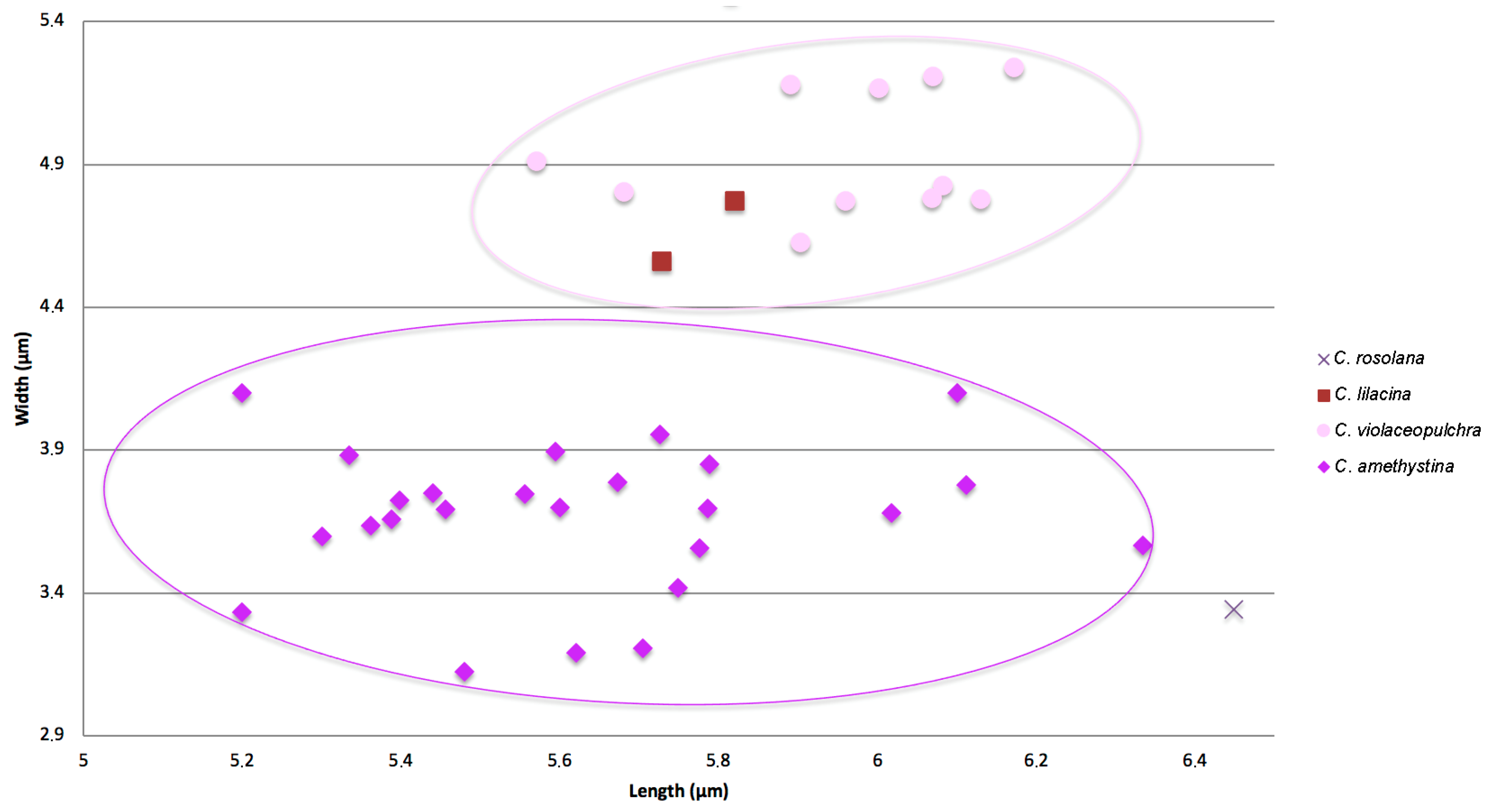
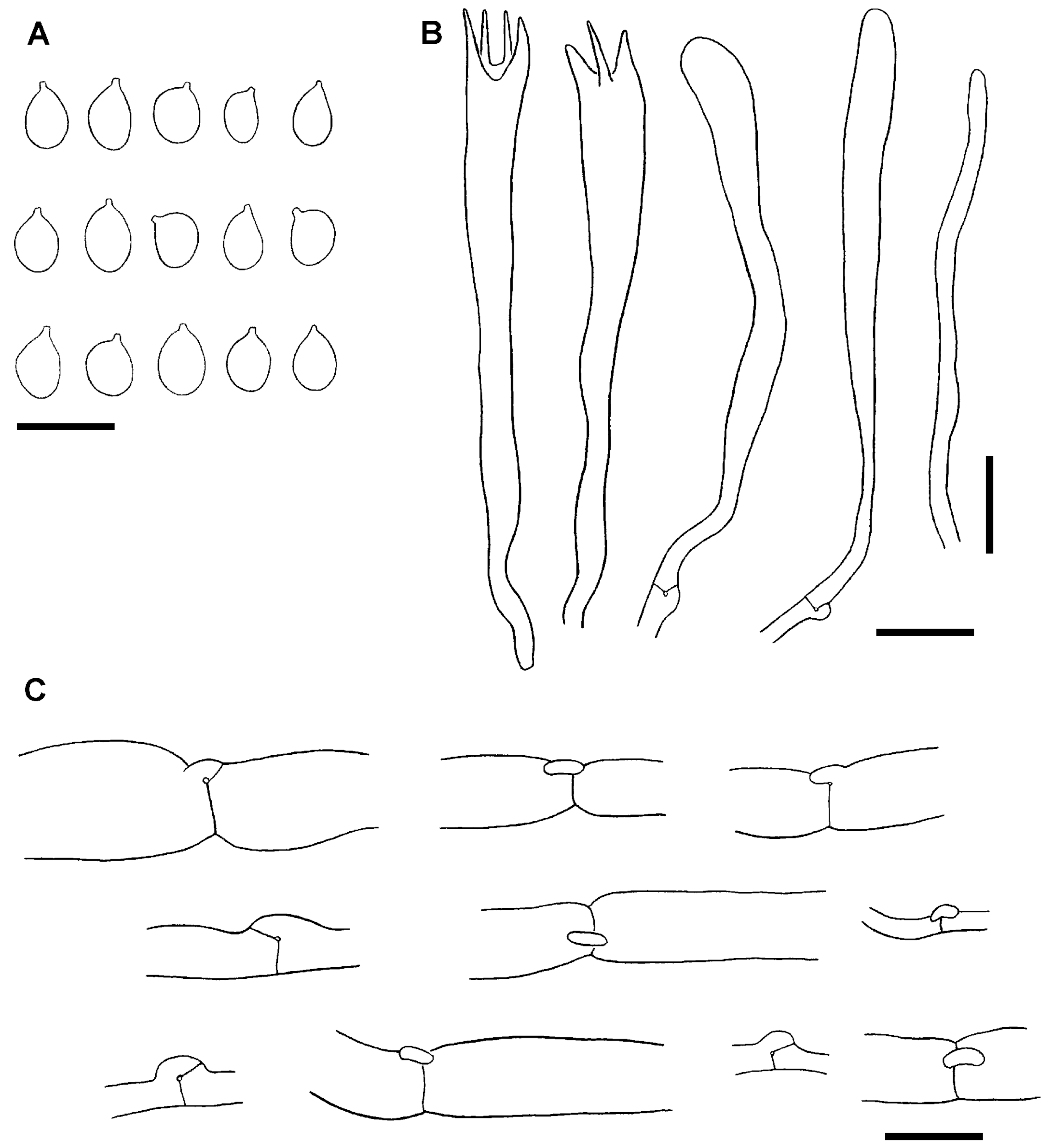
3.1. Morphological Study
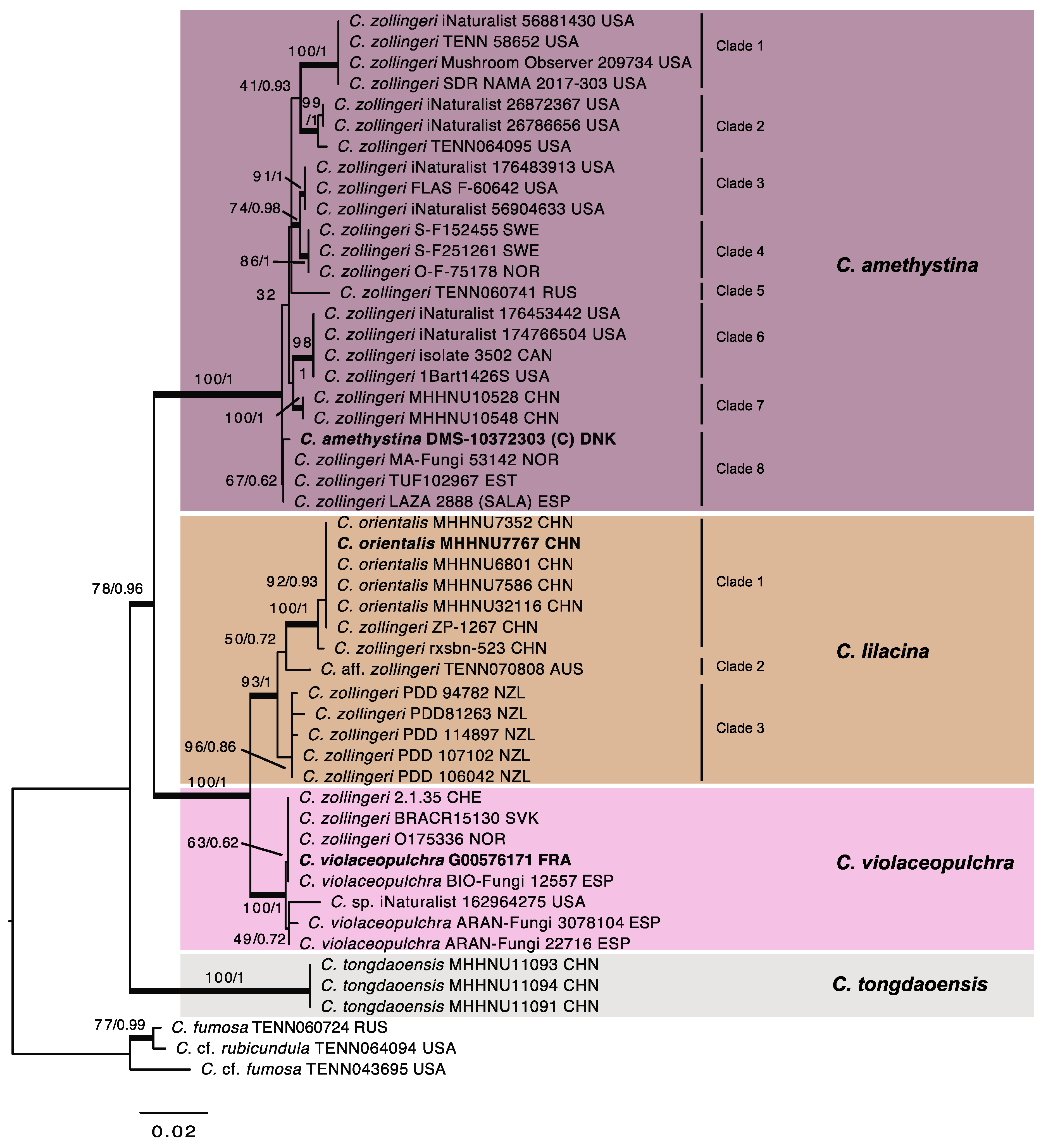
3.2. Molecular Study
3.3. Type Study of Clavaria zollingeri
3.4. Taxonomic Treatment
3.4.1. Clavaria amethystina
- Description
- Distribution and ecology
- Specimens examined
- Taxonomic comments
- Nomenclatural comments on Clavaria amethystina and its synonyms
3.4.2. Clavaria lilacina
- Description
- Distribution and ecology
- Specimens examined
- Taxonomic comments
- Nomenclatural comments
3.4.3. Clavaria violaceopulchra
- Description
- Distribution and ecology
- Distribution and ecology
- Taxonomic comments
3.4.4. New Combination
3.4.5. Excluded Material
3.4.6. Excluded Names
- Clavaria alcicornis Zoll. & Moritzi in Zollinger, Natuur-Geneesk. Arch. Ned.-Indië 1: 382. 1844. Lectotype: Indonesia, Java, Tjikoya, March 1843, Plantae Javanicae 1125 (FH) (designation achieved by Petersen [73] accepting it as the holotype, according to Arts. 7.11 and 9.10).
