


Virulence Factors and Molecular Identification of Candida Species Causing Candidemia in Honduras


 , ,
, ,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Candida Isolation Procedures
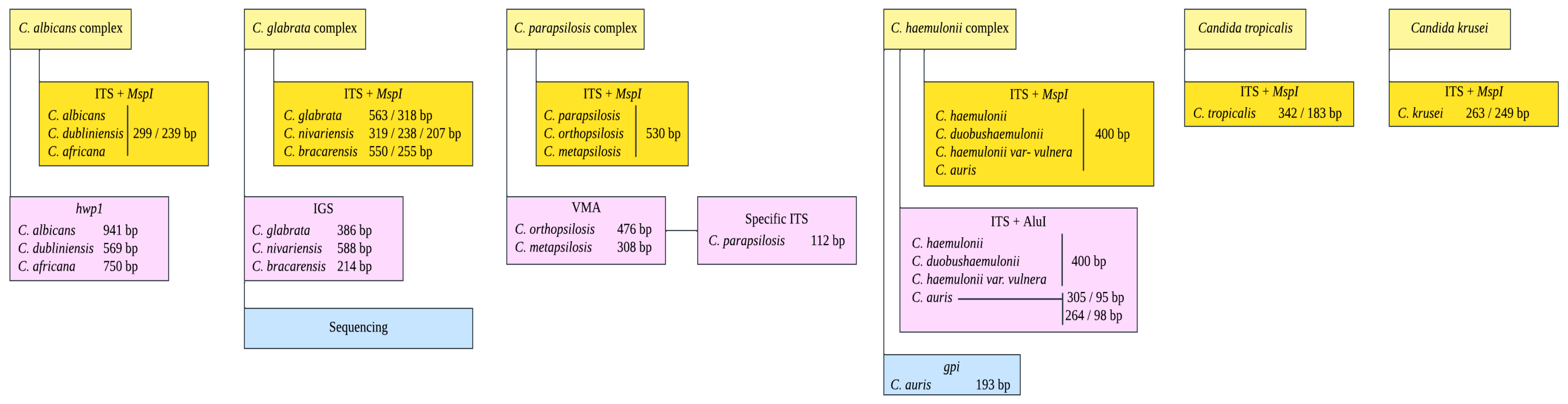
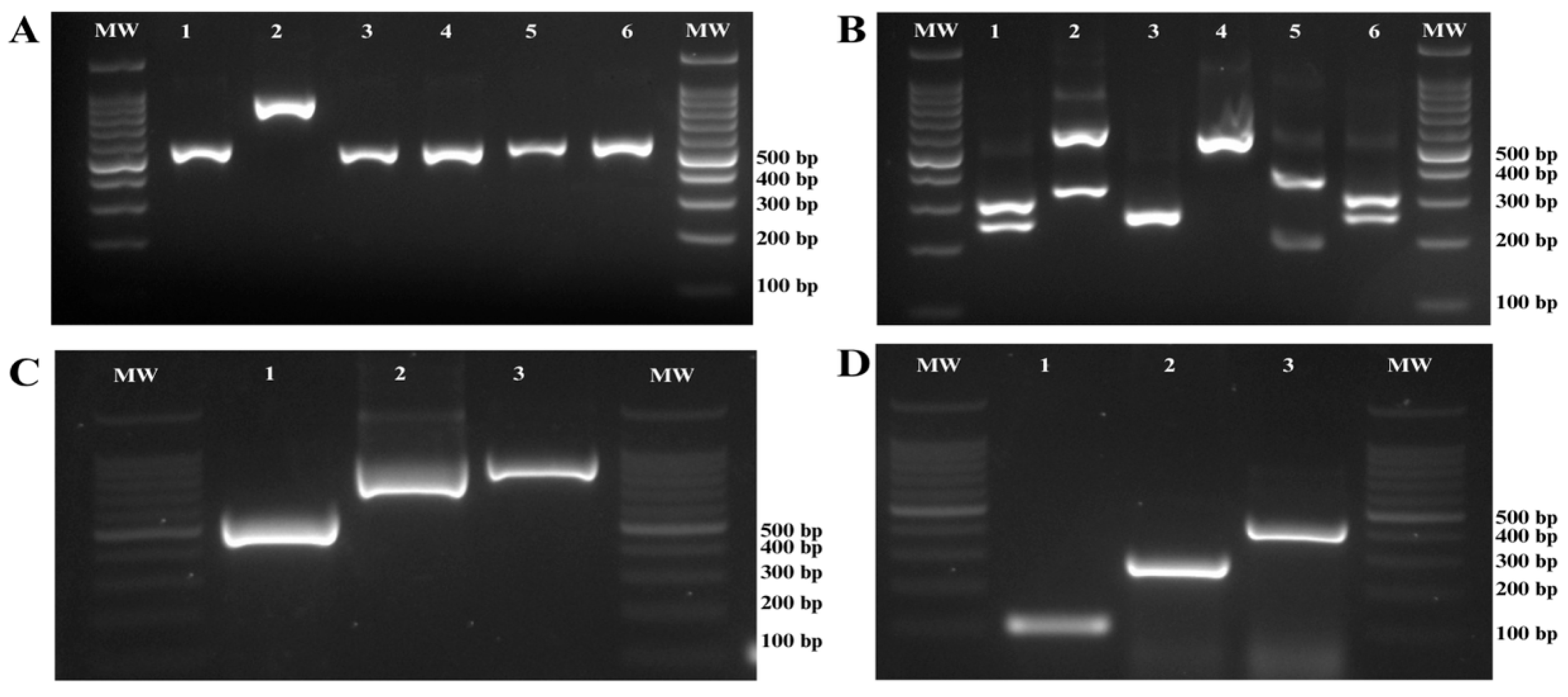
2.2. DNA Extraction and Molecular Identification of Candida Complexes
2.3. Hydrolytic Enzyme Activity Assays
2.4. Biofilm Formation
2.5. Statistical Analyses
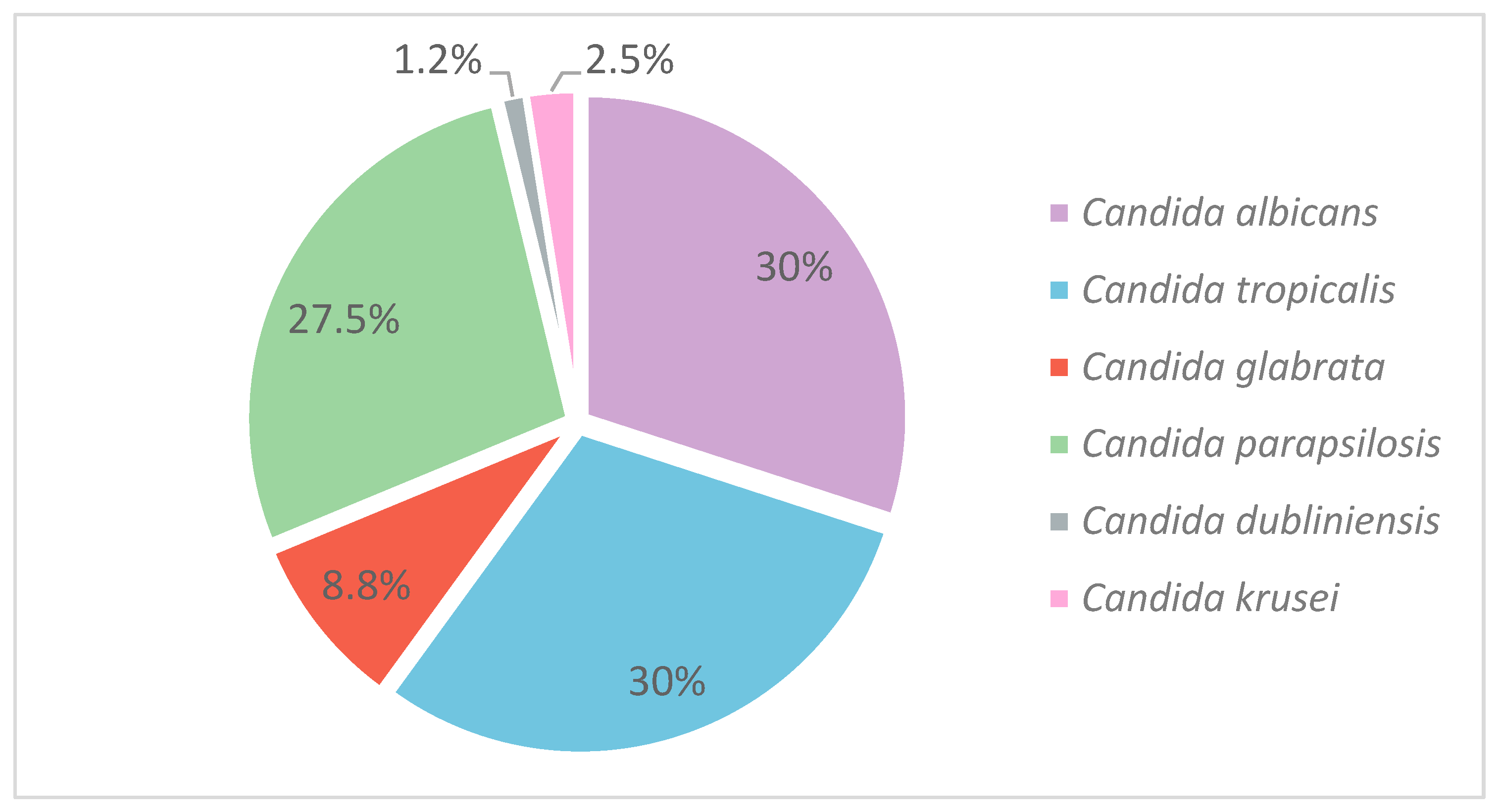
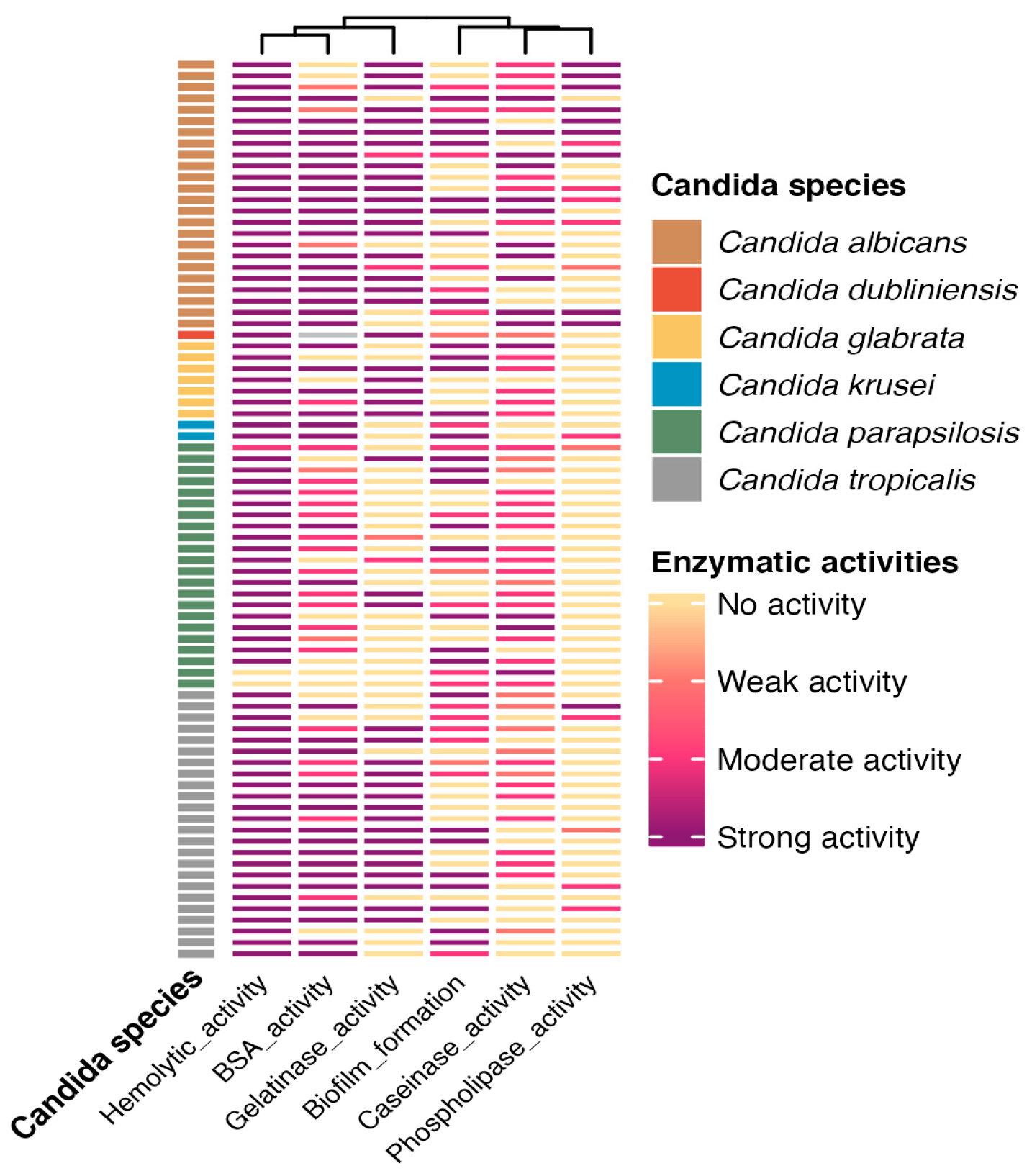
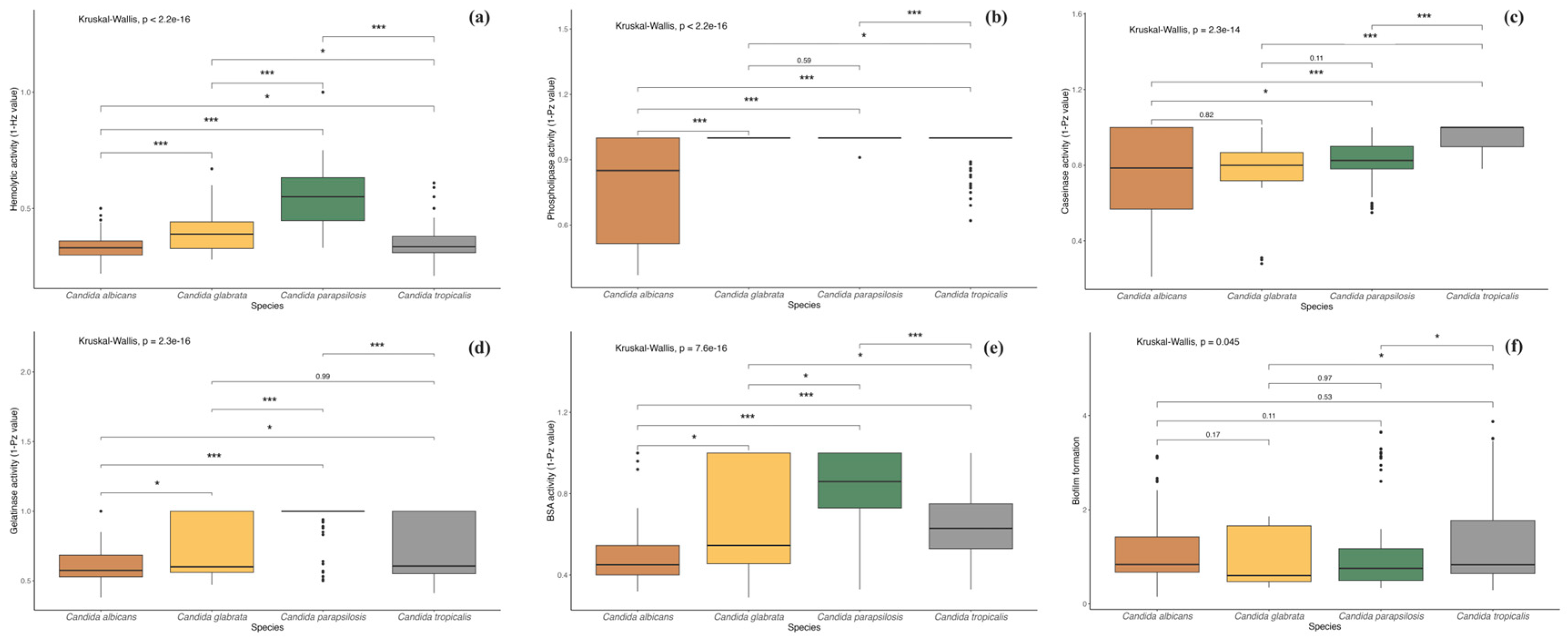
3. Results
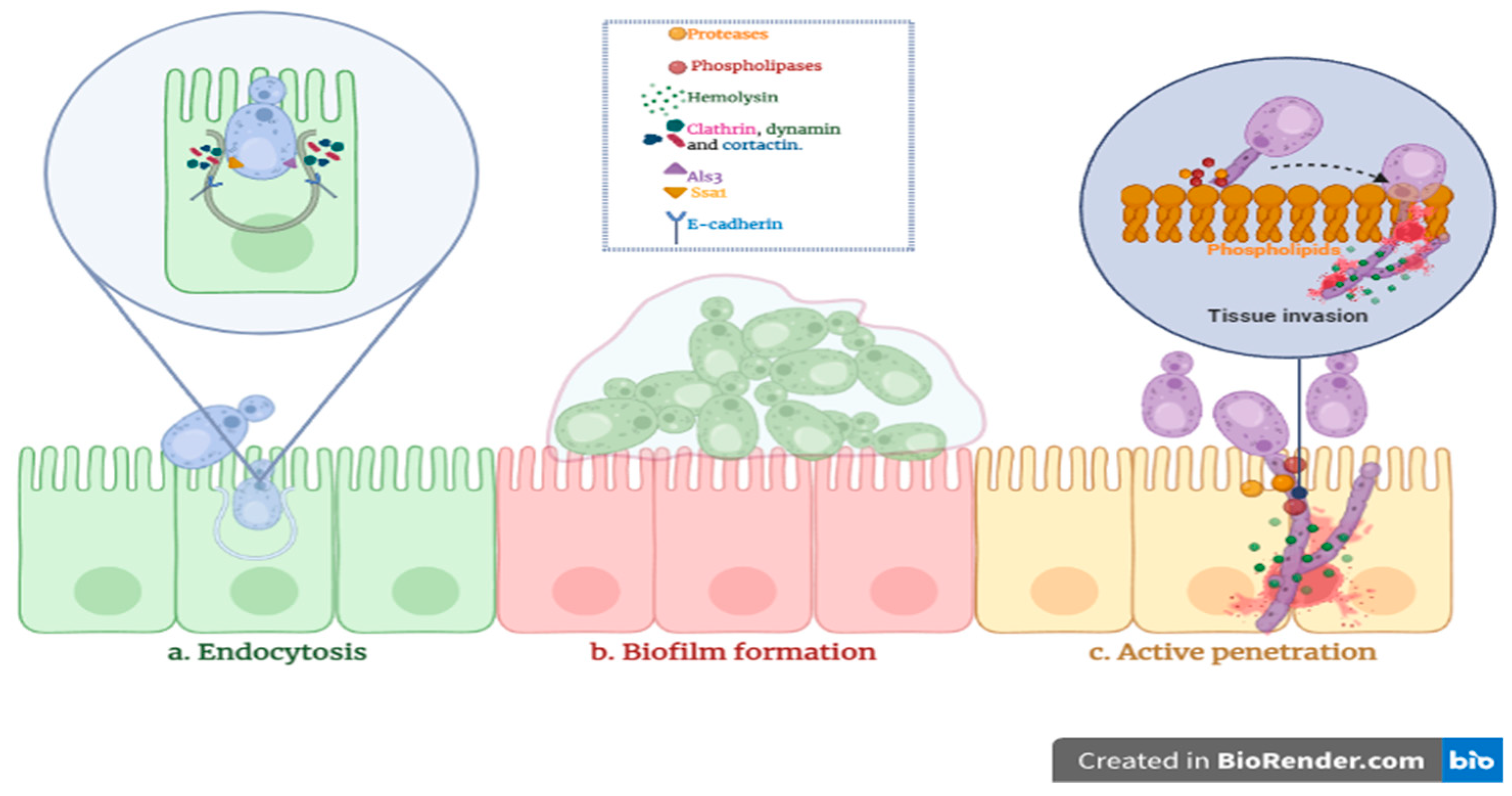
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denning, D.W. Global incidence and mortality of severe fungal disease. Lancet Infect. Dis. 2024, 24, e428–e438. [Google Scholar] [CrossRef] [PubMed]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases—Estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Hoenigl, M.; Strenger, V.; Buzina, W.; Valentin, T.; Koidl, C.; Wölfler, A.; Seeber, K.; Valentin, A.; Strohmeier, A.T.; Zollner-Schwetz, I. European Organization for the Research and Treatment of Cancer/Mycoses Study Group (EORTC/MSG) host factors and invasive fungal infections in patients with haematological malignancies. J. Antimicrob. Chemother. 2012, 67, 2029–2033. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, B.; Walsh, T.J.; Donnelly, J.P.; Stevens, D.A.; Edwards, J.E.; Calandra, T.; Pappas, P.G.; Maertens, J.; Lortholary, O.; Kauffman, C.A. Revised definitions of invasive fungal disease from the European organization for research and treatment of cancer/invasive fungal infections cooperative group and the national institute of allergy and infectious diseases mycoses study group (EORTC/MSG) consensus group. Clin. Infect. Dis. 2008, 46, 1813–1821. [Google Scholar]
- Wolfgruber, S.; Sedik, S.; Klingspor, L.; Tortorano, A.; Gow, N.A.; Lagrou, K.; Gangneux, J.-P.; Maertens, J.; Meis, J.F.; Lass-Flörl, C. Insights from three Pan-European multicentre studies on invasive Candida infections and outlook to ECMM Candida IV. Mycopathologia 2024, 189, 70. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty years of the SENTRY antifungal surveillance program: Results for Candida species from 1997–2016. Open Forum Infect. Dis. 2019, 6, S79–S94. [Google Scholar] [CrossRef]
- Guinea, J. Global trends in the distribution of C andida species causing candidemia. Clin. Microbiol. Infect. 2014, 20, 5–10. [Google Scholar] [CrossRef]
- Quindós, G.; Marcos-Arias, C.; San-Millán, R.; Mateo, E.; Eraso, E. The continuous changes in the aetiology and epidemiology of invasive candidiasis: From familiar Candida albicans to multiresistant Candida auris. Int. Microbiol. 2018, 21, 107–119. [Google Scholar] [CrossRef]
- Mulet Bayona, J.V.; Tormo Palop, N.; Salvador García, C.; Herrero Rodríguez, P.; Abril López de Medrano, V.; Ferrer Gómez, C.; Gimeno Cardona, C. Characteristics and management of candidaemia episodes in an established Candida auris outbreak. Antibiotics 2020, 9, 558. [Google Scholar] [CrossRef]