- Clavaria amethystina subsp. coerulescens P. Karst., Meddeland. Soc. Fauna Fl. Fenn. 16: 2. 1888. Type specimen: not kept at H (checked in person by I. Olariaga), probably lost.
- Clavaria bizzozeriana Sacc., Syll. Fung. 6: 693. 1888. [“Bizzozeriana”][nom. nov. based on Clavaria tenuissima Sacc., Michelia 1(4): 436. 1878, nom. illeg. Art. 53, non Clavaria tenuissima Lév., Ann. Sci. Nat., Bot., sér. 3, 5: 156. 1846]. ≡ Ramariopsis bizzozeriana (Sacc.) Schild, Z. Pilzk. 38: 26. 1972. ≡ Clavulinopsis bizzozeriana (Sacc.) Jülich, Int. J. Mycol. Lichenol. 2(1): 120. 1985. Syntypes: Italy, Padova, in uliginosis, October 1878, leg. G. Bizzozero, Mycotheca Veneta 1309 (B, BM, BUCM, DBN, E, FH, GE, HBG, K, L, M, PRE, S, SIENA, STR, TLA, TO, W, WRSL).
- Clavaria brachycera Pers., Comm. Fung. Clav.: 61. 1797. Type specimen: not kept at L, probably lost.
- Clavaria nymaniana Henn. in Warburg, Monsunia 1: 9. 1900. Type: lost (see below).
- Clavaria violacea Vill., Hist. Pl. Dauphiné 3(2): 1050. 1789. non Clavaria violacea Petch, Ann. Roy. Bot. Gard. Peradeniya 7(4): 290. 1922 [nom. illeg. Art. 53.1].
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Léveillé, J.H. Descriptions des Champignons de l’herbier du Muséum de Paris. Ann. Sci. Nat. Sér. 3 1846, 5, 111–167. [Google Scholar]
- Coker, W.C. The Club and Coral Mushrooms (Clavarias) of the United States and Canada; University of North Carolina Press: Chapel Hill, NC, USA, 1923. [Google Scholar]
- Vasco-Palacios, A.M.; Franco-Molano, A.E. Diversity of Colombian Macrofungi (Ascomycota-Basidiomycota). Mycotaxon 2013, 121, 1–58. [Google Scholar]
- Yan, J.; Xiong, L.; Yang, L.-X.; He, Z.-M.; Zhang, P.; Liao, K. Morphological and Multi-Locus Phylogenetic Analyses Reveal Three New Branched Species of Clavaria (Clavariaceae, Agaricales) from China. MycoKeys 2025, 115, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.H. The Clavarioid Fungi of New Zealand; Information Series; Department of Scientific and Industrial Research: Auckland, New Zealand, 1988; Volume 236, pp. 1–170.