- Ngamchokwathana, C.; Chongtrakool, P.; Waesamaae, A.; Chayakulkeeree, M. Risk factors and outcomes of non-albicans Candida bloodstream infection in patients with Candidemia at Siriraj hospital—Thailand’s largest National Tertiary Referral Hospital. J. Fungi 2021, 7, 269. [Google Scholar] [CrossRef]
- Shigemura, K.; Osawa, K.; Jikimoto, T.; Yoshida, H.; Hayama, B.; Ohji, G.; Iwata, K.; Fujisawa, M.; Arakawa, S. Comparison of the clinical risk factors between Candida albicans and Candida non-albicans species for bloodstream infection. J. Antibiot. 2014, 67, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gaviria, M.; Ramírez-Sotelo, U.; Mora-Montes, H.M. Non-albicans Candida species: Immune response, evasion mechanisms, and new plant-derived alternative therapies. J. Fungi 2022, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-J.; Song, Y.-H.; Lv, G.; Liu, J.-Y.; Zhao, J.-T.; Wang, L.-L.; Xiang, M.-J. Emergence of invasive candidiasis with multiple Candida species exhibiting azole and echinocandin resistance. Front. Microbiol. 2025, 16, 1550894. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tian, S.; Han, X.; Chu, Y.; Wang, Q.; Zhou, B.; Shang, H. Is the superbug fungus really so scary? A systematic review and meta-analysis of global epidemiology and mortality of Candida auris. BMC Infect. Dis. 2020, 20, 827. [Google Scholar] [CrossRef]
- Salmanton-García, J.; Cornely, O.A.; Stemler, J.; Barać, A.; Steinmann, J.; Siváková, A.; Akalin, E.H.; Arikan-Akdagli, S.; Loughlin, L.; Toscano, C. Attributable mortality of candidemia–results from the ECMM Candida III multinational European observational cohort study. J. Infect. 2024, 89, 106229. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Mulet Bayona, J.V.; Tormo Palop, N.; Salvador García, C.; Fuster Escrivá, B.; Chanzá Aviñó, M.; Ortega García, P.; Gimeno Cardona, C. Impact of the SARS-CoV-2 pandemic in candidaemia, invasive aspergillosis and antifungal consumption in a tertiary hospital. J. Fungi 2021, 7, 440. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef]
- Mba, I.E.; Nweze, E.I. Mechanism of Candida pathogenesis: Revisiting the vital drivers. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1797–1819. [Google Scholar] [CrossRef]
- Saiprom, N.; Wongsuk, T.; Oonanant, W.; Sukphopetch, P.; Chantratita, N.; Boonsilp, S. Characterization of Virulence Factors in Candida Species Causing Candidemia in a Tertiary Care Hospital in Bangkok, Thailand. J. Fungi 2023, 9, 353. [Google Scholar] [CrossRef]
- Makled, A.F.; Ali, S.A.; Labeeb, A.Z.; Salman, S.S.; Shebl, D.Z.; Hegazy, S.G.; Sabal, M.S. Characterization of Candida species isolated from clinical specimens: Insights into virulence traits, antifungal resistance and molecular profiles. BMC Microbiol. 2024, 24, 388. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, B.; Pérez-Alemán, E.; Galo, C.; Fontecha, G. Molecular identification of Candida species from urinary infections in Honduras. Rev. Iberoam. Micol. 