- Holec, J.; Beran, M. Červený Seznam Hub (makromycetů) České Republiky [Red List of Fungi (macromycetes) of the Czech Republic]. Příroda, Praha 2006, 24, 1–282. [Google Scholar]
- Moeslund, J.E.; Nygaard, B.; Ejrnæs, R.; Alstrup, V.; Baagøe, H.J.; Bell, N.; Bruun, L.D.; Bygebjerg, R.; Carl, H.; Christensen, M.; et al. Den Danske Rødliste; Aarhus Universitet, DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2023. [Google Scholar]
- Brandrud, T.E.; Bendiksen, E.; Blaalid, R.; Hofton, T.H.; Jordal, J.B.; Nordén, J.; Nordén, B.; Wollan, A.K. Sopper: Vurdering av Fiolett Greinkøllesopp Clavaria Zollingeri for Norge. Rødlista for Arter 2021; Artsdatabanken: Trondheim, Norway, 2021. [Google Scholar]
- Wojewoda, W.; Ławrynowicz, M. Red List of the Macrofungi in Poland. In Red List of Plants and Fungi in Poland; Mirek, Z., Zakrzycki, K., Wojewoda, W., Szeląg, Z., Eds.; Szafer Institute of Botany, Polish Academy of Science: W. Kraków, Poland, 2006; pp. 53–70. [Google Scholar]
- Knutsson, T.; Krikorev, M.; Ottosson, E.; Dahlberg, A.; Edman, M.; Hansen, K.; Jeppson, M.; Karström, M.; Larsson, E.; Nitare, J.; et al. 2020. Rödlista 2020—Expertkommittén För Svampar; SLU Artdatabanken: Uppsala, Sweden, 2020. [Google Scholar]
- Jordal, J.; Kautmanová, I. Clavaria zollingeri. The IUCN Red List of Threatened Species 2019: E.T70402563A70402575. Available online: https://doi.org/10.2305/IUCN.UK.2019-2.RLTS.T70402563A70402575.en (accessed on 5 April 2024).
- Corner, E.J.H. A Monograph of Clavaria and Allied Genera. Ann. Bot. Mem. 1950, 1, 1–740. [Google Scholar]
- Knudsen, H.; Vesterholt, J. Agaricoid, Boletoid, Clavarioid, Cyphelloid and Gastroid Genera. In Funga Nordica, 2nd ed.; Nordsvamp: Copenhagen, Denmark, 2012. [Google Scholar]
- Donk, M.A. Revision Der Niederländischen Homobasidiomycetae-Aphyllophoraceae II. Meded. Ned. Mycol. Ver. 1933, 22, 1–278. [Google Scholar]
- Petersen, R.H. Notes on Clavarioid Fungi. XV. Reorganization of Clavaria, Clavulinopsis and Ramariopsis. Mycologia 1978, 70, 660–671. [Google Scholar] [CrossRef]
- Dentinger, B.T.M.; McLaughlin, D.J. Reconstructing the Clavariaceae Using Nuclear Large Subunit rDNA Sequences and a New Genus Segregated from Clavaria. Mycologia 2006, 98, 746–762. [Google Scholar] [CrossRef]
- Kautmanová, I.; Tomšovský, M.; Dueñas, M.; Martín, M.P. European Species of Clavaria (Agaricales, Agaricomycetes) with Dark Basidiomata—A Morphological and Molecular Study. Persoonia 2012, 29, 133–145. [Google Scholar] [CrossRef]
- Schaeffer, J.C. Fungorum qui in Bavaria et Palatinatu Circa Ratisbonam Nascuntur Icons Natives Coloribus Expressae; typis Zunkelianis: Regensburg, Germany, 1774; Volume 4. [Google Scholar]
- Holmskjold, T. Beata Ruris Otia Fungis Danicis; Friderico Brummer: Havniae [Kjøbenhavn], Denmark, 1790; Volume 1. [Google Scholar]
- Nees von Esenbeck, C.G. Das System der Pilze und Schwamme. Ein Versuch; Stahelschen Buchhandlung: Würzburg, Germany, 1816. [Google Scholar]
- Venturi, A. Studi Micologici; Francisco Cavalieri: Brescia, Italy, 1842. [Google Scholar]
- Persoon, C.H. Commentatio de Fungis Clavaeformibus; Petrum Philippum Wolf: Lipsiae, Germany, 1797. [Google Scholar]
- Villars, M. Histoire des Plantes de Dauphiné. Tome Troisieme. Contenant la Deuxieme Partie & Les Planches des Deux Volumes; Grande Chartreuse: Briançon, France; Gap & de Montelimar: Briançon, France; chez les frères Perisse: Lyon, France; chez Prevost: Paris, France; Grenoble, France, 1789. [Google Scholar]
- Zollinger, H. Observationes Phytographicae Praecipur Genera et Species Nova Nonnula Respicientes. Natuur Geneesk. Arch. Ned. Indië 1844, 1, 372–405. [Google Scholar]
- Olariaga, I. The Order Cantharellales in the Iberian Peninsula and the Balearic Islands; University of the Basque Country (UPV/EHU): Bilbao, Spain, 2009. [Google Scholar]
- Franchi, P.; Marchetti, M. I Funghi Clavarioidi in Italia. Bresadola, A.M., Ed.; Graffica Sette: Trento, Italy, 2021; Volume 1. [Google Scholar]
- Kautmanová, I. Taxonomy of Central-European Representatives of the Genus Clavaria (Basidiomycetes, Clavariaceae); University of Bratislava: Bratislava, Slovakia, 2012. [Google Scholar]
- The Royal Horticultural Society. R.H.S. Colour Charts; RHS: London, UK, 1995. [Google Scholar]
- Munsell Color Corporation Soil Color Charts; Macbeth Division of Kollmorgen Instruments Corporation: New York, NY, USA, 1990.