2018, 35, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Montes, K.; Ortiz, B.; Galindo, C.; Figueroa, I.; Braham, S.; Fontecha, G. Identification of Candida species from clinical samples in a Honduran tertiary hospital. Pathogens 2019, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- Romeo, O.; Criseo, G. First molecular method for discriminating between Candida africana, Candida albicans, and Candida dubliniensis by using hwp1 gene. Diagn. Microbiol. Infect. Dis. 2008, 62, 230–233. [Google Scholar] [CrossRef]
- Arastehfar, A.; Fang, W.; Pan, W.; Liao, W.; Yan, L.; Boekhout, T. Identification of nine cryptic species of Candida albicans, C. glabrata, and C. parapsilosis complexes using one-step multiplex PCR. BMC Infect. Dis. 2018, 18, 480. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Ruiz-Gaitán, A.C.; Fernández-Pereira, J.; Valentin, E.; Tormo-Mas, M.A.; Eraso, E.; Pemán, J.; de Groot, P.W. Molecular identification of Candida auris by PCR amplification of species-specific GPI protein-encoding genes. Int. J. Med. Microbiol. 2018, 308, 812–818. [Google Scholar] [CrossRef]
- Neji, S.; Hadrich, I.; Trabelsi, H.; Abbes, S.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Ayadi, A. Virulence factors, antifungal susceptibility and molecular mechanisms of azole resistance among Candida parapsilosis complex isolates recovered from clinical specimens. J. Biomed. Sci. 2017, 24, 67. [Google Scholar] [CrossRef]
- Price, M.F.; Wilkinson, I.D.; Gentry, L.O. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia J. Med. Vet. Mycol. 1982, 20, 7–14. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.3.1 V; R Foundation for Statistical Computing: Vienna, Austria, 2023. Available online: https://www.R-project.org/ (accessed on 11 March 2025).