- Thiers, B. Updated Continuously. Index Herbariorum. A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2019; Available online: http://sweetgum.nybg.org/ih/ (accessed on 9 April 2025).
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Gandhi, K.N.; Gravendyck, J.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Klopper, R.R.; Knapp, S.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Madrid Code) Adopted by the Twentieth International Botanical Congress Madrid, Spain, July 2024; Regnum Vegetabile; The University of Chicago Press: Chicago, IL, USA, 2025. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 9780123721808. [Google Scholar]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalis, R. Characterization of Anastomosis Groups of Binucleate Rhizoctonia species Using Restriction Analysis of an Amplified Ribosomal RNA Gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A Multiple Sequence Alignment Method with Reduced Time and Space Complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Birkebak, J.M.; Adamčík, S.; Looney, B.P.; Matheny, P.B. Multilocus Phylogenetic Reconstruction of the Clavariaceae (Agaricales) Reveals Polyphyly of Agaricoid Members. Mycologia 2016, 108, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- ISO 3166-1; Codes for the Representation of Names of Countries and Their Subdivisions – Part 1: Country Code. International Organization for Standardization: Geneve, Switzerland, 2013.
- Van Overeem, C. Beiträge Zur Pilzflora von Niederländisch Indien. Bull. Jard. Bot. Buitenzorg 1923, 5, 247–293. [Google Scholar]
- Zollinger, H. Systematisches Verzeichniss der im Indischen Archipel in Den Jahren 1842–1848 Gesammelten Sowie der aus Japan Empfangenen Pflanzen; Verlag von E. Kiesling: Zürich, Switzerland, 1854. [Google Scholar]
- Moritzi, A. Systematisches Verzeichniss der von H. Zollinger; Verlag des Verfassers: Solothurn, Switzerland, 1846. [Google Scholar]
- Petersen, R.H. Notes on Clavarioid Fungi. Persoonia 1971, 6, 219–229. [Google Scholar]
- Cotton, A.D.; Wakefield, E.M. A Revision of the British Clavariae. Trans. Br. Mycol. Soc. 1919, 6, 164–198. [Google Scholar] [CrossRef]
- Fries, E.M. Systema Mycologicum; Ernesti Mauritii: Greifswaldiae, Germany, 1821; Volume 1. [Google Scholar]
- Schaeffer, J.C. Fungorum qui in Bavaria et Palatinatu Circa Ratisbonam Nascuntur Icons Natives Coloribus Expressae; typis Zunkelianis: Regensburg, Germany, 1763; Volume 2. [Google Scholar]
- Bulliard, G.B.F. Herbier de la France; Didot J.ne, Debure, Libraires: Paris, France, 1791; Volume 11. [Google Scholar]
- Berkeley, M.J. Outlines of British Fungology; Lovell Reeve: London, UK, 1860. [Google Scholar]
- Cooke, M.C. A Plain and Easy Account of British Fungi; Robert Hardwicke: London, UK, 1862. [Google Scholar]
- Fries, E.M. Hymenomycetes Europaei; Ed. Berling: Uppsala, Sweden, 1874. [Google Scholar]
- Pilát, A. Přehled Hub Kyjankovitych-Clavariaceae Se Zvláštním Zřetelem K československým Druhûm. Sborn. Nár. Mus. Praze Řada B Přir. Vědy 1958, 14, 129–255. [Google Scholar]
- Jülich, W. Die Nichtblätterpilze, Gallerpilze und Bauchpilze Kleine Kryptogamenflora Band II; Gustav Fischer Verlag: Sttutgart, Germany, 1984. [Google Scholar]
- Olariaga, I.; Jugo, B.M.; García-Etxebarria, K.; Salcedo, I. Species Delimitation in the European Species of Clavulina (Cantharellales, Basidiomycota) Inferred from Phylogenetic Analyses of ITS Region and Morphological Data. Mycol. Res. 2009, 113, 1261–1270. [Google Scholar] [CrossRef]
- Calonge, F.D.; de Sequeira, M.M.; Rocha, E.; Hernández, C.J.C. Algunos Hongos Interesantes de Madeira (Portugal). Bull. Sem. Féd. Assoc. Mycol. Médit. 2010, 37, 29–34. [Google Scholar]
- Castro-Marcote, J.L. Clavulina amethystina (Bull.) Donk. In Bolets de Catalunya 29; Impressors de Barcelona S.L.: Barcelona, Spain, 2010; p. 1420. [Google Scholar]
- Petersen, R.H.; Olexia, P.D. Type Studies in the Clavarioid Fungi. I. The Taxa Described by Charles Horton Peck. Mycologia 1967, 59, 767–802. [Google Scholar] [CrossRef]
- Petch, B.A. Additions to Ceylon Fungi (II.). Ann. Roy. Bot. Gard. (Peradeniya) 1922, 7, 279–322. [Google Scholar]
- Van Overeem, C. Clavariaceae. In Icones Fungorum Malayensium 4; Van Overeem, C., Weese, J., Eds.; Selbstverlag des Mykologischen Museums in Weesp: Wien, Austria, 1923; pp. 1–2. [Google Scholar]
- Petersen, R.H. Type Studies in the Clavariaceae. VI. Four Pivotal Types from the Pacific Tropics. Mycotaxon 1980, 12, 281–286. [Google Scholar]
- Rea, C. British Basidiomycetidae. A Handbook to the Larger British Fungi; University Press: Cambridge, UK, 1922. [Google Scholar]
- Donk, M.A. Nomenclatural Notes on Generic Names of Agarics (Fungi: Agaricales). Bull. Jard. Bot. Buitenzorg Sér. 3 1949, 18, 271–402. [Google Scholar]
- Donk, M.A. The Generic Names Proposed for the Hymenomycetes— III. “Clavariaceae.” Reinwardtia 1954, 2, 441–493. [Google Scholar]
- Donk, M.A. The Generic Names Proposed for Polyporaceae. Persoonia 1960, 2, 173–302. [Google Scholar]
- Donk, M.A. The Generic Names Proposed for Agaricaceae. Nova Hedwigia (Beih.) 1962, 5, 1–320. [Google Scholar]
- Battarra, A.J. Fungi Agri Ariminensis Historia; Typis Martinianis: Faenza, Italy, 1759. [Google Scholar]
- Bulliard, G.B.F. Histoire des Champignons de La France; Barrois le Jeune, Belin: Paris, France, 1791; Volume 1. [Google Scholar]
- Fries, E.M. Systema Mycologicum, Vol. 3. (Index); Ernesti Mauritii: Greifswaldae, Germany, 1832. [Google Scholar]
- iNaturalist. Naturalist: A World-Wide Database of Biodiversity Observations. Available online: https://www.inaturalist.org/observations?place_id=any&subview=map&taxon_id=126160 (accessed on 10 May 2024).
- Petersen, R.H. Type Studies in the Clavariaceae. Sydowia 1967, 21, 105–122. [Google Scholar]
- Saccardo, P.A. Sylloge Fungorum; Typis Seminarii: Padua, Italy, 1888; Volume 6. [Google Scholar]
- Barrelier, J. Plantae per Galliam, Hispaniam et Italiam Observatae; Stephane Ganeau: Paris, France, 1714. [Google Scholar]
- Hennings, P. Fungi. In Monsunia Beiträge zur Kenntniss der Vegetation süd-und Ostasiatischen Monsungebietes; Warburg, O., Ed.; Verlag von Wilhelm Engelman: Leipzig, Germany, 1900; pp. 9–38. [Google Scholar]
- Hein, B. Liste der Arten und Arten und Infraspecifischen taxa von P. Hennings. Englera 1988, 10, 1–374. [Google Scholar]
- Jullien-[l’Abbe]. Decuria Fungorum Quos Legi in Pago Thejiano. Unpublished manuscript kept in the Museum of Natural History of Grenoble (GRM). 1784.
- Fries, E.M. Observationes Mycologicae; Gehr. Bonnier: Kjøbenhavn, Denmark, 1818; Volume 2. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Updated Identification | Original Identification | ITS | LSU | Voucher | Country | Lm × Wm (µm) | Qm |
|---|---|---|---|---|---|---|---|
| C. amethystina | C. zollingeri | KM248911 | — | isolate 3502 | Canada | — | — |
| C. amethystina | C. zollingeri | KP257142 | KP257211 | TFB11857 (TENN060741) | Russia | — | — |
| C. amethystina | C. zollingeri | ON231700 | ON228398 | MHHNU10548 | China | — | — |
| C. amethystina | C. zollingeri | ON231699 | ON228397 | MHHNU10528 | China | — | — |
| C. amethystina | C. zollingeri | PV603497 | — | BRACR21846 | Czech Republic | 6.1 × 3.8 | 1.62 |
| C. amethystina | C. zollingeri | PV399839 | PV400542 | DMS-10372303 (C) | Denmark | 5.7 × 3.2 | 1.78 |
| C. amethystina | C. zollingeri | UDB022810 | — | TUF102967 | Estonia | — | — |
| C. amethystina | C. zollingeri | PV603492 | — | BRACR4318 | Finland | 5.5 × 3.7 | 1.48 |
| C. amethystina | C. zollingeri | PV603473 | — | BRACR13263 | Norway | 6.7 × 3.6 | 1.86 |
| C. amethystina | C. zollingeri | PV603474 | — | BRACR13372 | Norway | 5.4 × 3.7 | 1.47 |
| C. amethystina | C. zollingeri | PV603475 | — | BRACR25780 | Norway | 5.4 × 3.8 | 1.45 |
| C. amethystina | C. zollingeri | PV603476 | — | BRACR25781 | Norway | 5.3 × 3.9 | 1.37 |
| C. amethystina | C. zollingeri | PV603477 | — | O-F-223396 | Norway | 5.6 × 3.7 | 1.51 |
| C. amethystina | C. zollingeri | PV603478 | — | O-F-223492 | Norway | 5.2 × 4.1 | 1.27 |