- Hadley, W. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Agudelo Higuita, N.I.; Varela Bustillo, D.; Denning, D.W. Burden of serious fungal infections in Honduras. Mycoses 2022, 65, 429–439. [Google Scholar] [CrossRef]
- Global Action Fund for Fungal Infections. The Burden of Serious Fungal Infections in Latin America. 2019. Available online: https://life-slides-and-videos.s3.eu-west-2.amazonaws.com/LIFE+articles/Burden+of+serious+fungal+infections+in+Latin+America.pdf (accessed on 12 March 2025).
- Ortiz, B.; Varela, D.; Fontecha, G.; Torres, K.; Cornely, O.A.; Salmanton-García, J. Strengthening Fungal Infection Diagnosis and Treatment: An In-depth Analysis of Capabilities in Honduras. Open Forum Infect. Dis. 2024, 11, ofae578. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. Respuesta Mundial al VIH. Available online: https://www.unaids.org/es (accessed on 9 March 2025).
- Secretaría de Salud. Informe Estadistico de la Epidemia de VIH en Honduras período 1985—Marzo 2024. 2024. Available online: https://salud.gob.hn/sshome/index.php/component/jdownloads/?task=download.send&id=1522 (accessed on 31 January 2025).
- Sistema Estadístico Nacional, I. Honduras. Cifras de País. Available online: https://sen.ine.gob.hn/Home/Index?ReturnUrl=%2F (accessed on 16 February 2025).
- Silva, D.L.; Peres, N.T.; Santos, D.A. Key fungal coinfections: Epidemiology, mechanisms of pathogenesis, and beyond. mBio 2025, 16, e0056225. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Rodrigues, M.E.; Henriques, M. Candida sp. infections in patients with diabetes mellitus. J. Clin. Med. 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- OPS Honduras. IV Congreso Nacional de Diabetes. Available online: https://www.paho.org/es/historias/atencion-pacientes-diabeticos-personas-con-problemas-salud-mental-tiempos-covid-19 (accessed on 13 January 2025).
- Instituto Nacional del Diabético. ¿Sabías Que en Honduras Hay 1 Millón de Diabéticos? Available online: https://inadi.gob.hn/sitio/sabias-que-en-honduras-hay-1-millon-de-diabeticos/ (accessed on 13 January 2025).
- Rodríguez, L.U.P.; Arévalo, Á.R.E. Caracterización del sistema de salud de Honduras. Rev. Méd. Hondureña 2018, 86, 22–27. [Google Scholar]
- González, K.J.S. Deficiencias en el sistema de salud pública y su impacto en la pandemia del COVID-19. Rev. Méd. Hondureña 2021, 89, 148–150. [Google Scholar] [CrossRef]
- Fajardo Leitzelar, F.; Ramírez-Osorto, L.J.; Pérez Arias, C.M.; Benítez López, A.A.; Ordóñez Banegas, G.M.; Gómez-Flores, E.O.; Merino, A. Prevalencia y caracterización de nefropatía de etiología no determinada en hospital de segundo nivel, Honduras. Rev. Colomb. Nefrol. 2024, 11. [Google Scholar] [CrossRef]
- Riera, F.O.; Caeiro, J.P.; Angiolini, S.C.; Vigezzi, C.; Rodriguez, E.; Icely, P.A.; Sotomayor, C.E. Invasive candidiasis: Update and current challenges in the management of this mycosis in South America. Antibiotics 2022, 11, 877. [Google Scholar] [CrossRef]
- Nucci, M.; Queiroz-Telles, F.; Alvarado-Matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J. Epidemiology of candidemia in Latin America: A laboratory-based survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef]
- Ortiz, B.; Aguilar, K.; Galindo, C.; Molina, L.; Fontecha, G. Candida species isolated from clinical samples in a tertiary hospital in Honduras: Where is Candida auris? Curr. Med. Mycol. 2022, 8, 1. [Google Scholar] [CrossRef]
- Falci, D.R.; Pasqualotto, A.C. Clinical mycology in Latin America and the Caribbean: A snapshot of diagnostic and therapeutic capabilities. Mycoses 2019, 62, 368–373. [Google Scholar] [CrossRef]
- Salmanton-García, J.; Au, W.-Y.; Hoenigl, M.; Chai, L.Y.A.; Badali, H.; Basher, A.; Brockhoff, R.A.; Chen, S.C.-A.; Chindamporn, A.; Chowdhary, A. The current state of laboratory mycology in Asia/Pacific: A survey from the European Confederation of Medical Mycology (ECMM) and International Society for Human and Animal Mycology (ISHAM). Int. J. Antimicrob. Agents 2023, 61, 106718. [Google Scholar] [CrossRef] [PubMed]
- Sifuentes-Osornio, J.; Corzo-León, D.E.; Ponce-de-León, L.A. Epidemiology of invasive fungal infections in Latin America. Curr. Fungal Infect. Rep. 2012, 6, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Dhakad, M.S.; Goyal, R.; Haque, A.; Mukhopadhyay, G. Identification and antifungal susceptibility testing of Candida species: A comparison of Vitek-2 system with conventional and molecular methods. J. Glob. Infect. Dis. 2016, 8, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Won, E.J.; Shin, J.H.; Kim, M.-N.; Choi, M.J.; Joo, M.Y.; Kee, S.J.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Evaluation of the BD Phoenix system for identification of a wide spectrum of clinically important yeast species: A comparison with Vitek 2-YST. Diagn. Microbiol. Infect. Dis. 2014, 79, 477–480. [Google Scholar] [CrossRef]
- Morovati, H.; Kord, M.; Ahmadikia, K.; Eslami, S.; Hemmatzadeh, M.; Kurdestani, K.M.; Khademi, M.; Darabian, S. A comprehensive review of identification methods for pathogenic yeasts: Challenges and approaches. Adv. Biomed. Res. 2023, 12, 187. [Google Scholar] [CrossRef]
- Calderón-Hernández, A.; Castro-Bonilla, N.; Cob-Delgado, M. Chromogenic, biochemical and proteomic identification of yeast and yeast-like microorganisms isolated from clinical samples from animals of costa rica. J. Fungi 2024, 10, 218. [Google Scholar] [CrossRef]
- Ambaraghassi, G.; Dufresne, P.J.; Dufresne, S.F.; Vallières, É.; Muñoz, J.F.; Cuomo, C.A.; Berkow, E.L.; Lockhart, S.R.; Luong, M.-L. Identification of Candida auris by use of the updated Vitek 2 yeast identification system, version 8.01: A multilaboratory evaluation study. J. Clin. Microbiol. 2019, 57, e00884-19. [Google Scholar] [CrossRef]
- Zurita, J.; y Miño, A.P.; Solís, M.; Sevillano, G. Failed identification of Candida vulturna using the updated Vitek 2 yeast identification system, version 9.02 and CHROMagar Candida Plus. New Microbes New Infect. 2022, 48, 101012. [Google Scholar] [CrossRef]
- Setoguchi, D.; Iwanaga, N.; Ito, Y.; Hirayama, T.; Yoshida, M.; Takeda, K.; Ide, S.; Nagayoshi, Y.; Kondo, A.; Tashiro, M. Case report and literature review of refractory fungemia caused by Candida vulturna. Heliyon 2024, 10, e31464. [Google Scholar] [CrossRef]
- Benkova, M.; Soukup, O.; Marek, J. Antimicrobial susceptibility testing: Currently used methods and devices and the near future in clinical practice. J. Appl. Microbiol. 2020, 129, 806–822. [Google Scholar] [CrossRef]
- Fontecha, G.; Montes, K.; Ortiz, B.; Galindo, C.; Braham, S. Identification of cryptic species of four Candida complexes in a culture collection. J. Fungi 2019, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Wächtler, B.; Citiulo, F.; Jablonowski, N.; Förster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-epithelial interactions: Dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE 2012, 7, e36952. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A. Potential role of phospholipases in virulence and fungal pathogenesis. Clin. Microbiol. Rev. 2000, 13, 122–143. [Google Scholar] [CrossRef] [PubMed]
- Hube, B.; Rüchel, R.; Monod, M.; Sanglard, D.; Odds, F.C. Functional aspects of secreted Candida proteinases. In Aspartic Proteinases: Retroviral and Cellular Enzymes; Springer: Boston, MA, USA, 1998; pp. 339–344. [Google Scholar]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef]
- Silva, S.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida species biofilms’ antifungal resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef]
- El-Kholy, M.A.; Helaly, G.F.; El Ghazzawi, E.F.; El-Sawaf, G.; Shawky, S.M. Virulence factors and antifungal susceptibility profile of C. tropicalis isolated from various clinical specimens in Alexandria, Egypt. J. Fungi 2021, 7, 351. [Google Scholar] [CrossRef]
- de Melo, A.V.; Zuza-Alves, D.; da Silva-Rocha, W.; de Souza, L.F.C.; Francisco, E.; de Azevedo Melo, A.S.; Chaves, G.M. Virulence factors of Candida spp. obtained from blood cultures of patients with candidemia attended at tertiary hospitals in Northeast Brazil. J. Mycol. Medicale 2019, 29, 132–139. [Google Scholar] [CrossRef]
- Tiwari, P.; Nikhil, A.; Tiwari, K.; Paonikar, T.; Gupta, M.K.; Tilak, R. In vitro determination of antifungal susceptibility and virulence factors in Candida species causing candidemia in North India Region. Discov. Public Health 2024, 21, 50. [Google Scholar] [CrossRef]
- Pandey, N.; Gupta, M.K.; Tilak, R. Extracellular hydrolytic enzyme activities of the different Candida spp. isolated from the blood of the Intensive Care Unit-admitted patients. J. Lab. Physicians 2018, 10, 392–396. [Google Scholar] [CrossRef]