| C. amethystina | C. zollingeri | PV603496 | — | O-F-223561 | Norway | 6.0 × 3.7 | 1.63 |
| C. amethystina | C. zollingeri | PV603479 | — | O-F-242537 | Norway | 5.3 × 3.6 | 1.47 |
| C. amethystina | C. zollingeri | PV603480 | — | O-F-244573 | Norway | 5.6 × 3.9 | 1.44 |
| C. amethystina | C. zollingeri | PV603481 | — | O-F-244642 | Norway | 5.4 × 3.6 | 1.47 |
| C. amethystina | C. zollingeri | PV603482 | — | O-F-251624 | Norway | 6.3 × 3.6 | 1.78 |
| C. amethystina | C. zollingeri | PV603483 | — | O-F-254064 | Norway | 5.6 × 3.7 | 1.48 |
| C. amethystina | C. zollingeri | PV603484 | — | O-F-257161 | Norway | 5.8 × 3.9 | 1.50 |
| C. amethystina | C. zollingeri | PV603498 | — | O-F-224204 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603495 | — | O-F-065543 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603494 | — | O-F-2461666 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603493 | — | O-F-312241 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603499 | — | O-F-63441 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603500 | — | O-F-161801 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603501 | — | O-F-287940 | Norway | — | — |
| C. amethystina | C. zollingeri | PV603502 | — | O-F-69072 | Norway | — | — |
| C. amethystina | C. zollingeri | UDB035814 | — | O-F-75178 | Norway | — | — |
| C. amethystina | C. zollingeri | PV404188 | JQ415955 | MA-Fungi 53142 | Norway | 5.6 × 3.2 | 1.77 |
| C. amethystina | C. zollingeri | PV603485 | — | BRACR15965 | Slovakia | 5.8 × 3.7 | 1.57 |
| C. amethystina | C. zollingeri | PV603486 | — | BRACR24274 | Slovakia | 5.7 × 4.0 | 1.45 |
| C. amethystina | C. zollingeri | PV603487 | — | BRACR36900 | Slovakia | 5.4 × 3.7 | 1.45 |
| C. amethystina | C. zollingeri | PV603488 | — | BRACR36929 | Slovakia | 6.1 × 4.1 | 1.49 |
| C. amethystina | C. zollingeri | PV603489 | — | BRACR36938 | Slovakia | — | — |
| C. amethystina | C. zollingeri | PV603490 | — | BRACR27015 | Slovakia | — | — |
| C. amethystina | C. zollingeri | PV603491 | — | BRACR28365 | Slovakia | — | — |
| C. amethystina | C. zollingeri | PV399836 | PV399846 | LAZA 2888 (SALA) | Spain | 5.5 × 3.1 | 1.76 |
| C. amethystina | C. zollingeri | PV399837 | PV399847 | S-F152455 | Sweden | 5.8 × 3.6 | 1.64 |
| C. amethystina | C. zollingeri | PV399838 | PV399848 | S-F251261 | Sweden | 5.2 × 3.3 | 1.57 |
| C. amethystina | C. zollingeri | KP257141 | HQ877700 | JMB08040912 (TENN064095) | USA | — | — |
| C. amethystina | C. zollingeri | ON650097 | — | iNaturalist 26872367 | USA | — | — |
| C. amethystina | C. zollingeri | OR824662 | — | iNaturalist 176453442 | USA | — | — |
| C. amethystina | C. zollingeri | OR800133 | — | iNaturalist 174766504 | USA | — | — |
| C. amethystina | C. zollingeri | ON059229 | — | iNaturalist 56904633 | USA | — | — |
| C. amethystina | C. zollingeri | MH016820 | — | FLAS-F-60642 | USA | — | — |
| C. amethystina | C. zollingeri | HQ021907 | — | 1Bart1426S | USA | — | — |
| C. amethystina | C. zollingeri | OR987432 | — | iNaturalist 176483913 | USA | — | — |
| C. amethystina | C. zollingeri | ON650096 | — | iNaturalist 26786656 | USA | — | — |
| C. amethystina | C. zollingeri | AY854071 | AY639882 | TENN 58652 | USA | — | — |
| C. amethystina | C. zollingeri | ON650095 | — | Mushroom Observer 209734 | USA | — | — |
| C. amethystina | C. zollingeri | MK575453 | — | SDR NAMA 2017-303 | USA | — | — |
| C. amethystina | C. zollingeri | OM473852 | — | iNaturalist 56881430 | USA | — | — |
| C. lilacina | C. zollingeri | MK427064 | — | ZP-1267 (MHHNU 7767) | China | — | — |
| C. lilacina | C. zollingeri | MW374244 | — | rxsbn-523 | China | — | — |
| C. lilacina | C. orientalis | PQ819512 | PQ814271 | MHHNU6801 | China | — | — |
| C. lilacina | C. orientalis | PQ819513 | PQ814272 | MHHNU7352 | China | — | — |
| C. lilacina | C. orientalis | PQ819514 | PQ814273 | MHHNU7586 | China | — | — |
| C. lilacina | C. orientalis | PQ819515 | PQ814274 | MHHNU7767 | China | — | — |
| C. lilacina | C. orientalis | PQ819516 | PQ814275 | MHHNU32116 | China | — | — |