- Nouraei, H.; Pakshir, K.; ZareShahrabadi, Z.; Zomorodian, K. High detection of virulence factors by Candida species isolated from bloodstream of patients with candidemia. Microb. Pathog. 2020, 149, 104574. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.S.; Mirbod, F.; Filler, S.G.; Banno, Y.; Cole, G.T.; Kitajima, Y.; Edwards, J.E., Jr.; Nozawa, Y.; Ghannoum, M.A. Evidence implicating phospholipase as a virulence factor of Candida albicans. Infect. Immun. 1995, 63, 1993–1998. [Google Scholar] [CrossRef] [PubMed]
- Gaziano, R.; Sabbatini, S.; Monari, C. The interplay between Candida albicans, vaginal mucosa, host immunity and resident microbiota in health and disease: An overview and future perspectives. Microorganisms 2023, 11, 1211. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, A. Molecular determinants involved in Candida albicans biofilm formation and regulation. Mol. Biotechnol. 2024, 66, 1640–1659. [Google Scholar] [CrossRef]
- Sathi, F.A.; Aung, M.S.; Paul, S.K.; Nasreen, S.A.; Haque, N.; Roy, S.; Ahmed, S.; Alam, M.M.; Khan, S.; Rabbany, M.A. Clonal Diversity of Candida auris, Candida blankii, and Kodamaea ohmeri Isolated from Septicemia and Otomycosis in Bangladesh as Determined by Multilocus Sequence Typing. J. Fungi 2023, 9, 658. [Google Scholar] [CrossRef]
- Mantecón-Vallejo, M.d.l.Á.; Mesquida, A.; Ortiz, M.d.V.; Buzón-Martín, L.; Ossa-Echeverri, S.; Fisac-Cuadrado, L.; Megías-Lobón, G.; Ortega-Lafont, M.P.; Muñoz, P.; Escribano, P. Clonal spread of fluconazole-resistant C. parapsilosis in patients admitted to a referral hospital located in Burgos, Spain, during the COVID-19 pandemic. Mycoses 2024, 67, e13685. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Hilmioğlu-Polat, S.; Fang, W.; Yaşar, M.; Polat, F.; Metin, D.Y.; Rigole, P.; Coenye, T.; Ilkit, M. First report of candidemia clonal outbreak caused by emerging fluconazole-resistant Candida parapsilosis isolates harboring Y132F and/or Y132F+ K143R in Turkey. Antimicrob. Agents Chemother. 2020, 64, e01001-20. [Google Scholar] [CrossRef]
- Boonsilp, S.; Homkaew, A.; Phumisantiphong, U.; Nutalai, D.; Wongsuk, T. Species distribution, antifungal susceptibility, and molecular epidemiology of Candida species causing candidemia in a tertiary care hospital in Bangkok, Thailand. J. Fungi 2021, 7, 577. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Locus | Sequence 5′ to 3′ | Annealing Temperature °C | Reference |
|---|---|---|---|---|
| ITS1 | ITS-ADNr | TCCGTAGGTGAACCTGCGG | 56 | [26] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| CR-f | hpw1 | GCTACCACTTCAGAATCATCATC | 58 | [24] |
| CR-r | GCACCTTCAGTCGTAGAGACG | |||
| OM-f | VMA | GAGAAAGCACGCCTCTTTGC | 60 | [25] |
| OM-r | TCAGCATTTTGGGCTCTTGC | |||
| P-f | ITS-ADNr | GCGGAAGGATCATTACAGAATG | 60 | |
| P-r | CTGGCAGGCCCCATATAG | |||
| PCG-f | IGS | TCACTTTCAACTGCTTTCGC | 60 | |
| G-r | TGCGAGTCATGGGCGGAA | |||
| N-r | ACCCCAGAGGCATAAATAGC | 60 | ||
| B-r | GCAACTGGACGAAAGTGC | |||
| 03410_F | GPI | GCCGCTAGATTGATCACCGT | 60 | [27] |
| 03410_R | TAGGTGTGGGTACCCTTGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chávez, J.F.; Ortiz, B.; López, R.; Muñoz, C.; Aguilar, K.; Laínez-Arteaga, I.; Galindo, C.; Rivera, L.; Ballesteros-Monrreal, M.G.; Montes, K.; et al. Virulence Factors and Molecular Identification of Candida Species Causing Candidemia in Honduras. J. Fungi 2025, 11, 470. https://doi.org/10.3390/jof11070470
Chávez JF, Ortiz B, López R, Muñoz C, Aguilar K, Laínez-Arteaga I, Galindo C, Rivera L, Ballesteros-Monrreal MG, Montes K, et al. Virulence Factors and Molecular Identification of Candida Species Causing Candidemia in Honduras. Journal of Fungi. 2025; 11(7):470. https://doi.org/10.3390/jof11070470
Chicago/Turabian StyleChávez, José Fernando, Bryan Ortiz, Roque López, Carlos Muñoz, Kateryn Aguilar, Isis Laínez-Arteaga, Celeste Galindo, Luis Rivera, Manuel G. Ballesteros-Monrreal, Kathy Montes, and et al. 2025. "Virulence Factors and Molecular Identification of Candida Species Causing Candidemia in Honduras" Journal of Fungi 11, no. 7: 470. https://doi.org/10.3390/jof11070470
APA StyleChávez, J. F., Ortiz, B., López, R., Muñoz, C., Aguilar, K., Laínez-Arteaga, I., Galindo, C., Rivera, L., Ballesteros-Monrreal, M. G., Montes, K., Hernández, M., Barahona, A. V., Hereira-Pacheco, S., & Fontecha, G. (2025). Virulence Factors and Molecular Identification of Candida Species Causing Candidemia in Honduras. Journal of Fungi, 11(7), 470. https://doi.org/10.3390/jof11070470