| C. lilacina | C. zollingeri | OR567605 | — | JAC14886 (PDD 107102) | New Zealand | — | — |
| C. lilacina | C. zollingeri | OR567670 | — | JAC17787 (PDD 114897) | New Zealand | — | — |
| C. lilacina | C. zollingeri | OR567562 | OR567721 | JAC13837 (PDD 106042) | New Zealand | — | — |
| C. lilacina | C. zollingeri | PV399840 | PV399849 | PDD 81263 | New Zealand | 5.7 × 4.6 | 1.26 |
| C. lilacina | C. zollingeri | PV399841 | PV399850 | PDD 94782 | New Zealand | 5.8 × 4.8 | 1.22 |
| C. lilacina | C. aff. zollingeri | KP257143 | KP257212 | PBM3386 (TENN070808) | Australia | — | — |
| C. tongdaoensis | C. tongdaoensis | PQ819517 | PQ814276 | MHHNU11091 | China | — | — |
| C. tongdaoensis | C. tongdaoensis | PQ819518 | PQ814277 | MHHNU11093 | China | — | — |
| C. tongdaoensis | C. tongdaoensis | PQ819519 | PQ814278 | MHHNU11094 | China | — | — |
| C. violaceopulchra | C. zollingeri | PV603503 | — | BRACR29005 | Czech Republic | 5.6 × 4.9 | 1.13 |
| C. violaceopulchra | C. zollingeri | PV399842 | PV399850 | G00576171 | France | 6.2 × 5.2 | 1.18 |
| C. violaceopulchra | C. zollingeri | PV603504 | — | O-F-175336 | Norway | 6.1 × 5.2 | 1.17 |
| C. violaceopulchra | C. zollingeri | PV603511 | — | BRACR15130 | Slovakia | 6.1 × 4.8 | 1.26 |
| C. violaceopulchra | C. zollingeri | PV603505 | — | BRACR23959 | Slovakia | 6.1 × 4.8 | 1.28 |
| C. violaceopulchra | C. zollingeri | PV603506 | — | BRACR30724 | Slovakia | 5.9 × 4.6 | 1.28 |
| C. violaceopulchra | C. zollingeri | PV603507 | — | BRACR30732 | Slovakia | 6.1 × 4.8 | 1.27 |
| C. violaceopulchra | C. zollingeri | PV603510 | — | BRACR41672 | Slovakia | 5.9 × 5.2 | 1.14 |
| C. violaceopulchra | C. zollingeri | PV603508 | — | BRACR30735 | Slovakia | — | — |
| C. violaceopulchra | C. zollingeri | PV603509 | BRACR15909 | Slovakia | — | — | |
| C. violaceopulchra | C. zollingeri | PV399843 | PV399852 | ARAN-Fungi 22716 | Spain | 6 × 4.8 | 1.25 |
| C. violaceopulchra | C. zollingeri | PV399844 | PV399853 | BIO-Fungi 12557 | Spain | 5.7 × 4.8 | 1.18 |
| C. violaceopulchra | C. zollingeri | PV399845 | PV399854 | ARAN-Fungi A3078104 | Spain | 6 × 5.2 | 1.16 |
| C. violaceopulchra | C. zollingeri | OP538791 | — | 2.1.424 | Switzerland | — | — |
| C. violaceopulchra | C. zollingeri | OP538785 | — | 2.1.35 | Switzerland | — | — |
| C. violaceopulchra | C. zollingeri | OP538705 | — | 14_100a | Switzerland | — | — |
| C. violaceopulchra | C. sp. | OR858736 | — | iNaturalist 162964275 | USA | — | — |
| C. fumosa | KP257126 | KP257199 | TFB11839 (TENN060724) | Russia | — | — | |
| C. cf. fumosa | KP257127 | HQ877697 | TENN043695 | USA | — | — | |
| C. cf. rubicundula | HQ877696 | JN214482 | MR00170 (TENN064094) | USA | — | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olariaga, I.; Parra, L.A.; Læssøe, T.; Velasco, J.M.; Kautmanova, I.; Kruys, Å.; Salcedo, I. Disentangling a Complex of Violet, Endangered Species of Clavaria Subsumed Under the Misapplied Name Clavaria zollingeri Lév. (Clavariaceae, Fungi). J. Fungi 2025, 11, 482. https://doi.org/10.3390/jof11070482
Olariaga I, Parra LA, Læssøe T, Velasco JM, Kautmanova I, Kruys Å, Salcedo I. Disentangling a Complex of Violet, Endangered Species of Clavaria Subsumed Under the Misapplied Name Clavaria zollingeri Lév. (Clavariaceae, Fungi). Journal of Fungi. 2025; 11(7):482. https://doi.org/10.3390/jof11070482
Chicago/Turabian StyleOlariaga, Ibai, Luis A. Parra, Thomas Læssøe, Juan Manuel Velasco, Ivona Kautmanova, Åsa Kruys, and Isabel Salcedo. 2025. "Disentangling a Complex of Violet, Endangered Species of Clavaria Subsumed Under the Misapplied Name Clavaria zollingeri Lév. (Clavariaceae, Fungi)" Journal of Fungi 11, no. 7: 482. https://doi.org/10.3390/jof11070482
APA StyleOlariaga, I., Parra, L. A., Læssøe, T., Velasco, J. M., Kautmanova, I., Kruys, Å., & Salcedo, I. (2025). Disentangling a Complex of Violet, Endangered Species of Clavaria Subsumed Under the Misapplied Name Clavaria zollingeri Lév. (Clavariaceae, Fungi). Journal of Fungi, 11(7), 482. https://doi.org/10.3390/jof11070482